Магнапорте гризеа
| Магнапорте гризеа | |
|---|---|
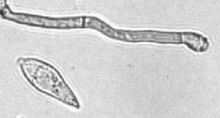 | |
| Конидий и конидиогенная клетка | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Грибы |
| Разделение: | Аскомикота |
| Сорт: | Сордариомицеты |
| Заказ: | Магнапорталес |
| Семья: | Магнапортовые |
| Род: | Магнапорте |
| Разновидность: | М. Гризеа |
| Биномиальное имя | |
| Магнапорте гризеа (Т.Т. Хеберт) М.Э. Барр | |
| Синонимы | |
Ceratosphaeria grisea TT Hebert, (1971) | |
Magnaporthe grisea , также известный как грибок рисового ожога , гнилая шейка риса , болезнь рисовых рассады , болезнь риса , овальная пятнистость листьев злаков , точечная болезнь , болезнь райграса , пятнистость Джонсона , [1] [2] [3] [4] [5] [6] [7] взрыв шеи , [8] [9] [10] [11] пшеничный взрыв [12] и Имоти ( 稲熱 ) — растительнопатогенный гриб и модельный организм. [13] это вызывает серьезное заболевание, поражающее рис . Сейчас известно, что M. grisea представляет собой загадочный комплекс видов , содержащий по крайней мере два биологических вида, которые имеют явные генетические различия и не скрещиваются между собой. [14] Члены комплекса, выделенные из Digitaria, были более узко определены как M. grisea . Остальные члены комплекса, выделенные из риса и ряда других хозяев, были переименованы в Magnaporthe oryzae в рамках того же комплекса M. grisea . [14] Сохраняется путаница в отношении того, какое из этих двух названий использовать для обозначения возбудителя рисового взрыва, поскольку оба сейчас используются разными авторами.
Члены комплекса M. grisea могут также инфицировать другие важные для сельского хозяйства злаки , включая пшеницу , рожь , ячмень и просо, вызывая заболевания, называемые взрывной болезнью или фитофторозом . Рисовый взрыв ежегодно приводит к экономически значимым потерям урожая. По оценкам, ежегодно уничтожается достаточно риса, чтобы накормить более 60 миллионов человек. Известно, что гриб встречается в 85 странах мира. [15] и по состоянию на 2003 год [update] был самым разрушительным грибковым патогеном растений в мире. [13]
Хозяева и симптомы
[ редактировать ]
M. grisea — аскомицетный гриб. Это чрезвычайно эффективный патоген растений, поскольку он может размножаться как половым, так и бесполым путем с образованием специализированных инфекционных структур, аппрессорий , которые заражают воздушные ткани, и гифов, которые могут заражать ткани корня .
Рисовый взрыв наблюдался на штаммах риса М-201, М-202, М-204, М-205, М-103, М-104, S-102, L-204, Calmochi-101, причем М-201 является самый уязвимый. [16] Первоначальные симптомы представляют собой поражения от белого до серо-зеленого цвета или пятна с более темными краями, возникающие на всех частях побега, тогда как более старые поражения имеют эллиптическую или веретенообразную форму и от белесого до серого цвета с некротическими краями. Поражения могут увеличиваться и сливаться, приводя к гибели всего листа. Симптомы наблюдаются на всех надземных частях растения. [17] Поражения можно увидеть на воротнике листа , стебле , узлах соломины и метелки узле шейки . Межузловое заражение стебля протекает по полосчатой схеме. Узловая инфекция приводит к разрыву стебля в зараженном узле (гнилая шейка). [18] Это также влияет на размножение, заставляя хозяина производить меньше семян. Это вызвано заболеванием, препятствующим созреванию настоящего зерна. [15]
Цикл заболевания
[ редактировать ]
Возбудитель заражается в виде споры, которая вызывает повреждения или пятна на частях растения риса, таких как лист, листовая шейка, метелка, стебель и узлы соломины. С помощью структуры, называемой апрессорием , возбудитель проникает в растение. аппрессориума Клеточная стенка хитиновая , а ее внутренняя сторона содержит меланин . [1] : 184 что необходимо для повреждения структур хозяина. [1] : 184 [13] Тургорное давление, создаваемое во время этого процесса, достаточно для обычного проникновения в кутикулу растений и экспериментально может проникать через кевлар . Этот впечатляющий тургор создается за счет синтеза глицерина и поддерживается вышеупомянутым аппрессорным меланином. [13] Возбудитель способен перемещаться между растительными клетками, используя свои инвазивные гифы и проникая через плазмодесмы . [19] Затем M. grisea образует споры из пораженной ткани риса и рассеивается в виде конидиоспор . [20] После зимовки в таких источниках, как рисовая солома и стерня, цикл повторяется. [15]
