Эйхштеттизавр
| Эйхштеттизавр Временной диапазон:
| |
|---|---|

| |
| Тип и единственный экземпляр E. schroederi. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Заказ: | Чешуйчатый |
| Семья: | † Эйхштеттизавриды |
| Род: | † Эйхштеттизавр Кун, 1958 г. |
| Типовой вид | |
| † Eichstaettisaurus schroederi (Бройл, 1938)
| |
| Другие виды | |
| |
| Синонимы | |
|
Э. Шредери
| |
Eichstaettisaurus (что означает «ящерица Эйхштетта») — род ящериц . поздней юры и раннего мела Германии, Испании и Италии Имея уплощенную голову, ориентированные вперед и частично симметричные ступни и высокие когти, эйхштеттизавр имел множество приспособлений к образу жизни лазания, приближающихся к образу жизни гекконов . Типовой вид , E. schroederi , является одним из старейших и наиболее полных представителей Squamata , известный по одному экземпляру, происходящему из титонского возраста известняка Зольнхофен в Германии. Второй вид, E. gouldi , был описан по другому скелету, найденному в горах Матезе в Италии. Несмотря на то, что он был очень похож на E. schroederi , он жил гораздо позже, на альбском этапе . Окаменелости обоих видов демонстрируют исключительную сохранность благодаря отложению в с низким содержанием кислорода морской среде .
в 1938 году как вид рода Ardeosaurus Первоначально названный Фердинандом Бройли , E. schroederi имел запутанную таксономическую историю. Первоначально его считали родственником ночных ящериц , пока исследования 1960-х годов не идентифицировали его и ардеозавра как гекконов. Однако филогенетический анализ, проведенный в 1990-х годах, позволил предположить, что он был ближе к предку всех чешуйчатых . Различные противоречивые позиции были обнаружены до появления анализа, включающего больше видов и более точные данные, который определил E. schroederi как близкого родственника гекконов в Gekkonomorpha . Однако положение Ardeosaurus по отношению к E. schroederi остается неопределенным: в некоторых анализах один вид, A. digitatellus , был обнаружен как его близкий родственник, но другой вид, A. brevipes, оказался более отдаленным родственником.
Открытие и присвоение имени
[ редактировать ]Э. Шредери
[ редактировать ]
В 1938 году Фердинанд Бройли описал прекрасно сохранившийся экземпляр ящерицы, сохранившийся вверх дном, из отложений скал юрского возраста в муниципалитете Винтерсхоф, Айхштет , Германия. Экземпляр поступил в Баварскую государственную коллекцию палеонтологии и исторической геологии (BSPG), где его подготовил Л. Хаймерль. Он состоит из почти полного скелета, хотя и с раздробленным черепом, хвоста, у которого отсутствует хвост после пятого позвонка (вероятно, в результате ампутации и замены хрящом ), и нескольких позвонков, сохранившихся лишь в виде отпечатков (но их наличие все же можно было установить). проверено соответствующими ребрами). к Также существует контрплита образцу. [ 1 ] Сегодня известно, что отложения горных пород принадлежат к формации Альтмюльталь , а образец и его контрплита хранятся под инвентарными номерами BSPG 1937 I 1a и 1b. [ 2 ]
Хотя он не смог изучить его лично, Бройли отметил из Кристиана Эриха Германа фон Мейера описания типового экземпляра Ardeosaurus brevipes , что он имеет сходство с этим экземпляром, но также и что они имеют важные различия в морды и лобных костей. форме , а также подсчет позвонков. Однако он не решился создать новый род из-за сохранности черепа. Таким образом, он отнес его к роду Ardeosaurus с оговорками как новый вид, A.? schröderi , названный в честь своего коллеги Й. Шрёдера. [ 1 ] (Текущее видовое название - schroederi : согласно статье 32.5.2.1 Международного кодекса зоологической номенклатуры в немецких названиях , умлауты опубликованных до 1985 года, должны быть удалены и заменены буквой «е», вставленной в качестве следующей буквы.) [ 3 ]
Роберт Хоффстеттер формально признал различие между А.? schroederi и brevipes в 1953 году и установили новое родовое название Broilisaurus A. . Однако Оскар Кун уже дал стегоцефалической такое же имя амфибии в 1938 году; таким образом, в 1958 году он установил еще одно родовое название вида — Eichstättisaurus . (Опять же, согласно ICZN, нынешнее родовое название — Eichstaettisaurus .) В 1963 году Маргерит Кокуд-Мишель высказала мнение, что E. schroederi действительно представляет тот же вид, что и Homoeosaurus digitatellus , который ранее назывался нетиповым видом Homoeosaurus. в 1914 году Н. М. Гриер. Таким образом, она создала новую комбинацию Eichstaettisaurus digitatellus . [ 4 ] [ 5 ] [ 6 ]
В отличие от Кокуд-Мишеля, Хоффстеттер идентифицировала digitatellus как члена Ardeosaurus, отдельного от Eichstaettisaurus, в 1964 году. Хотя Кокуд-Мишель знала о статье Хоффстеттера, в 1965 году она повторила свой аргумент и не предприняла никаких попыток обратиться к Хоффстеттеру. Хоффстеттер ответил в 1966 году, назвав синоним Кокуде-Мишеля «невыносимым». что это повлечет за собой пересмотр диагностических характеристик Eichstatettisaurus Он отметил , до такой степени, что он будет практически неотличим от Ardeosaurus , что противоречит ее на семейном уровне . разделению [ 6 ] сохранялись В последующей литературе Eichstaettisaurus schroederi и Ardeosaurus digitatellus как отдельные, хотя им уделялось мало внимания, пока Тьяго Симойнс и его коллеги не опубликовали переописание обоих в 2017 году. [ 2 ]
Э. голди
[ редактировать ]
В течение многих лет Eichstaettisaurus был известен по единственному экземпляру E. schroederi , пока останки мелового возраста не были обнаружены в других частях Европы. В 2000 году Сьюзан Эванс и ее коллеги описали небольшой экземпляр ящерицы в , Испания, берриасско - валанжинского местности Монсек в Каталонии возраста . Несмотря на плохую сохранность, экземпляр явно отличался от более распространенной в этой местности ящерицы Мейазавра ; Эванс и его коллеги предположили, что он имеет сходство с Eichstaettisaurus . [ 7 ]
В 2004 году Эванс и его коллеги сообщили о еще более молодых остатках Eichstaettisaurus , которые произошли от альбского возраста Pietraroja Plattenkalk в местности Pietraroia , которая расположена в горах Матезе на юге Италии. Образец, каталогизированный как MPN 19457, образовал типовой образец нового вида, E. gouldi , который они назвали в честь покойного биолога Стивена Джея Гулда ; это скелет, сохранившийся в перевернутом виде, включающий череп, части позвоночного столба и конечностей. Второй экземпляр представляет собой раздробленный череп, который сохранился как часть содержимого кишечника неопределенного ринхоцефала, каталогизированного как MPN A01/82. Оба экземпляра хранятся в Музее палеонтологии Неаполитанского университета (MPN). [ 4 ]
