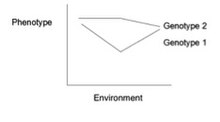
Response to the same environmental variation differently by different genotypes
«GXE» перенаправляется сюда. Чтобы узнать о самолете, см. Waco GXE .
Взаимодействие гена и окружающей среды (или взаимодействие генотипа и окружающей среды , или G×E ) — это когда два разных генотипа по-разному реагируют на изменения окружающей среды. Норма реакции — это график, показывающий взаимосвязь между генами и факторами окружающей среды , когда фенотипические различия непрерывны. [1] Они могут помочь проиллюстрировать взаимодействие GxE. Когда норма реакции не параллельна, как показано на рисунке ниже, происходит взаимодействие гена с окружающей средой. Это указывает на то, что каждый генотип по-разному реагирует на изменения окружающей среды. Изменения окружающей среды могут быть физическими, химическими, биологическими, моделями поведения или жизненными событиями. [2]
Эта норма реакции показывает непараллельные линии, указывающие на взаимодействие гена с окружающей средой. Каждый генотип по-разному реагирует на изменения окружающей среды.
Взаимодействия гена и окружающей среды изучаются, чтобы лучше понять различные явления. В генетической эпидемиологии взаимодействие генов и окружающей среды полезно для понимания некоторых заболеваний . Иногда чувствительность к факторам риска заболевания окружающей среды передается по наследству, а не само заболевание. На людей с разными генотипами воздействие одних и тех же факторов окружающей среды по-разному влияет, и, таким образом, взаимодействие генов и окружающей среды может приводить к различным фенотипам заболеваний. Например, воздействие солнечного света оказывает более сильное влияние на рака кожи риск у людей со светлой кожей, чем у людей с более темной кожей . [3]
Эти взаимодействия представляют особый интерес для генетических эпидемиологов для прогнозирования уровня заболеваемости и методов профилактики с точки зрения общественного здравоохранения. [2] Этот термин также используется психологами развития , чтобы лучше понять индивидуальное и эволюционное развитие. [4]
Споры о природе и воспитании предполагают, что вариации признака в первую очередь обусловлены либо генетическими различиями, либо различиями в окружающей среде. Однако нынешнее научное мнение гласит, что ни генетические различия, ни различия окружающей среды не несут исключительной ответственности за возникновение фенотипических вариаций и что практически все признаки находятся под влиянием как генетических, так и экологических различий. [5] [6] [7]
Статистический анализ генетических и экологических различий, способствующих фенотипу, должен быть использован для подтверждения того, что это взаимодействие гена и окружающей среды. В генетике развития причинного взаимодействия достаточно для подтверждения взаимодействия гена и окружающей среды. [8]
История определения взаимодействия генов и окружающей среды восходит к 1930-м годам и сегодня остается темой дискуссий. Первый случай дебатов произошел между Рональдом Фишером и Ланселотом Хогбеном .Фишер стремился исключить взаимодействие из статистических исследований, поскольку это явление можно было устранить, изменив масштаб. Хогбен считал, что это взаимодействие следует исследовать, а не устранять, поскольку оно дает информацию о причинно-следственной связи определенных элементов развития.
С аналогичным аргументом столкнулись многие учёные в 1970-х годах. Артур Дженсен опубликовал исследование « Насколько мы можем повысить IQ и успеваемость?» », что, помимо большой критики, также вызвало споры со стороны ученых Ричарда Левонтина и Дэвида Лайзера . Левонтин и Лайзер утверждали, что для того, чтобы сделать вывод о причинно-следственных механизмах, нельзя игнорировать взаимодействие гена и окружающей среды в контексте исследования, в то время как Дженсен утверждал, что взаимодействие является чисто статистическим явлением и не связано с развитием. [9]
Примерно в то же время Кеннет Дж. Ротман поддержал использование статистического определения взаимодействия, в то время как исследователи Куппер и Хоган считали, что определение и существование взаимодействия зависят от используемой модели. [10]
Самая последняя критика была вызвана исследованиями Моффита и Каспи о 5-HTTLPR , стрессе и его влиянии на депрессию. В отличие от предыдущих дебатов, Моффитт и Каспи теперь использовали статистический анализ, чтобы доказать, что взаимодействие существует и может быть использовано для раскрытия механизмов уязвимости. Утверждение исходило от Заммита, Оуэна и Льюиса, которые подтвердили обеспокоенность Фишера тем, что статистический эффект не был связан с процессом развития и не может быть воспроизведен с разницей в масштабе. [9]
Сегодня существуют две разные концепции взаимодействия генов и окружающей среды. Табери [11] назвал их биометрическим взаимодействием и взаимодействием развития , в то время как Сесардический [12] использует термины «статистическое взаимодействие» и «здравый смысл» .