Один цикл может быть завершен примерно за неделю при благоприятных условиях, когда одно поражение может привести к образованию до тысяч спор за одну ночь. Однако очаги заболевания могут появиться через три-четыре дня после заражения. [21] Благодаря способности продолжать производить споры более 20 дней, поражения риса, вызванные взрывом, могут иметь разрушительные последствия для восприимчивых культур риса. [22]
Заражение риса вызывает фосфорилирование белка светособирающего комплекса II. LHCB5 . [23] LHCB5 необходим для выброса активных форм кислорода, вырабатываемого хозяином, что обеспечивает устойчивость к этому патогену . [23]
Среда
[ редактировать ]Рисовый взрыв является серьезной проблемой в регионах с умеренным климатом и может быть обнаружен в таких районах, как орошаемые низменности и возвышенности. [24] Условия, благоприятные для заражения риса, включают длительные периоды свободной влаги и/или высокой влажности, поскольку для заражения необходима влажность листьев. [24] Спорообразование увеличивается при высокой относительной влажности, а при 25–28 ° C (77–82 ° F) прорастание спор, образование повреждений и спорообразование находятся на оптимальном уровне. [15]
С точки зрения борьбы, чрезмерное использование азотных удобрений , а также стресс от засухи повышают восприимчивость риса к возбудителю, поскольку растение находится в ослабленном состоянии и его защитные силы низки. [15] Затопление и осушение полей являются нормальным явлением при выращивании риса, однако оставление поля осушенным в течение длительного периода времени также способствует заражению, поскольку это приведет к аэрации почвы, преобразованию аммония в нитрат и, таким образом, также вызывая стресс у посевов риса. [15]
Географическое распространение
[ редактировать ]Взрыв пшеницы был обнаружен в сезон дождей 2017-2018 годов в Замбии , в районе Мпика провинции Мучинга. [25] [26]
В феврале 2016 года разрушительная эпидемия пшеницы поразила Бангладеш . [27] [28] Транскриптомный анализ показал, что это линия M. grisea , скорее всего, из Минас-Жерайс , Сан-Паулу , Бразилиа и Гояс штатов в Бразилии , а не из каких-либо географически близких штаммов. [27] [28] Этот успешный диагноз демонстрирует способность генетического надзора раскрыть новые последствия трансконтинентальных перевозок для биобезопасности. [27] [28] и позволяет быстро применить бразильский опыт к ситуации в Бангладеш. [27] [28] С этой целью правительство создало систему раннего предупреждения для отслеживания его распространения по стране. [28]
Управление
[ редактировать ]
Этот гриб сталкивается как с фунгицидами , так и с генетической устойчивостью в некоторых сортах риса, выведенных селекционерами . Он способен создать как устойчивость к этим химическим обработкам , так и вирулентность к устойчивости сельскохозяйственных культур посредством генетических изменений посредством мутации . Для наиболее эффективного контроля инфекции, вызванной M. grisea , необходимо реализовать комплексную программу управления, чтобы избежать чрезмерного использования одного метода контроля и борьбы с генетической устойчивостью. Например, удаление растительных остатков может снизить вероятность перезимовки и препятствовать инокуляции в последующие сезоны. Другая стратегия заключалась бы в высадке устойчивых сортов риса, которые не так восприимчивы к заражению M. grisea . [15] Знания о патогенности M. grisea и ее потребности в свободной влаге позволяют предположить другие стратегии борьбы, такие как регулируемое орошение и сочетание химических обработок с различными способами действия. [15] Управление количеством воды, подаваемой посевам, ограничивает подвижность спор, тем самым уменьшая возможность заражения. Было показано, что химические средства контроля, такие как карпропамид, предотвращают проникновение аппрессорий в эпидермальные клетки риса, оставляя зерно нетронутым. [29] Пападжани и др. В 2015 году было установлено, что эфирные масла душицы обыкновенной и розмарина лекарственного эффективны in vitro , и указаны пороговые значения лечения. [30] : 107–108
Штамм бластной пшеницы можно диагностировать путем секвенирования. [12] : 45 Тьерри и др. , 2020 представляет набор генетических маркеров , которые можно обнаружить с помощью полимеразной цепной реакции (ПЦР), ПЦР в реальном времени (ОТ-ПЦР) и петлевой изотермической амплификации (LAMP). [12] : 45 Большим преимуществом маркеров Тьерри является то, что они не пропускают , например, изоляты, не имеющие последовательности Mot3. BR0032 и его высокая чувствительность . [12] : 45