Описание
[ редактировать ]Эйхштеттизавр был относительно небольшим животным. У E. schroederi длина отверстия морды (измеренная от кончика морды до отверстия клоаки ) составляла 94 мм (3,7 дюйма), [ 2 ] в то время как E. gouldi имел длину морды и отверстия 56 мм (2,2 дюйма). [ 4 ]
Череп
[ редактировать ]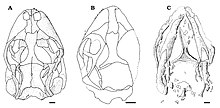
Эйхштеттизавра можно узнать по уплощенному черепу с короткой, тупой и закругленной мордой. У E. schroederi были большие ноздри. Кости морды, несущие зубы, предчелюстная и верхняя челюсти , вероятно, были соединены полосой мягких тканей у E. schroederi . было 6 или 7 зубов На предчелюстной кости у E. gouldi , но их количество у E. schroederi неизвестно; на верхней челюсти было примерно 22 зуба у E. schroederi и не менее 30 у E. gouldi . Зубы обоих видов были мелкими, узкими, коническими, одностворчатыми ; у них была имплантация плевродонта , то есть они были слиты с внутренней стороной челюстной кости. Как и у ардеозавра , у обоих видов отсутствовала слезная кость , обычно являющаяся частью глазницы. Также в глазнице постфронтальная и заглазничная кости были отдельными в отличие от ардеозавра , а задняя проекция последних костей была относительно широкой. У E. schroederi контакт между этими костями был дальше вперед, чем у большинства ящериц. Чешуйчатые кости были тонкими и выступали вниз, чтобы соответствовать кости квадратные , как у большинства ящериц; однако, в отличие от Iguania и Teiidae , у них не было выступов вверх. У современных гекконов отсутствуют заглазничные и чешуйчатые кости, а также у них относительно более короткие скуловые кости, чем у Eichstaettisaurus . На нёбе эктоптеригоидные E. schroederi кости перекрывали крыловидные кости сзади. [ 2 ] [ 4 ]
В отличие от A. brevipes , у E. schroederi отсутствовал костный орнамент и остеодермы на крыше черепа . У обоих видов лобные кости срослись и между глазницами стали уже, чем спереди. Нисходящие выступы лобных костей, известные как субольфакторные отростки, были хорошо развиты у обоих видов, но встречались по средней линии только у E. schroederi . Шов , между лобной и теменной костями был выпуклым у E. schroederi и вогнутым у E. gouldi , в отличие от Ardeosaurus у которого он был прямым. E. gouldi и большинство других ящериц имеют сросшиеся теменные кости, но у E. schroederi они были парными и несросшимися . Теменные кости обоих видов не имели выступов назад и затылочных ямок . У E. schroederi внешние края теменных костей загнуты внутрь, а задние выступы, известные как надвисочные отростки, были короткими, широко расставленными и имели впадины. Также у E. schroederi черепной коробки присутствовала пара гребней на надзатылочной кости , которые, вероятно, были отпечатаны полукружные каналы черепа из-за снижения окостенения . [ 2 ] [ 4 ]
Позвонки и ребра
[ редактировать ]По сравнению с большинством игуанских, геккотанских и сцинкоморфных ящериц, которые обычно имеют 24–26 позвонков перед крестцом (или бедром), у E. schroederi их было 31, включая 7 шейных позвонков и примерно 24 задних позвонка . [ 2 ] У E. gouldi было 6–7 шейных позвонков и неизвестное количество задних позвонков. [ 4 ] У E. gouldi второй ( ось ) и третий шейные позвонки были соединены со следующими элементами, известными как интерцентры. [ 4 ] У E. schroederi у большинства шейных позвонков отсутствовали шейные ребра , за исключением двух последних. Выступающие вверх нервные отростки на задних позвонках были маленькими, а у последних нескольких задних позвонков не было ребер в венах поясничных позвонков . Зигосфен-зигантральные сочленения имеются на задних позвонках обоих видов. Ребра были одноглавыми и имели круглые сочленяющиеся поверхности. [ 2 ] У E. gouldi центры (вогнутыми с обоих концов (основные тела) хвостовых позвонков были широкими, цилиндрическими, амфицелозными ) и имели сильно развитые поперечные отростки . [ 4 ]
Конечности и пояса конечностей
[ редактировать ]
У эйхштеттизавра был хорошо развит акромиальный отросток, или прикрепление ключицы на лопатке (лопатке) . Ключицы E. schroederi не выгибались вперед, в отличие от большинства представителей Autarchoglossa , а также не имели выступов назад. Как и у Ardeosaurus , у обоих видов Eichstaettisaurus были относительно толстые передние конечности. [ 2 ] [ 4 ] У E. schroederi локтевой локтевой отросток кости был хорошо развит. [ 2 ] У E. gouldi кости запястья , вероятно, не были полностью окостеневшими. [ 4 ] Пятипалая рука E. schroederi имела на каждом пальце по 2, 3, 4, 5 и 3 фаланги (кости пальцев); предпоследние фаланги на кончиках были относительно длинными. [ 2 ] У E. gouldi , по-видимому, было 4 фаланги на четвертом пальце, а когти были короткими, глубокими и острыми, с хорошо развитыми сгибательными бугорками . [ 4 ]
В тазу расширение E. schroederi лобковых костей в сросшийся лобковый фартук было относительно узким; на лобке также отсутствовали выступы вперед, известные как лобковые бугорки . Седалищные кости были увеличены и широко соприкасались друг с другом, а заднее расширение лопатки подвздошной кости было длинным. Оба вида Eichstaettisaurus имели толстые задние конечности, схожие по длине с передними конечностями. Бедренная кость имела прямой стержень, в отличие от большинства других ящериц, у которых она имела сигмовидную форму . На стопе первая плюсневая кость была немного короче второй-четвертой плюсневых костей. Необычно то, что пятая плюсневая кость была намного короче и крючковатой. У E. schroederi стопа была пятипалой, как и руки, с 2, 3, 4, 5 и 4 фалангами на каждом пальце, а предпоследние фаланги и когти были одинаковыми. E. schroederi разделял две особенности когтей на ногах с современными токайскими гекконами : когти имели расширенные нижние концы; а сочленения между фалангами представляли собой простые вогнуто-выпуклые суставы, а не двумыщелковый (т.е. основанный на двух закругленных выступах на концах фаланг), в отличие от большинства других ящериц. [ 2 ] [ 4 ]
Классификация
[ редактировать ]
Филогенетическая Ardeosaurus история Eichstaettisaurus была бурной, как и история , что было следствием как неполных данных, так и его примитивной формы. [ 2 ] [ 8 ] фон Мейер не указал местонахождение ардеозавра , когда называл род; Рихард Лидеккер и Карл Альфред фон Циттель поместили его и гомоеозавра в группу ринхоцефалий, барон Франц Нопса поместил ардеозавра как близкого родственника сцинков , а Чарльз Кэмп поместил его рядом с гекконидами. [ 9 ] [ 10 ] [ 11 ] Основываясь на хорошо развитых краях глазниц и надвисочных отверстиях на черепе у типового экземпляра E. schroederi , Бройли согласился с выводом Нопца в 1938 году. В пределах Scincomorpha он отметил, что амфицельные позвонки Ardeosaurus (включая E. schroederi ) больше всего напоминал таковой у Xantusiidae (ночных ящериц), как и у большинства других сцинков, имеются процельные (выпуклые на задней поверхности) позвонки. [ 1 ]