Биометрическая (или статистическая) концепция берет свое начало в исследовательских программах, направленных на измерение относительных пропорций генетического и экологического вклада в фенотипические вариации внутри популяций. Биометрическое взаимодействие гена и окружающей среды имеет особое значение в популяционной генетике и поведенческой генетике . [11] Любое взаимодействие приводит к нарушению аддитивности основных эффектов наследственности и окружающей среды, но присутствует ли такое взаимодействие в определенных условиях — эмпирический вопрос. Биометрическое взаимодействие актуально в контексте исследования индивидуальных различий, а не в контексте развития конкретного организма. [4]
Взаимодействие генов и окружающей среды в процессе развития — это концепция, чаще используемая генетиками развития и психобиологами развития . Взаимодействие в процессе развития не рассматривается просто как статистическое явление. Независимо от того, присутствует статистическое взаимодействие или нет, онтогенетическое взаимодействие в любом случае проявляется в причинном взаимодействии генов и окружающей среды при создании фенотипа индивидуума. [4]
В эпидемиологии для группировки различных взаимодействий между геном и окружающей средой можно использовать следующие модели.
Модель А описывает генотип, который повышает уровень экспрессии фактора риска, но не вызывает самого заболевания. Например, ген ФКУ приводит к более высокому уровню фенилаланина, чем обычно, что, в свою очередь, вызывает умственную отсталость.
Фактор риска в модели B, напротив, оказывает прямое влияние на восприимчивость к заболеваниям, которая усиливается генетической предрасположенностью. Модель C демонстрирует обратную картину: генетическая предрасположенность напрямую влияет на заболевание, в то время как фактор риска усиливает этот эффект. В каждой независимой ситуации фактор, непосредственно воздействующий на заболевание, сам по себе может вызвать заболевание.
Модель D отличается тем, что ни один фактор в этой ситуации не может повлиять на риск заболевания, однако, когда присутствуют и генетическая предрасположенность, и фактор риска, риск увеличивается. Например, ген дефицита G6PD в сочетании с потреблением фасоли приводит к гемолитической анемии. Это заболевание не возникает ни у людей, которые едят конские бобы и не имеют дефицита Г6ФД, ни у людей с дефицитом Г6ФД, которые не едят конские бобы.
Наконец, Модель E описывает сценарий, в котором фактор риска окружающей среды и генетическая восприимчивость могут индивидуально влиять на риск заболевания. Однако в сочетании влияние на риск заболевания различно.
Модели ограничены тем фактом, что переменные являются двоичными и поэтому не учитывают сценарии переменных полигенного или непрерывного масштаба. [2]
Исследования усыновления использовались для того, чтобы выяснить, насколько усыновленные люди похожи на своих биологических родителей, с которыми они не жили в одной среде. Кроме того, усыновленных сравнивают с приемной семьей из-за различий в генах, но в общей среде обитания. Например, исследование по усыновлению показало, что шведские мужчины с неблагоприятной средой усыновления и генетической предрасположенностью чаще злоупотребляют алкоголем. [13]
Используя монозиготных близнецов , можно было наблюдать влияние различных сред на идентичные генотипы. Более поздние исследования используют методы биометрического моделирования для сравнения дизиготных близнецов, чтобы в конечном итоге определить различные уровни экспрессии генов в разных средах. [13]
Семейные исследования направлены на сравнение контрольной группы с низким уровнем риска и детей с высоким риском, чтобы определить влияние окружающей среды на субъектов с различными уровнями генетического риска. Например, датское исследование детей из группы высокого риска, чьи матери страдали шизофренией, показало, что дети без постоянного опекуна были связаны с повышенным риском развития шизофрении. [13]
Часто используемый метод обнаружения взаимодействий гена и окружающей среды заключается в изучении влияния одной вариации гена ( гена-кандидата ) на конкретную среду. Однонуклеотидные полиморфизмы (SNP) сравниваются с одиночными бинарными факторами воздействия для определения каких-либо эффектов.