Некоторые инновационные биологически-имитирующие фунгициды разрабатываются на основе малых РНК и коротких пептидов . [31] SNP-D4 представляет собой короткий пептид, обнаруженный при скрининге библиотеки in vitro на M. oryzae кальмодулин . [31] Он связывается с кальмодулином, ингибирует образование конидий и блокирует прорастание спор . [31]
Важность
[ редактировать ]Рисовая болезнь является наиболее опасным заболеванием рисовых культур в мире. Поскольку рис является важным источником пищи для большей части мира, его воздействие имеет широкий диапазон. Он был обнаружен более чем в 85 странах мира и достиг Соединенных Штатов в 1996 году. Ежегодно количество урожая, погибающего в результате рисового взрыва, могло прокормить 60 миллионов человек. Хотя существуют некоторые устойчивые штаммы риса, болезнь сохраняется везде, где выращивается рис. Болезнь никогда не была искоренена в регионе. [32]
Штаммы
[ редактировать ]Три штамма : альбинос (определяемый мутацией в локусе ALB1 ), желтоватый ( BUF1 ) и розовый ( RSY1 ) были тщательно изучены, поскольку они непатогенны. Было обнаружено, что это связано с их неспособностью синтезировать меланин , который является фактором вирулентности у некоторых грибов. [1] : 184 pathovar triticum Штамм ( M. o. pv. triticum ) вызывает пиролиз пшеницы . [12]
Генетика
[ редактировать ]Полногеномные последовательности стали возможны и стали доступны только в 2003 году. [13]
Митоген -активируемая протеинкиназа (MAPK) под названием pmk1 генетически близка к той, которая необходима для спаривания и морфологии клеток у дрожжей , FUS3 / KSS1 . Дефектные мутантные дрожжи частично или полностью восстанавливают функцию спаривания, если им дать копию pmk1 . Поэтому предполагалось, что это должен быть только ген спаривания и развития у M. grisea , однако оказалось, что он жизненно важен как для процесса спаривания самок, так и для функции аппрессориума и патогенности в целом. [13]
и ряд других, было показано, что сигнальные связи между MAPK и циклическими аденозинмонофосфатами необходимы для спаривания Поскольку в нескольких других моделях, включая Ustilago maydis , предполагалось, что это верно для M. grisea , однако затем было показано, что в этой модели это не является необходимым. модель. Это демонстрирует значительное разнообразие клеточных функций грибов. [13]
аланин трансаминаза Было показано, что : глиоксилатаминотрансфераза 1 (AGT1) имеет решающее значение для патогенности M. grisea благодаря поддержанию окислительно-восстановительного гомеостаза в пероксисомах. Липиды, транспортируемые в аппрессории во время проникновения в хозяина, разлагаются в большой центральной вакуоли, в результате чего образуются жирные кислоты . β-Окисление жирных кислот представляет собой процесс производства энергии, в ходе которого образуются ацетил-КоА и восстановленные молекулы ФАДН 2 и НАДН , которые должны быть окислены для поддержания окислительно-восстановительного гомеостаза в аппрессориях. AGT1 способствует ферментации лактата, окисляя НАДН/ФАДН 2 . при этом [33]
Было обнаружено, что мутанты M. grisea , лишенные гена AGT1, являются непатогенными из-за их неспособности проникать через поверхностные мембраны хозяина. Это указывает на возможность нарушения утилизации липидов у M. grisea appressoria в отсутствие гена AGT1. [34]
Биохимия взаимодействия хозяин-возбудитель
[ редактировать ]В обзоре 2010 года сообщалось, что клоны обладают количественной устойчивостью растений к болезням. [35] Растение риса реагирует на возбудитель взрыва, высвобождая жасмоновую кислоту , которая активирует дальнейшие метаболические пути , вызывающие защитную реакцию. [36] Он накапливается в виде метилжасмоновой кислоты . [36] Возбудитель реагирует синтезом , окислительного фермента который предотвращает это накопление и вызывающий его сигнал тревоги. [36] Os Pii-2 — рисовый белок, действующий как иммунорецептор . [37] Он связывается с собственным рисом Exo70-F3 Белок . [37] Этот белок является мишенью M. oryzae. эффектора АВР-Пии , выделяемые грибом при заражении. Таким образом, это позволяет белку Os Pii-2 отслеживать M. oryzae на эту цель. атаку [37] Некоторые сорта риса несут аллели устойчивости гена OsSWEET13 , который производит молекулярную мишень X. oryzae pv. oryzae эффектор PthXo2 . [38]
См. также
[ редактировать ]- Серая пятнистость листьев кукурузы , аналогичное заболевание кукурузы/кукурузы.