Выделив schroederi в род Broilisaurus , Хоффстеттер сохранил его в составе Xantusiidae в 1953 году, но заметил, что его череп странно напоминал череп гекконов. Поскольку ксантузииды и гекконы тогда были помещены в две основные признанные ветви ящериц, Autarchoglossa и Ascalabota , Хоффстеттер предположил, что они были близки к предковой группе ящериц. [ 5 ] Переименовав его в Eichstaettisaurus , Кун основал семейство Eichstaettisauridae, все еще входящее в состав Xantusiidae. В 1961, а затем в 1963 году Кокуд-Мишель согласился с Кэмпом в том, что Ardeosauridae и Eichstaettisauridae на самом деле были гекконами, а не ксантузиидами, на основании серьезных различий в посткраниальном скелете, и отнес обоих к Gekkonomorpha . [ 5 ] Хоффстеттер принял это определение в 1966 году. [ 6 ] как и Ричард Эстес в 1983 году. [ 2 ]
Однако филогенетический анализ , проведенный Виктором Рейносо в 1996 году (не опубликован), Эвансом и Луисом Хавьером Барбадилло в 1998 и 1999 годах, а также Эвансом и Дэниелом Чюре в 1998 году, показал, что и Eichstaettisaurus , и Ardeosaurus были гораздо более базальными (раннерасходящимися) чешуйчатыми . Было обнаружено, что наряду с Bavarisaurus , Scandensia и Hoyalacerta они не входят в кронную группу Squamata. Это сделало бы их стволово-чешуйчатыми, т.е. не потомками общего предка всех ныне живущих чешуйчатых. [ 2 ] [ 12 ] [ 13 ] Однако в анализе 1999 года Эванс и Барбадилло признали, что положение Eichstaettisaurus было нестабильным из-за отсутствия данных (только 44% характеристик, использованных в их анализе, можно было закодировать для него), и одно из их филогенетических деревьев поместило его как сестринская группа среди Scleroglossa корончатых чешуек. [ 14 ] Эванс и его коллеги в 2004 году обнаружили, что E. gouldi является сестринской группой E. schroederi , и восстановили одинаковое положение для обоих. [ 4 ]
Последующий филогенетический анализ, проведенный Эвансом и его коллегами, выявил множество противоречивых позиций для Eichstaettisaurus и Ardeosaurus . В 2005 году Эванс и Юань Ван провели два анализа: один на основе анализа Майкла Ли 1998 года, а другой на основе анализа Эванса и Чура 1998 года, но они добавили Далингозавра , Дорсетизавра и Парвираптора к обоим . Было обнаружено, что Eichstaettisaurus является сестринской группой группы, состоящей из Xantusiidae, Gekkonidae, Pygopodidae и Amphisbaenia , при этом Ardeosaurus близок к Lacertidae и Teiioidea ; другой объединял Eichstaettisaurus с Dalinghosaurus и Xenosauridae в Anguimorpha , с Ardeosaurus как сестринской группой Iguania. [ 15 ] После того, как Эванс, Ван и Чун Ли добавили тому анализу, было обнаружено, что Yabeinosaurus и Parviraptor образуют Eichstaettisaurus сестринскую группу Scleroglossa Ябейнозавра , , Ardeosaurus к , же и Scandensia образуют группу среди Scleroglossa. [ 16 ]
Филогенетический анализ 2006 года, проведенный Джеком Конрадом и Марком Нореллом, также обнаружил, что Eichstaettisaurus максимально близок к Scleroglossa. [ 17 ] В 2008 году Конрад опубликовал большой филогенетический анализ чешуйчатых, включающий 222 таксона и 363 характеристики, в котором была предпринята попытка решить проблему неполной выборки таксонов (т.е. недостаточного количества включенных таксонов), вызывающей филогенетическую нестабильность. Чтобы включить Eichstaettisaurus , он объединил данные E. schroederi и E. gouldi . Он обнаружил, что и Eichstaettisaurus , и Ardeosaurus , наряду с Bavarisaurus , были частью стволовой группы Scleroglossa. Наряду с кроновой группой Scleroglossa они входили в состав Scincogekkonomorpha . [ 18 ] Арнау Болет и Эванс провели два анализа на основе анализа Конрада для описаний Pedrerasaurus и Jucaraseps в 2010 и 2012 годах и восстановили аналогичные позиции для Eichstaettisaurus . Однако первый содержал политомию , поскольку отношения между Игуанией, Педреразавром , группой, состоящей из Ардеозавра , Ябейнозавра и Сакуразавра , и всеми другими чешуйчатыми не могли быть решены. [ 19 ] [ 20 ]
Жак Готье и его коллеги опубликовали в 2012 году еще один крупный филогенетический анализ, содержащий 192 таксона и 976 характеристик. Как и Конрад, они также объединили данные E. schroederi и E. gouldi . Они обнаружили более производное положение Eichstaettisaurus как части стволовой группы Gekkota вместе с безымянным экземпляром AMNH FR 21444; в частности, они отметили, что пропорции его конечностей сильно напоминали геккотанов. [ 8 ] После добавления Ardeosaurus digitatellus и перекодирования Eichstaettisaurus , чтобы включить только данные по E. schroederi , Симойнс и его коллеги обнаружили E. schroederi в аналогичном положении, хотя и в политомии с A. digitatellus и AMNH FR 21444. Они определили ряд признаков геккотана. : парные теменные кости; закрытая меккелева борозда на нижней челюсти; маленькие конические верхние зубы; короткая тупая морда и уплощенный череп; эктоптеригоиды перекрывают крыловидные кости; две крыловидные кости и небные кости широко разделены; широкий шов, отделяющий крыловидные и небные кости друг от друга; и поясничноподобные позвонки. [ 2 ]
В 2018 году Матеуш Таланда добавил Ardeosaurus brevipes к анализу Готье и его коллег (но не включил поправки Симойнса и его коллег). Он нашел Eichstaettisaurus в том же месте рядом с Norellius , но также обнаружил A. brevipes как сцинкоморфа, тесно связанного со сцинками (в отличие от стеблевого геккотанского положения A. digitatellus ). Несмотря на разное филогенетическое положение, Таланда заметил, что эти два вида различаются только шириной теменных костей. Он предположил, что на эти результаты могли повлиять неполные данные из-за плохой сохранности A. digitatellus , и поэтому он сохранил их в роде Ardeosaurus . [ 21 ] Другой анализ, проведенный Симойнсом, Таландой и коллегами в 2018 году, на этот раз с использованием пересмотренных данных Симойнса и коллег, обнаружил аналогичное положение для E. schroederi , но более базальное для A. brevipes в Autarchoglossa. [ 22 ] В 2019 году Владимир Алифанов считал Eichstaettisaurus и Ardeosaurus сцинкоморфами, близкими к Carusia , но не к Xenosauridae, но не проводил филогенетический анализ. [ 23 ]