Кандидатские исследования, подобные этим, требуют убедительных биологических гипотез, которые в настоящее время трудно выбрать, учитывая недостаточное понимание биологических механизмов, которые приводят к более высокому риску.
Эти исследования также часто трудно воспроизвести из-за небольших размеров выборки, что обычно приводит к спорным результатам.
Полигенная природа сложных фенотипов предполагает , что исследования отдельных кандидатов могут быть неэффективными при определении различных эффектов меньшего масштаба от большого количества влияющих вариантов генов. [14]
Поскольку один и тот же фактор окружающей среды может взаимодействовать с несколькими генами, для анализа взаимодействий GxE можно использовать полигенный подход. Полигенный показатель генерируется с использованием аллелей, связанных с признаком, и их соответствующих весов на основе эффекта и исследуется в сочетании с воздействием окружающей среды. Хотя этот метод исследования еще является ранним, он соответствует психическим расстройствам. В результате совпадения эндофенотипов среди расстройств это позволяет предположить, что результаты взаимодействия гена и окружающей среды применимы к различным диагнозам. [14]
Полногеномные исследования ассоциаций и полногеномные исследования взаимодействия
Подход полногеномного сканирования взаимодействия (GEWIS) исследует взаимодействие между окружающей средой и большим количеством независимых SNP. Эффективный подход к этому всеобъемлющему исследованию состоит из двух этапов: сначала геном фильтруется с использованием тестов на уровне генов и анализа набора генов на основе путей. На втором этапе используются ассоциации SNP с GE и тесты на взаимодействие. [15]
Гипотеза дифференциальной восприимчивости была подтверждена с помощью полногеномных подходов. [16]
Особую озабоченность в исследованиях взаимодействия генов и окружающей среды вызывает отсутствие воспроизводимости. Исследования особо сложных черт оказались под пристальным вниманием из-за результатов, которые невозможно воспроизвести. Например, исследования гена 5-HTTLPR и стресса, приводящего к изменению риска депрессии, дали противоречивые результаты. [17] [15]
Возможным объяснением противоречивых результатов является интенсивное использование множественных тестов. Предполагается, что исследования дают неточные результаты из-за изучения множества фенотипов и факторов окружающей среды в отдельных экспериментах. [15]
Существуют две разные модели шкалы измерения, которые помогают определить, существует ли взаимодействие гена и окружающей среды в статистическом контексте. Существуют разногласия по поводу того, какую шкалу следует использовать. Согласно этому анализу, если объединенные переменные соответствуют любой модели, взаимодействия нет. Комбинированные эффекты должны быть либо выше для синергического результата, либо меньше, чем для антагонистического результата. Аддитивная модель измеряет различия в рисках, а мультипликативная модель использует коэффициенты для измерения эффектов. Было высказано предположение, что аддитивная модель лучше подходит для прогнозирования риска заболеваний в популяции, тогда как мультипликативная модель больше подходит для оценки этиологии заболевания. [2]
Эпигенетика является примером основного механизма воздействия генов на окружающую среду, однако она не позволяет сделать вывод, являются ли эффекты окружающей среды аддитивными, мультипликативными или интерактивными. [13]
Новые исследования также выявили интерактивный эффект множества факторов окружающей среды. Например, ребенок с плохим окружением будет более чувствителен к плохому окружению во взрослом возрасте, что в конечном итоге приведет к более высоким показателям психологического дистресса. Это отражает трехстороннее взаимодействие: Ген x Окружающая среда x Окружающая среда. В том же исследовании предлагается использовать подход на протяжении всей жизни для определения генетической чувствительности к воздействиям окружающей среды в рамках психических заболеваний. [18]