- Серая пятнистость листьев , аналогичное заболевание и у других трав.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Талбот, Николас Дж. (2003). «По следам зернового убийцы: изучение биологии Magnaporthe grisea ». Ежегодный обзор микробиологии . 57 (1). Годовые обзоры : 177–202. дои : 10.1146/annurev.micro.57.030502.090957 . ISSN 0066-4227 . ПМИД 14527276 .
Три мутанта M. grisea , albino , buff и rosy (соответствующие локусам ALB1 , BUF1 и RSY1 соответственно) широко изучены и являются непатогенными. Это происходит из-за невозможности пересечь кутикулу растения из-за отсутствия отложения меланина в аппрессории.
- ^ Зейглер, RS; Леонг, ЮАР; Тинг, PS (1994). «Рисовая болезнь». Уоллингфорд, Великобритания CABI : Международный центр сельского хозяйства и биологических наук .
- ^ Уилсон, РА; Талбот, Нью-Джерси (2009). «Под давлением: исследование биологии заражения растений Magnaporthe oryzae ». Обзоры природы Микробиология . 7 (3): 185–95. дои : 10.1038/nrmicro2032 . ПМИД 19219052 . S2CID 42684382 .
- ^ Сесма, А.; Осборн, А.Э. (2004). «Возбудитель взрыва листьев риса претерпевает процессы развития, типичные для грибов, поражающих корни». Природа . 431 (7008): 582–6. Бибкод : 2004Natur.431..582S . дои : 10.1038/nature02880 . ПМИД 15457264 . S2CID 549194 .
- ^ Дин, РА; Талбот, Нью-Джерси ; Эббол, диджей; Фарман, ML; Митчелл, ТК; Орбах, MJ; Тон, М; Кулкарни, Р; Сюй, младший; Пан, Х; Прочтите, Н.Д.; Ли, Ю.Х.; Карбоне, я; Браун, Д; О, да; Донофрио, Н.; Чон, Дж.С.; Соунс, DM; Джонович, С; Коломиец Е; Ремейер, К; Ли, В; Хардинг, М; Ким, С; Лебрен, Миннесота; Бонерт, Х; Кофлан, С; Батлер, Дж; Кальво, С; и др. (2005). «Последовательность генома рисового гриба Magnaporthe grisea » . Природа . 434 (7036): 980–6. Бибкод : 2005Natur.434..980D . дои : 10.1038/nature03449 . ПМИД 15846337 .
- ^ Коуч, Британская Колумбия; Кон, LM (2002). «Генеалогия мультилокусных генов, согласующаяся с предпочтениями хозяина, указывает на отделение нового вида, Magnaporthe oryzae , от M. grisea ». Микология . 94 (4): 683–93. дои : 10.2307/3761719 . JSTOR 3761719 . ПМИД 21156541 .
- ^ Magnaporthe grisea. Архивировано 12 октября 2007 г. в Wayback Machine в Сборнике по защите растений. Архивировано 16 июля 2007 г. в Wayback Machine , CAB International.