Ниже филогенетические деревья из двух основных филогенетических анализов, включая Eichstaettisaurus : одно основано на исследовании Конрада (Bolet & Evans, 2012, с отношениями внутри Scleroglossa из Conrad, 2008), показаны [ 18 ] [ 20 ] и один основан на исследовании Готье и его коллег (Simões and коллеги, 2018). [ 22 ]
Топология A: Болет и Эванс (2012) со Склероглоссой из Конрада (2008) [ 18 ] [ 20 ]
| Чешуйчатый |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Топология B: Simões et al. (2018) [ 22 ]
Палеобиология
[ редактировать ]
Современные гекконы необычны среди ящериц тем, что пальцы их конечностей относительно симметричны по длине и растопырены по широкой дуге; напротив, у других ящериц пальцы обычно почти параллельны друг другу, особенно на ступнях. Рисунок, наблюдаемый у гекконов, облегчает захват, когда тело находится в различных положениях, поскольку он распределяет липкие щетинки (щетинки) на подушечках ног, позволяя при этом первым и последним пальцам располагаться напротив друг друга. [ 24 ] [ 25 ] Eichstaettisaurus schroederi обладал двумя характеристиками, которые способствовали симметрии стопы: его первая плюсневая кость была длиннее третьей плюсневой кости (при этом третья плюсневая кость была всего в 1,41 раза длиннее первой, в отличие от 2 раз у типичной ящерицы), а четвертая плюсневая кость была длиннее первой. плюсневая кость была относительно короткой. В 2017 году Симойнс и его коллеги заметили, что у E. schroederi более сильная симметрия стопы, чем у Ardeosaurus digitatellus , и пришли к выводу, что ступни обоих, вероятно, были направлены дальше вперед, чем у других ящериц. Это также подтверждается отсутствием двумыщелковых сочленений в фалангах, которые представляют собой адаптацию стопы, обращенной наружу, которая препятствует смещению в горизонтальной плоскости, но обеспечивает сгибание в вертикальной плоскости. [ 2 ]
Симойнс и его коллеги также определили несколько других характеристик у E. schroederi , которые позволяют предположить, что сканирующий (лазающий) образ жизни современных гекконов возник раньше, чем считалось ранее. Его высокие когти и удлиненные кончики фаланг на пальцах — обе черты, которые тесно коррелируют со сканирующим образом жизни ящериц. [ 26 ] [ 27 ] [ 28 ] [ 29 ] [ 30 ] В частности, высокие когти могли иметь более длинные рычаги для сухожилий сгибателей, которые их втягивают, тем самым улучшая их силу захвата. [ 31 ] Между тем, его относительно короткие конечности и уплощенное тело могли улучшить способность лазать за счет понижения центра тяжести, как это было предложено для токайского геккона. [ 31 ] но эта особенность может не коррелировать со скандинавским образом жизни. [ 32 ] Наконец, его конечности одинаковой длины могут иметь улучшенный захват, как у острорылой скальной ящерицы ; Значение этого признака может быть уменьшено, учитывая клейкие подушечки пальцев гекконов и отсутствие корреляции между соотношением длины конечностей и скансориальностью у Lacertidae . [ 2 ] [ 32 ]
В 2004 году Эванс и его коллеги также обсуждали образ жизни E. gouldi с учетом пропорций его тела. Они указали на предыдущее исследование о том, как различия между пропорциями среди видов снежных сцинков ( Carinascincus ) коррелировали с использованием среды обитания: короткое туловище и длинные задние конечности коррелировали со скалолазанием (у крупных видов) и лазаньем по деревьям (у мелких видов), в то время как длинное туловище и более короткие задние конечности коррелировали с наземным проживанием. [ 33 ] Эванс и его коллеги обнаружили, что E. gouldi по своим пропорциям ближе всего к наземным видам, которые имеют низкую скорость бега и относительно плохо лазают. Тем не менее они признали, что плоские тела и высокие когти E. gouldi были приспособлены для цепляния за шероховатые поверхности. Они предложили гибридный образ жизни E. gouldi : медленно передвигающаяся наземная ящерица, способная лазить по камням и прятаться в расщелинах от хищников, таких как ринхоцефалы. [ 4 ]
Палеоэкология
[ редактировать ]Э. Шредери
[ редактировать ]
Горные породы в Винтерсхофе, из которых образовался единственный известный экземпляр E. schroederi , являются частью известняков Зольнхофена на юге Германии, которые хорошо известны своими окаменелостями исключительной сохранности. [ 2 ] Исторически стратиграфия известняков Зольнхофена была предметом значительной путаницы, причем путаницу усугубляли разнообразие сред и моделей отложения. [ 34 ] Сейчас считается, что они состоят из нескольких геологических формаций группы Вайсюра : в самом строгом смысле они состоят из формации Альтмюльталь к северо-западу от Ингольштадта (включая местности вокруг Айхштетта) и формации Пейнтен к северо-востоку от Ингольштадта. В пределах формации Альтмюльталь карьер Винтерсхоф является частью пачки Верхний Айхштетт, которая с точки зрения аммонитов биостратиграфии залегает в горизонте Euvirgalithacoceras eigeltingense -β между Lithacoceras riedense и Hybooticeras hybonotum подзонами . [ 35 ] [ 36 ] Отложения формации Альтмюльталь, которые были датированы нижним титонским этапом юрского ) , периода , состоят из платтенкалька (очень мелкозернистого ила на основе известняка, такого как микрит образующего ровные тонкие слои толщиной около 1 сантиметра (0,39 дюйма). толстые, на которых обычно отсутствуют признаки биотурбации (нарушения со стороны живых организмов). [ 37 ] [ 38 ] [ 39 ]
В течение титона платтенкалк формации Альтмюльталь откладывался в океанических бассейнах (так называемых «ваннен») в теплом мелководном море, окружающем архипелаг . [ 39 ] Эти бассейны, которые, возможно, были лагунами , имели палеошироту примерно 34° северной широты и располагались на северной окраине океана Тетис . Окаменелости донных животных, таких как хрупкие звезды и брюхоногие моллюски, практически отсутствуют, что позволяет предположить, что условия на морском дне были негостеприимны для жизни; это могло быть вызвано одним из нескольких факторов, включая повышенную соленость , истощение кислорода или накопление токсичного сероводорода . [ 38 ] [ 40 ] Эти условия были ответственны за исключительную сохранность окаменелостей, таких как E. schroederi . [ 41 ] которые преимущественно переносились бы с окрестных земель во время сильных дождей и штормов. [ 42 ] На суше отсутствие папоротников и преобладание адаптированных к засухе хвойных пород позволяют предположить, что климат был полузасушливым . [ 43 ]