Врачи заинтересованы в том, можно ли предотвратить заболевание, снизив воздействие экологических рисков. Некоторые люди являются носителями генетических факторов, которые придают восприимчивость или устойчивость к определенному заболеванию в определенной среде. Взаимодействие между генетическими факторами и стимулами окружающей среды приводит к фенотипу заболевания. [19] может принести значительную пользу общественному здравоохранению . Использование взаимодействия генов и окружающей среды для предотвращения или лечения заболеваний [20]
Реакция человека на лекарство может быть результатом различных взаимодействий генов и окружающей среды. [19] Таким образом, клиническое значение фармакогенетики и взаимодействия генов с окружающей средой обусловлено возможностью того, что геномика, наряду с информацией об окружающей среде, позволит более точно прогнозировать реакцию человека на лекарство. Это позволит врачам более точно подбирать определенный препарат и дозировку для достижения терапевтического ответа у пациента, сводя при этом к минимуму побочные эффекты и побочные реакции на лекарство . [21] Эта информация также может помочь предотвратить расходы на здравоохранение, связанные с побочными реакциями на лекарства и неудобным назначением лекарств пациентам, которые, скорее всего, не будут на них реагировать. [19]
Аналогичным образом человек может реагировать на другие стимулы, факторы или проблемы окружающей среды по-разному в зависимости от конкретных генетических различий или аллелей. Эти другие факторы включают диету и определенные питательные вещества в рационе, физическую активность, употребление алкоголя и табака, сон (время сна, продолжительность) и любое из ряда воздействий (или экспозомов ), включая токсины, загрязняющие вещества, солнечный свет (северная широта). – к югу от экватора), среди множества других. Диету, например, можно изменить, и она оказывает значительное влияние на множество кардиометаболических заболеваний, включая сердечно-сосудистые заболевания, ишемическую болезнь сердца, ишемическую болезнь сердца, диабет 2 типа , гипертонию , инсульт , инфаркт миокарда и неалкогольную жировую болезнь печени. В клинике обычно оцениваемые риски этих состояний включают липиды крови (триглицериды, ЛПВП, ЛПНП и общий холестерин), гликемические характеристики (глюкоза и инсулин в плазме, HOMA-IR, функция бета-клеток как HOMA-BC), антропометрические показатели ожирения (ИМТ). /ожирение, ожирение, масса тела, окружность талии, соотношение талии к бедрам), сосудистые показатели (диастолическое и систолическое артериальное давление) и биомаркеры воспаления. Взаимодействия гена и окружающей среды могут модулировать неблагоприятные эффекты аллели, которая приводит к повышенному риску заболевания, или могут усугублять отношения генотип-фенотип и повышать риск, что часто называют нутригенетика . [22] Доступен каталог генетических вариантов, которые связаны с этими и родственными кардиометаболическими фенотипами и модифицируются общими факторами окружающей среды. [23]
И наоборот, исследование заболеваний с использованием рака молочной железы, диабета 2 типа и ревматоидного артрита показывает, что включение взаимодействий GxE в модель прогнозирования риска не улучшает идентификацию риска. [24]
У дрозофилы : Классический пример взаимодействия гена и окружающей среды был продемонстрирован на дрозофиле Гуптой и Левонтином в 1981 году. В своем эксперименте они продемонстрировали, что среднее количество щетинок у дрозофилы может меняться при изменении температуры. Как видно на графике справа, разные генотипы по-разному реагировали на изменение окружающей среды. Каждая линия представляет данный генотип, а наклон линии отражает изменение фенотипа (количества щетинок) с изменением температуры. У некоторых особей количество щетинок увеличивалось с повышением температуры, тогда как у других наблюдалось резкое уменьшение количества щетинок с повышением температуры. Это показало, что нормы реакции у этих мух не параллельны, что доказывает существование взаимодействия генов и окружающей среды. [25]