- ^ Те Бест (2007). «Рисовый взрыв». Инструктор по здоровью растений . Американское фитопатологическое общество (APS). дои : 10.1094/phi-i-2007-0313-07 . ISSN 1935-9411 .
- ^ Хан, Мохаммад Ашик Икбал; Реджван Бхуян, Мохаммед; Хоссейн, Мохаммад Шахадат; Пратим Сен, Парта; Ара, Анджуман; Абубакар Сиддик, доктор медицины; Ансар Али, Мэриленд (2014). «Шейковая инфекция влияет на урожайность зерна и качественные характеристики ароматного риса» . Comptes Rendus Biologies . 337 (11). Эльзевир Массон : 635–641. дои : 10.1016/j.crvi.2014.08.007 . ISSN 1631-0691 . ПМИД 25444707 .
- ^ Румен, ЕС (1992). «Частичная устойчивость к шейковому взрыву зависит от стадии развития метелки и генотипа риса». Эвфитика . 64 (3). Springer Science and Business Media LLC : 173–182. дои : 10.1007/bf00046046 . ISSN 0014-2336 . S2CID 45126761 .
- ^ Титоне, Патриция; Монджано, Габриэле; Тамборини, Луиджи (4 января 2015 г.). к шейному взрыву, вызванному Pyriclelia oryzae «Устойчивость итальянских сортов риса » . Европейский журнал патологии растений . 142 (1). Springer Science and Business Media LLC : 49–59. Бибкод : 2015EJPP..142...49T . дои : 10.1007/s10658-014-0588-1 . ISSN 0929-1873 . S2CID 14478689 .
- ^ Jump up to: а б с д и Кумар, Судхир; Кашьяп, Прем; Сингх, Гьянендра (2020). Пшеничный взрыв (1-е изд.). Бока-Ратон, Флорида , США: CRC Press . дои : 10.1201/9780429470554 . ISBN 978-0-429-47055-4 . OCLC 1150902336 . S2CID 235049332 .
- ^ Jump up to: а б с д и ж г Ярден, О.; Эббол, диджей; Фриман, С.; Родригес, Р.Дж.; Дикман, МБ (2003). «Грибная биология и сельское хозяйство: новый взгляд на эту область» . Молекулярные растительно-микробные взаимодействия . 16 (10). Американское фитопатологическое общество (APS): 859–866. дои : 10.1094/mpmi.2003.16.10.859 . ISSN 0894-0282 . ПМИД 14558687 . S2CID 20430256 .
- ^ Jump up to: а б Коуч, Британская Колумбия; Фудал, я; Лебрен, Миннесота; Тарро, Д; Валент, Б; Ван Ким, П; Ноттегем, JL; Кон, LM (2005). «Происхождение специфичных для хозяина популяций возбудителя взрыва Magnaporthe oryzae при одомашнивании сельскохозяйственных культур с последующим распространением пандемических клонов на рисе и сорняках риса» . Генетика . 170 (2): 613–30. doi : 10.1534/genetics.105.041780 . ПМЦ 1450392 . ПМИД 15802503 .
- ^ Jump up to: а б с д и ж г час Скардачи, Южная Каролина; и др. (2003). «Рисовый взрыв: новая болезнь в Калифорнии» . Серия информационных бюллетеней по агрономии . Калифорнийский университет в Дэвисе (UCD). Архивировано из оригинала 11 сентября 2006 г. Проверено 25 февраля 2014 г.
- ^ Rice Blast в Комплексной борьбе с вредителями Калифорнийского университета (UC-IPM)
- ^ Rice Blast в онлайн-информационной службе по нехимической борьбе с вредителями в тропиках.
- ^ Rice Blast. Архивировано 20 октября 2010 г. в Wayback Machine в информационных бюллетенях по химическим и биологическим боевым агентам.
- ^ Сакулку, Васин; Осес-Руис, Мириам; Оливейра Гарсия, Эли; Соунс, Даррен; Литтлджон, Джордж; Хакер, Кристиан; Коррейя, Ана; Валент, Барбара; Талбот, Николас (23 марта 2018 г.). «Одна грибковая MAP-киназа контролирует межклеточную инвазию рисового гриба» . Наука . 359 (6382): 1399–1403. Бибкод : 2018Sci...359.1399S . doi : 10.1126/science.aaq0892 . hdl : 10871/32530 . ПМИД 29567712 .