И Ardeosaurus brevipes , и A. digitatellus были обнаружены в Винтерсхофе вместе с E. schroederi . [ 2 ] наряду с Homoeosaurus maximiliani и ринхоцефалическим Pleurosaurus goldfussi . [ 5 ] Из Винтерсхофа также известны птерозавр Rhamphorhynchus muensteri , [ 44 ] крокодилообразный , bavaricus Alligatorellus [ 45 ] и различные водные животные: рыба Anaethalion angustus , [ 46 ] Аскалабос воитии , [ 47 ] Аспидоринхус остролистный , [ 48 ] Белоностомус спираеноидес , [ 49 ] Катурус гигантский [ 50 ] круговой гироскоп , [ 51 ] Macrosemius rostratus , Palaeomacrosemius thiollieri , [ 52 ] удлиненный поэтому [ 53 ] и Зандфуро тишлингери ; [ 54 ] акула -ангел Pseudorhina alifera ; [ 55 ] кальмароподобные колеоиды Acanthoteuthis проблематика , [ 56 ] Белемнотеутис майри , [ 57 ] и Plesioteuthis prisca ; [ 58 ] криноид Saccocoma tenella , очень распространенный в отложениях Зольнхофена; [ 59 ] креветки Дуса рески ; [ 60 ] и мечехвоста Mesolimulus walchi . [ 61 ] В близлежащих карьерах был найден айхштетский экземпляр птичьего Archeopteryx динозавра Lithographica . [ 62 ] и птерозавры Aerodactylus scolopaciceps , Germanodactylus cristatus и, возможно, Cycnorhamphus . [ 63 ]
Э. голди
[ редактировать ]
Местонахождение Пьетрароя или Пьетрароя, откуда известны экземпляры E. gouldi , образовано обнажением платтенкалька на вершине скального склона («Чивита ди Пьетрароя»), расположенного на восточной окраине гор Матезе. Эти горы не начали формироваться до миоцена , причем большая часть тектонического поднятия , которое привело их к нынешней высоте, произошла между плиоценом и плейстоценом . На этом участке выделяются два четко выраженных пласта платтенкалька; нижний слой более крупнозернистый и в основном лишен окаменелостей, а верхний слой более илистый и является источником большинства окаменелостей. Два пласта разделены лагунными известняками. Платтенкалк образован слоями пакстоуна и грейнстоуна толщиной 2–25 см (0,79–9,84 дюйма), которые чередуются со слоями аргиллита , вакстоуна и мергеля ; для верхнего слоя характерно также присутствие спикул губок , кремней , битумов и копролитов . [ 4 ] [ 64 ]
По фораминиферам местонахождение Пиетаройя датировано альбской эпохой раннего мела. За это время платтенкальк Пьетрарои должен был заложиться на мелководной карбонатной платформе недалеко от небольшого острова. Первоначально предполагалось, что среда осадконакопления представляла собой лагуну. [ 65 ] но его переинтерпретировали как подводный канал, идущий на восток , который постепенно заполнялся в аптском периоде на основании закономерностей в расположении окаменелостей, водных потоков и переноса отложений. Отсутствие биотурбации также указывает на бескислородную среду, возможно, связанную с глобальным океаническим бескислородным явлением . Некоторые окаменелости хорошо сохранились, тогда как другие были почти уничтожены, что позволяет предположить, что они произошли из разных мест, а наземные животные, такие как E. gouldi, были занесены в канал потоками пресной воды. [ 4 ] [ 64 ] Климат был от тропического до субтропического . [ 65 ]

Помимо E. gouldi MPN A01/82, другие лепидозавры из Пьетрарои включают ящериц Chometokadmon fitzingeri Costasaurus rusconi. ринхоцефала и и [ 4 ] [ 66 ] [ 67 ] Самая известная окаменелость из Пьетрарои — это окаменелость компсогнатного динозавра Scipionyx samniticus , у которого сохранились мягкие ткани, включая внутренние органы. [ 68 ] Другие рептилии включают два экземпляра крокодилообразных, принадлежащих Pietraroiasuchus ormezzanoi . [ 69 ] и альбанерпетонтидная амфибия Celtedens megacephalus . [ 70 ] Из Пьетрарои известно множество рыб, наиболее распространенным из которых является пикнодонтид, ранее идентифицированный как Coelodus costae , но теперь известный как Gregoriopycnodus bassanii . [ 71 ] Другие включают Anaethalionrobustus , Belonostomus crassirostris , Caeus leopoldi , Cavinichthys pietrarojae , Chirocentrites coroninii , родственники Diplomystus brevissimus и Elopopsis fenzii , Hemieloposis gibbus , Ionoscopus petrarojae , Italophiopsis derasmoi , вид Lepidotes . , Notagogus pentlandi , Pleuropholis decastroi , Propterus scacchii , и Sauropsidium laevissimum [ 72 ] [ 73 ] [ 74 ] [ 75 ] [ 76 ] К хрящевым рыбам относились акула-ангел Phorcynis и скат Rhinobatus obtusatus . [ 72 ] К беспозвоночным относились рудисты , двустворчатые моллюски , морские улитки рода Ninea , креветки и морские звезды. [ 64 ] [ 65 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Бройли, Ф. (1938). «Новая находка? Ардеозавра Х. фон Мейера» [Новая находка? Ардеозавр Х. фон Мейер] (PDF) . Протоколы заседаний отделения математических и естественных наук Баварской академии наук в Мюнхене : 97–114.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Симойнс, ТР; Колдуэлл, штат Вашингтон; Нидам, РЛ; Хименес-Уидобро, П. (2017). «Остеология, филогения и функциональная морфология двух юрских видов ящериц и ранняя эволюция скансориальности у гекконов». Зоологический журнал Линнеевского общества . 180 (1): 216–241. дои : 10.1111/zoj.12487 .
- ^ Сниттинг, Д.; Блом, Х. (2009). «Исправление названий таксонов, содержащих диакритические знаки, - примеры палеозойских позвоночных». Журнал палеонтологии позвоночных . 29 (1): 269–270. дои : 10.1080/02724634.2009.10010377 . S2CID 196603846 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Эванс, SE; Райя, П.; Барбера, К. (2004). «Новые ящерицы и ринхоцефалы из нижнего мела южной Италии» (PDF) . Acta Palaeontologica Polonica . 49 (3): 393–408.
- ^ Перейти обратно: а б с д Кокуд-Мишель, М. (1963). «Ринхоцефалы и ящеры из литографических известняков (верхняя юра) Западной Европы». Новые архивы Музея естественной истории Лиона . 7 : 143–185.
- ^ Перейти обратно: а б с Хоффстеттер, Р. (1966). «О родах и Eichstaettisaurus ( юры Франции » Ardeosaurus ) из верхней Reptilia , Sauria, Gekkonoidea . Бюллетень Геологического общества Франции . 8 (4): 592–595. дои : 10.2113/gssgfbull.S7-VIII.4.592 .
- ^ Эванс, SE; Руис, Алабама; Рей, Дж. (2000). «Ящерица из раннего мела (берриас-валанжин) Монсека, Каталония, Испания». Новогодний ежегодник геологии и палеонтологии - Трактаты . 215 : 1–15. дои : 10.1127/njgpa/215/2000/1 .
- ^ Перейти обратно: а б Готье, Дж.А.; Кирни, М.; Майзано, Дж.А.; Риппель, О.; Бельке, АБР (2012). «Сборка чешуйчатого древа жизни: перспективы фенотипа и летописи окаменелостей». Бюллетень Музея естественной истории Пибоди . 53 (1): 3–308. дои : 10.3374/014.053.0101 . S2CID 86355757 .
- ^ Лидеккер, Р. (1888). «Отряд Rhynchocephalia» . Каталог ископаемых рептилий и земноводных в Британском музее (Естественная история) . Том. 1. Лондон: Британский музей естественной истории. стр. 290–301. дои : 10.5962/bhl.title.61848 .