У растений: одним из очень интересных подходов к определению генотипа по стратегиям взаимодействия с окружающей средой является его использование при селекции сортов сахарного тростника, адаптированных к различным условиям окружающей среды. [26] В этой статье они проанализировали двадцать генотипов сахарного тростника, выращенных в восьми разных местах в течение двух сельскохозяйственных циклов, чтобы определить мега-среду, связанную с более высокой урожайностью тростника, измеряемой в тоннах тростника на гектар (TCH) и процентном содержании сахарозы (Pol% тростника) с использованием биплота. многомерные модели GEI. Затем авторы разработали новую стратегию для изучения обеих переменных урожайности в рамках двусторонней связанной стратегии, хотя результаты показали среднюю отрицательную корреляцию. С помощью коинерционного анализа удалось определить наиболее подходящие генотипы для обеих переменных урожайности во всех средах. [27] Использование этих новых стратегий, таких как коинерция в GEI, оказалось отличным дополнением к анализу AMMI и GGE, особенно когда повышение урожайности подразумевает несколько переменных доходности. Было собрано семь генетически различных растений тысячелистника и от каждого растения взято по три черенка. По одному черенку каждого генотипа высаживали на низкой, средней и высокой высоте соответственно. Когда растения созрели, ни один генотип не рос лучше всего на всех высотах, и на каждой высоте семь генотипов вели себя по-разному. Например, один генотип достиг наибольшего роста на средней высоте, но достиг лишь средней высоты на двух других высотах. Лучшие производители на низких и высоких высотах плохо росли на средней высоте. На средней высоте были получены худшие общие результаты, но все же были получены один высокий и два образца средней высоты. Высота над уровнем моря влияла на каждый генотип, но не в одинаковой степени и не одинаковым образом. [28] Популяцию сорго двуродительского на протяжении многих лет неоднократно выращивали в семи различных географических точках. Группе генотипов требуется одинаковое количество градусо-дней роста (GDD) для цветения во всех средах, в то время как другой группе генотипов для цветения требуется меньше GDD в определенных средах, но более высокий GDD в разных средах. Сложные закономерности времени цветения объясняются взаимодействием основных генов времени цветения ( Ma 1 , [29] Ма 6 , [30] FT , ELF3 ) и явный фактор окружающей среды — фототермическое время (PTT), отражающий взаимодействие между температурой и фотопериодом. [31]
Фенилкетонурия человека, (ФКУ) — генетическое заболевание вызванное мутациями гена, кодирующего определенный фермент печени. В отсутствие этого фермента аминокислота , известная как фенилаланин, не превращается в следующую аминокислоту биохимическим путем , и поэтому слишком много фенилаланина попадает в кровь и другие ткани. Это нарушает развитие мозга , что приводит к умственной отсталости и другим проблемам. ФКУ поражает примерно 1 из каждых 15 000 младенцев в США. Однако большинство больных младенцев не растут с нарушениями развития благодаря стандартной программе скрининга, используемой в США и других промышленно развитых странах. Новорожденным, у которых обнаружен высокий уровень фенилаланина в крови, можно посадить на специальную диету, не содержащую фенилаланин. Если их сразу же посадить на эту диету и продолжать ее придерживаться, эти дети смогут избежать тяжелых последствий ФКУ. [32] Этот пример показывает, что изменение окружающей среды (снижение потребления фенилаланина) может повлиять на фенотип определенного признака, демонстрируя взаимодействие гена и окружающей среды.