- ^ Нопца, Ф. (1908). «К познанию ископаемых ящериц» . Вклад в палеонтологию и геологию Австро-Венгрии и Востока . 21 :33–62.
- ^ Кэмп, К. (1923). «Классификация ящериц». Бюллетень Американского музея естественной истории . 48 : 289–481. hdl : 2246/898 .
- ^ Эванс, С.; Барбадилло, ЖЖ (1998). «Необычная ящерица (Reptilia: Squamata) из раннего мела Лас-Хойас, Испания» . Зоологический журнал Линнеевского общества . 124 (3): 235–265. дои : 10.1006/zjls.1997.0139 .
- ^ Эванс, SE; Чуре, округ Колумбия (1998). «Черепа ящериц-парамацеллодидов из юрской формации Моррисон в Национальном памятнике динозавров, штат Юта». Журнал палеонтологии позвоночных . 18 (1): 99–114. дои : 10.1080/02724634.1998.10011037 .
- ^ Эванс, SE; Барбадилло, ЖЖ (1999). «Короткоконечная ящерица из нижнего мела Испании» . Специальные статьи по палеонтологии . 60 : 73–85.
- ^ Эванс, SE; Ван, Ю. (2005). «Раннемеловая ящерица Далингозавр из Китая» (PDF) . Acta Palaeontologica Polonica . 50 : 725–742.
- ^ Эванс, SE; Ван, Ю.; Ли, К. (2005). «Раннемеловой род ящериц Yabeinosaurus из Китая: разгадка загадки». Журнал систематической палеонтологии . 3 (4): 319–335. дои : 10.1017/s1477201905001641 . S2CID 86410052 .
- ^ Конрад, Дж.Л.; Норелл, М. (2007). «Полный позднемеловой игуан (Squamata: Reptilia) из Гоби и идентификация новой клады игуан» . Новитаты Американского музея (3584): 1–47. doi : 10.1206/0003-0082(2007)3584[1:ACLCIS]2.0.CO;2 . hdl : 2246/5877 .
- ^ Перейти обратно: а б с Конрад, Дж.Л. (2008). «Филогения и систематика Squamata (Reptilia) на основе морфологии». Бюллетень Американского музея естественной истории . 310 : 1–182. дои : 10.1206/310.1 . hdl : 2246/5915 . S2CID 85271610 .
- ^ Болет, А.; Эванс, SE (2010). «Новая ящерица из раннего мела Каталонии (Испания) и мезозойские ящерицы Пиренейского полуострова». Меловые исследования . 31 (4): 447–457. дои : 10.1016/j.cretres.2010.06.002 .
- ^ Перейти обратно: а б с Болет, А.; Эванс, SE (2012). «Маленькая ящерица (Lepidosauria, Squamata) из нижнего мела Испании» . Палеонтология . 55 (3): 491–500. дои : 10.1111/j.1475-4983.2012.01145.x . S2CID 83645014 .
- ^ Таланда, М. (2018). «Исключительно сохранившийся юрский сцинк позволяет предположить, что диверсификация ящериц предшествовала фрагментации Пангеи». Палеонтология . 61 (5): 659–677. дои : 10.1111/пала.12358 . S2CID 134878128 .
- ^ Перейти обратно: а б с Симойнс, ТР; Колдуэлл, штат Вашингтон; Таланда, М.; Бернарди, М.; Пальчи, А.; Вернигора, О.; Бернардини, Ф.; Манчини, Л.; Нидам, РЛ (2018). «Происхождение чешуйчатых, выявленное среднетриасовой ящерицей из итальянских Альп». Природа . 557 (7707): 706–709. Бибкод : 2018Natur.557..706S . дои : 10.1038/s41586-018-0093-3 . ПМИД 29849156 . S2CID 44108416 .
- ^ Алифанов, В.Р. (2019). «Ящерицы семейств Eoxantidae, Ardeosauridae, Globauridae и Paramacellodidae (Scincomorpha) из апта-альба Монголии». Палеонтологический журнал . 53 (1): 74–88. дои : 10.1134/s0031030119010039 . S2CID 181824832 .
- ^ Рассел, AP (1986). «Морфологическая основа несущей способности сканеров токайского геккона (Reptilia: Sauria)». Канадский журнал зоологии . 64 (4): 948–955. дои : 10.1139/z86-144 .
- ^ Рассел, AP; Бауэр, AM; Лароия, Р. (1997). «Морфологические корреляты вторично симметричных стоп геккотанских ящериц». Журнал зоологии . 241 (4): 767–790. дои : 10.1111/j.1469-7998.1997.tb05747.x .
- ^ Зани, Пенсильвания (2000). «Сравнительная эволюция морфологии когтей и пальцев ящерицы и способности цепляться». Журнал эволюционной биологии . 13 (2): 316–325. дои : 10.1046/j.1420-9101.2000.00166.x . S2CID 62782034 .
- ^ Талли, MJ; Круз, ФБ; Херрел, А.; Ванхойдонк, Б.; Абдала, В. (2009). «Взаимодействие между морфологией когтей и использованием микросреды обитания у неотропических игуанских ящериц». Зоология . 112 (5): 379–392. дои : 10.1016/j.zool.2009.02.001 . hdl : 11336/78058 . ПМИД 19632100 .
- ^ Крэнделл, Кентукки; Херрел, А.; Саса, М.; Лосос, Дж.Б.; Осень, К. (2014). «Прилипать или хвататься? Совместная эволюция клейких подушечек пальцев и когтей у Anolis ящериц » . Зоология . 117 (6): 363–369. дои : 10.1016/j.zool.2014.05.001 . ПМИД 25069967 .
- ^ Кавана, КД; Шовал, О.; Уинслоу, BB; Алон, У.; Лири, BP; Кан, А.; Табин, CJ (2013). «Ошибка развития в эволюции фаланг» . Труды Национальной академии наук . 110 (45): 18190–18195. Бибкод : 2013PNAS..11018190K . дои : 10.1073/pnas.1315213110 . ПМЦ 3831474 . ПМИД 24151335 .
- ^ Ротье, PS; Брандт, Р.; Кольсдорф, Т. (2017). «Экологические ассоциации аутоподиальной остеологии неотропических гекконов». Журнал морфологии . 278 (3): 290–299. дои : 10.1002/jmor.20635 . ПМИД 28112828 . S2CID 24395310 .
- ^ Перейти обратно: а б Рассел, AP (1975). «Вклад в функциональный анализ стопы токайского геккона (Reptilia: Gekkonidae)». Журнал зоологии . 176 (4): 437–476. дои : 10.1111/j.1469-7998.1975.tb03215.x .
- ^ Перейти обратно: а б Ванхойдонк, Б.; Ван Дамм, Р. (1999). «Эволюционные взаимосвязи между формой тела и использованием среды обитания у ящериц-лацертид» (PDF) . Исследования в области эволюционной экологии . 1 : 785–805.
- ^ Мелвилл, Дж.; Суэйн, Р. (2000). «Эволюционные взаимосвязи между морфологией, продуктивностью и открытостью среды обитания у ящериц рода Niveoscincus (Scincidae: Lygosominae)» . Биологический журнал Линнеевского общества . 70 (4): 667–683. дои : 10.1111/j.1095-8312.2000.tb00222.x .
- ^ Швайгерт, Г. (2007). «Биостратиграфия аммонитов как инструмент для датирования литографических известняков верхней юры из Южной Германии – первые результаты и открытые вопросы». Новогодний ежегодник геологии и палеонтологии - Трактаты . 245 (1): 117–125. дои : 10.1127/0077-7749/2007/0245-0117 .
- ^ Нибур, Б.; Пюрнер, Т. (2014). «Платтенкалк и франконские доломиты - литостратиграфия белой юрской группы франконского альба (внеальпийская верхняя юра, Бавария)» . Серия публикаций Немецкого общества наук о Земле . 83 :5-72. дои : 10.1127/sdgg/83/2014/5 .
- ^ Раухут, OWM; Тишлингер, Х.; Фот, К. (2019). «Неархеоптеригидный птичий теропод из поздней юры южной Германии» . электронная жизнь . 8 : е43789. doi : 10.7554/eLife.43789 . ПМК 6516837 . ПМИД 31084702 .
- ^ Мённиг, Э.; Франц, М.; Швайгерт, Г. (2018). «Юрский период в стратиграфической таблице Германии (STD 2016)» [Стратиграфическая карта Германии (STD 2016): Юрский период]. Журнал Немецкого общества наук о Земле . 169 (2): 225–246. дои : 10.1127/zdgg/2018/0148 . S2CID 135104883 .
- ^ Перейти обратно: а б Муннеке, А.; Вестфаль, Х.; Кёльбл-Эберт, М. (2008). «Диагенез платтенкалка: примеры из района Зольнхофена (верхняя юра, южная Германия)». Седиментология . 55 (6): 1931–1946. Бибкод : 2008Седим..55.1931М . дои : 10.1111/j.1365-3091.2008.00975.x . S2CID 129127346 .
- ^ Перейти обратно: а б Уилкин, Дж. (2020). «Южногерманский Платтенкалькс». Геология сегодня . 36 (1): 27–32. дои : 10.1111/gto.12288 . S2CID 213606153 .
- ^ Виол, Г. (1994). «Рыбная тафономия Зольнхофенского платтенкалка — подход к реконструкции палеосреды». Геобиос . 27 : 81–90. дои : 10.1016/s0016-6995(94)80023-5 .
- ^ Сейлахер, А.; Рейф, МЫ; Вестфаль, Ф. (1985). «Седиментологические, экологические и временные закономерности ископаемых Лагерштеттенов». Философские труды Лондонского королевского общества B: Биологические науки . 311 (1148): 5–24. Бибкод : 1985РСТБ.311....5С . дои : 10.1098/rstb.1985.0134 .
- ^ Бердмор, СР; Лоулор, Э.; Хон, DWE (2017). «Использование тафономии для вывода о различиях в мягких тканях между таксонами: пример использования базальных и производных форм птерозавров Зольнхофена». Наука о природе . 104 (7–8): 65. Бибкод : 2017SciNa.104...65B . дои : 10.1007/s00114-017-1486-0 . ПМИД 28721556 . S2CID 33984022 .
- ^ Сьюс, Х.-Д. (2009). « Археоптерикс : Der Urvogel von Solnhofen, Питер Велнхофер». Журнал палеонтологии позвоночных . 29 (1): 310–311. дои : 10.1080/02724634.2009.10010387 . S2CID 85327790 .
- ^ Беннетт, Южная Каролина (1995). «Статистическое исследование Rhamphorhynchus из известняка Зольнхофен в Германии: годовые классы одного крупного вида». Журнал палеонтологии . 69 (3): 569–580. дои : 10.1017/S0022336000034946 .
- ^ Теннант, JP; Мэннион, Вашингтон; Апчерч, П. (2016). «Эволюционные связи и систематика Atoposauridae (Crocodylomorpha: Neosuria): значение для возникновения Eusuria» . Зоологический журнал Линнеевского общества . 177 (4): 854–936. дои : 10.1111/zoj.12400 . hdl : 10044/1/29026 .
- ^ Эберт, М. (2012). «Рачки как добыча рыб Зольнхофенского архипелага» . Археоптерикс . 30 : 1–4.
- ^ Арратия, Г. (2016). «Новые замечательные костистые кости позднеюрского периода из южной Германии: Ascalaboidae n. fam., их содержание, морфология и филогенетические взаимоотношения» . Ископаемый рекорд . 19 (1): 31–59. дои : 10.5194/фр-19-31-2016 .
- ^ Эберт, М.; Кёльбл-Эберт, М. (2010). «Морфология передних краев хвостового и других непарных плавников позднеюрских Aspidorhynchidae и некоторых других актиноптеригийных рыб в свете их функции» . Археоптерикс . 28 : 1–22.
- ^ Эберт, М. (2014). «Род Belonostomus Agassiz, 1834 (Neopterygii, Aspidorhynchiformes) в поздней юре архипелага Зольнхофен, с акцентом на Belonostomus kochii Münster, 1836 из Эттлинга (Германия)» . Археоптерикс . 32 : 15–43.
- ^ Ламберс, PH (1994). «Галекоморфные рыбы Caturus и Amblysemius в литографическом известняке Зольнхофена (Титон), Бавария». Геобиос . 27 : 91–99. дои : 10.1016/S0016-6995(94)80024-3 .
- ^ Кривет, Дж.; Шмитц, Л. (2005). «Новый взгляд на распространение и палеобиологию пикнодонтовой рыбы Gyrodus » . Acta Palaeontologica Polonica . 50 (1).
- ^ Эберт, М.; Лейн, Дж.А.; Кёльбл-Эберт, М. (2016). « Palaeomacrosemius thiollieri , gen. et sp. nov., новый Macrosemiidae (Neopterygii) из верхней юры архипелага Зольнхофен (Германия) и Церина (Франция), с ревизией рода Macrosemius ». Журнал палеонтологии позвоночных . 36 (5): e1196081. дои : 10.1080/02724634.2016.1196081 . S2CID 88649682 .
- ^ Эберт, М. (2012). « Histionotus со множеством вопросительных знаков» ) – род ( Actinopterygii , Macrosemiidae Археоптерикс . 30 :5-15.
- ^ Эберт, М. (2019). « Zandtfuro и Schernfeldfuro , новые роды Halecomorphi (Actinopterygii) из верхнеюрского архипелага Зольнхофен». Журнал палеонтологии позвоночных . 39 (2): e1592759. дои : 10.1080/02724634.2019.1592759 . S2CID 181452064 .
- ^ Тис, Д.; Лейднер, А. (2011). «Акулы и рыбы-гитаристы (Elasmobranchii) из поздней юры Европы» (PDF) . Палеоразнообразие . 4 : 63–184.
- ^ Фукс, Д.; Хейнг, AM; Кеупп, Х. (2013). « Acanthoeuthispromatica Naef, 1922, почти забытый таксон и его роль в интерпретации скелетов рук головоногих моллюсков». Новогодний ежегодник геологии и палеонтологии - Трактаты . 269 (3): 241–250. дои : 10.1127/0077-7749/2013/0347 .