Однонуклеотидный полиморфизм rs1800566 в NAD(P)H-хинондегидрогеназе 1 (NQO1) изменяет риск астмы и общего повреждения легких при взаимодействии с загрязнителями NOx у людей с этой мутацией. [33] [34]
Функциональный полиморфизм промотора гена моноаминоксидазы А (МАОА) может смягчить связь между травмой в раннем возрасте и повышенным риском насилия и антисоциального поведения . Низкая активность МАОА является значительным фактором риска агрессивного и антисоциального поведения у взрослых, которые сообщают о виктимизации в детстве. У людей, подвергшихся насилию в детстве, но имеющих генотип, обеспечивающий высокий уровень экспрессии МАОА, с меньшей вероятностью развиваются симптомы антисоциального поведения. [35] Однако эти результаты следует интерпретировать с осторожностью, поскольку исследования генных ассоциаций сложных признаков печально известны тем, что их очень трудно подтвердить. [36]
В яйцах дрозофилы : Время развития яиц в зависимости от температуры В отличие от приведенных выше примеров, продолжительность развития яиц у дрозофилы в зависимости от температуры демонстрирует отсутствие взаимодействия генов и окружающей среды. На прилагаемом графике показаны параллельные нормы реакции для различных отдельных мух -дрозофил , показывая, что между двумя переменными нет взаимодействия гена и окружающей среды. Другими словами, каждый генотип одинаково реагирует на изменение окружающей среды, создавая схожие фенотипы. Для всех отдельных генотипов среднее время развития яиц уменьшается с повышением температуры. Окружающая среда влияет на каждый из генотипов одинаково предсказуемо. [25]
^ Грин А., Трихопулос Д. (2002). «Рак кожи». Адами Х., Хантер Д., Трихопулос Д. (ред.). Учебник эпидемиологии рака . Оксфорд: Издательство Оксфордского университета. стр. 281–300.
^ Перейти обратно: а б с Табери Дж., Гриффитс П.Е. (2010). «Исторические и философские перспективы поведенческой генетики и науки о развитии». В Худ К.Э., Халперн К.Т., Гринберг Г., Лернер Р.М. (ред.). Справочник по наукам о развитии, поведении и генетике . Уайли-Блэквелл. стр. 41–60.
^ Раттер, Майкл. (2006) Гены и поведение: объяснение взаимодействия природы и воспитания Оксфорд, Великобритания: Blackwell Publishers
^ Куна Ф., Хекман Джей-Джей (2010). «Глава 18: Экономически эффективные программы для детей раннего возраста в первом десятилетии: интеграция человеческого капитала». В Рейнольдсе А.Дж., Рольнике А., Энглунде М.М., Темпле Дж. (ред.). Инвестиции в нашу молодежь . Нью-Йорк: Издательство Кембриджского университета. стр. 381–414.
^ Грабе Х.Дж., Шван С., Малер Дж., Шульц А., Спитцер С., Фенске К., Аппель К., Барноу С., Наук М., Шомерус Г., Биффар Р., Росскопф Д., Джон У., Вольцке Х., Фрейбергер Х.Дж. (апрель 2012 г.). «Умерение депрессии у взрослых с помощью варианта промотора переносчика серотонина (5-HTTLPR), жестокого обращения в детстве и травматических событий у взрослых в общей выборке населения». Американский журнал медицинской генетики. Часть B. Нейропсихиатрическая генетика . 159Б (3): 298–309. дои : 10.1002/ajmg.b.32027 . ПМИД 22328412 . S2CID 21356506 .
^ Ри Р., Де Соуза-Виейра О, Диас А., Рамон М., Брицено Р., Джордж Дж., Чайлд М., Бальзано-Ногейра Л. (2016). «Взаимодействие генотипа и окружающей среды, мегасреда и методы сопряжения двух таблиц для исследований урожайности сахарного тростника в Венесуэле». Сахарная Техника . 18 (4): 354–364. дои : 10.1007/ s12355-015-0407-9 S2CID 18351550 .
^ Клаузен Дж., Кек Д., Хизи В.М. (1948). «Экспериментальные исследования природы видов. III. Реакция климатических рас Achillea на окружающую среду, Carnegie Inst Washington Publ 581»: 1–129. {{cite journal}}: Для цитирования журнала требуется |journal= ( помощь )
Arc.Ask3.Ru Номер скриншота №: dbb35f21ef897ae51b44aad02cb585e6__1706309460 URL1:https://arc.ask3.ru/arc/aa/db/e6/dbb35f21ef897ae51b44aad02cb585e6.html Заголовок, (Title) документа по адресу, URL1: Gene–environment interaction - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)