- ^ Энгезер, Т.; Рейтнер, Дж. (1992). [Новый образец Belemnoteuthis mayri Solnhofener Plattenkalk (Untertithonium) von Wintershof, Bayern» « Ein neues Exemplar von Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) aus dem Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) из Зольнхофена известняк (нижний титон) из Винтерсхофа, Бавария] (PDF) . Археоптерикс . 10 :13–17.
- ^ Клюг, К.; Фукс, Д.; Швайгерт, Г.; Репер, М.; Тишлингер, Х. (2015). «Новая анатомическая информация о руках и плавниках исключительно сохранившегося Plesioteuthis (Coleoidea) из поздней юры Германии» (PDF) . Швейцарский журнал палеонтологии . 134 (2): 245–255. дои : 10.1007/s13358-015-0093-y . S2CID 83370427 .
- ^ Хесс, Х.; Эттер, В. (2011). «Жизнь и смерть Саккокомы тенеллы (Гольдфусса)» . Швейцарский журнал геонаук . 104 (1): 99–106. дои : 10.1007/s00015-011-0059-z . S2CID 85358369 .
- ^ Винклер, Н. (2017). «Две новые креветки-пенеиды (Crustacea: Decapoda: Dendrobranchiata) из литографических известняков Зольнхофена (верхняя юра, южная Германия)». Новогодний ежегодник геологии и палеонтологии - Трактаты . 283 (1): 9–24. дои : 10.1127/njgpa/2017/0623 .
- ^ Ломакс, ДР; Ракай, Калифорния (2012). «Длинный мортихниальный путь Mesolimulus walchi из верхнеюрского литографического известняка Зольнхофен недалеко от Винтерсхофа, Германия» . Ихнос . 19 (3): 175–183. дои : 10.1080/10420940.2012.702704 . S2CID 55610538 .
- ^ Гёлих, UB (2017). «Каталог запасов ископаемых птиц Баварской государственной коллекции палеонтологии и геологии в Мюнхене» (PDF) . Зиттелиана . 89 : 331–349.
- ^ Видович, СУ; Мартилль, DM (2017). «Таксономия и филогения Diopecephalus kochi (Wagner, 1837) и Germanodactylus rhamphastinus (Wagner, 1851)» (PDF) . Геологическое общество, Лондон, специальные публикации . 455 (1): 125–147. дои : 10.1144/sp455.12 . S2CID 219204038 .
- ^ Перейти обратно: а б с Караннанте, Г.; Синьор, М.; Вигорито, М. (2006). «Богатый позвоночными Платтенкалк Пьетрарои (нижний мел, Южные Апеннины, Италия): новая модель». Фации . 52 (4): 555–577. дои : 10.1007/s10347-006-0075-z . S2CID 129309028 .
- ^ Перейти обратно: а б с Брави, С.; Гарассино, А (1998). «Новые биостратографические и палеоэкологические наблюдения над «Платтенкалком» нижнего мела (альб) Пьетрарои (Беневенто, Южная Италия) и его комплексом десятиногих ракообразных» . Atti Societá Italiana Scienze Naturali, Милан . 138 : 119–171.
- ^ Эванс, SE; Райя, П.; Барбера, К. (2006). «Нижнемеловая ящерица рода Chometokadmon из Италии». Меловые исследования . 27 (5): 673–683. дои : 10.1016/j.cretres.2006.03.004 .
- ^ Кау, А.; Баяно, Массачусетс; Райя, П. (2014). «Новый сфенодонт (Reptilia, Lepidosauria) из нижнего мела Южной Италии и филогенетическое сходство ринхоцефалов Pietraroia Plattenkalk». Меловые исследования . 49 : 172–180. дои : 10.1016/j.cretres.2014.02.001 .
- ^ Даль Сассо, К.; Синьор, М. (1998). «Исключительная сохранность мягких тканей динозавра-теропода из Италии». Природа . 392 (6674): 383–387. Бибкод : 1998Natur.392..383D . дои : 10.1038/32884 . S2CID 4325093 .
- ^ Бускалиони, AD; Пирас, П.; Вулло, Р.; Синьор, М.; Барбера, К. (2011). «Ранняя eusuchia crocodylomorpha из богатого позвоночными Plattenkalk из Пьетрарои (нижний альб, южные Апеннины, Италия)» . Зоологический журнал Линнеевского общества . 163 (С1): С199–С227. дои : 10.1111/j.1096-3642.2011.00718.x .
- ^ Макгоуэн, Дж.; Эванс, SE (1995). «Альбанерпетонтидные амфибии из мела Испании». Природа . 373 (6510): 143–145. Бибкод : 1995Natur.373..143M . дои : 10.1038/373143a0 . S2CID 4279738 .
- ^ Таверн, Л.; Капассо, Л.; Дель Ре, М. (2020). «Остеология и филогенетические взаимоотношения Gregoriopycnodus bassanii gen. nov., рыбы-пикнодонта (Pycnodontidae) из морского альба (нижний мел) Пьетрарохи (южная Италия)» (PDF) . Гео Эко Троп . 44 (1): 161–174.
- ^ Перейти обратно: а б Далла Веккья, Ф.; Барбера, К.; Биццарини, Ф.; Молодец, С.; Дельфино, М.; Джусберти, Л.; Гуидотти, Дж.; Миетто, П.; Палаццони, К.; Роги, Г.; Лорд, М.; Симона, О. (2005). «Морской мел» . В Бонфильо, Л. (ред.). Палеонтология позвоночных в Италии: биологическая эволюция, экологическое и палеогеографическое значение [ Палеонтология позвоночных в Италии: биологическая эволюция, экологическое и палеогеографическое значение ]. Секция наук о Земле. Том 6. Мемуары Гражданского музея естественной истории Вероны. стр. 101–112.
- ^ Таверн, Л.; Капассо, Л. (2019). «Остеология Cavinichthys pachylepis gen. и sp. nov. (Teleostei, Crossognathiformes, Pachyrhizodontidae) из Пьетрарои (нижний мел, Кампания, южная Италия)». Талассия Салентина . 41 : 53–70. дои : 10.1285/i15910725v41p53 .
- ^ Таверн, Л.; Капассо, Л. (2017). «Остеология и взаимоотношения Caeus («Chanos») leopoldi (Teleostei, Gonorynchiformes, Chanidae) из морского альба (ранний мел) Пьетрарохи (Кампания, южная Италия)». Бюллетень Гражданского музея естественной истории Вероны, геология, палеонтология, предыстория . 41 :3–20. S2CID 52025491 .
- ^ Таверн, LP; Капассо, Л. (2016). «Ревизия Ionoscopus petrarojae (Ionoscopiformes, Osteichthyes) из альба Пьетрарохи (Кампания, южная Италия)». Талассия Салентина . 38 : 65–80. дои : 10.1285/i15910725v38p65 .
- ^ Таверн, Л.; Капассо, Л. (2017). « Italophiopsis derasmoi gen. and sp. nov. (Ionoscopiformes, Italophiopsidae fam. nov.) из мела Пьетрарохи (Италия)». Талассия Салентина . 39 :9–24. дои : 10.1285/i15910725v39p9 .