- ^ Агриос, Джордж Н. (2005). Патология растений . Амстердам : Elsevier Academic Press .
- ^ Уилсон, Ричард; Талбот, Николас (1 марта 2009 г.). «Под давлением: исследование биологии заражения растений Magnaporthe oryzae ». Обзоры природы Микробиология . 7 (3): 185–189. дои : 10.1038/nrmicro2032 . ПМИД 19219052 . S2CID 42684382 .
- ^ Методы диагностики рисового взрыва [ постоянная мертвая ссылка ] в наборе инструментов для биобезопасности растений PaDIL
- ^ Jump up to: а б Лю, Синьюй; Чжан, Чжэнгуан (2022). «Меч о двух концах: активные формы кислорода (АФК) во время взаимодействия рисового грибка и хозяина». Журнал ФЭБС . 289 (18). John Wiley & Sons, Inc. ( Федерация европейских биохимических обществ : 5505–5515. doi : /febs.16171 . PMID 34453409. . S2CID 237340135 10.1111
- ^ Jump up to: а б Куек, Девлин (2000). «Последствия корпоративных стратегий для исследований риса в Азии» . Зерно . Проверено 20 октября 2010 г.
- ^ «Исследователи в Замбии подтверждают: пшеничный взрыв совершил межконтинентальный прыжок в Африку» . 24 сентября 2020 года. Архивировано из оригинала 6 октября 2021 года . Проверено 1 октября 2020 г.
- ^ Тембо, Батисеба; Муленга, Рабсон М.; Сичилима, Сувиланджи; м'Сиска, Кеннет К.; Мвале, Моисей; Чикоти, Патрик С.; Сингх, Паван К.; Он, Синьяо; Педли, Керри Ф.; Петерсон, Гэри Л.; Сингх, Рави П.; Браун, Ханс Дж. (2020). «Обнаружение и характеристика грибка ( Magnaporthe oryzae патотип Triticum ), вызывающего пиролиз на неорошаемой пшенице ( Triticum aestivum L.) в Замбии» . ПЛОС ОДИН . 15 (9). Публичная научная библиотека : e0238724. Бибкод : 2020PLoSO..1538724T . дои : 10.1371/journal.pone.0238724 . ПМЦ 7505438 . ПМИД 32956369 . S2CID 221843315 .
- ^ Jump up to: а б с д Ислам, М. Тофаззал; Кролл, Дэниел; Гладье, Пьер; Соунс, Даррен М.; Персоны, Антуан; Бхаттачарджи, Паллаб; Хоссейн, штат Мэриленд Шаид; Гупта, Дипали Рани; Рахман, штат Мэриленд Махбубур; Махбуб, М. Голам; Кук, Никола; Салам, Мойн У.; Суровый, Мусрат Захан; Санчо, Ванесса Буэно; Масиэль, Жоау Леодато Нуньес; НханиЖуниор, Антонио; Кастроагудин, Ванина Лилиан; Реджес, Джулиана Т. де Ассис; Черезини, Пауло Сезар; Равель, Себастьян; Келлнер, Ронни; Фурнье, Элизабет; Тарро, Дидье; Лебрен, Марк-Анри; Макдональд, Брюс А.; Ститт, Тимоти; Лебедь, Дэниел; Талбот, Николас Дж.; Сондерс, Дайан ГО; Выиграй, Джо; Камун, Софиен (3 октября 2016 г.). «Появление болезни пшеницы в Бангладеш было вызвано южноамериканской линией Magnaporthe oryzae » . БМК Биология . 14 (1). Springer Science and Business Media LLC : 84. doi : 10.1186/s12915-016-0309-7 . ISSN 1741-7007 . ПМК 5047043 . ПМИД 27716181 .
- ^ Jump up to: а б с д и «Новая инфографика демонстрирует систему раннего предупреждения о взрыве пшеницы в Бангладеш» . CGIAR ПШЕНИЦА . 15 июля 2020 г. Архивировано из оригинала 01 декабря 2020 г. Проверено 26 декабря 2020 г.
- ^ Курахаси, Ёсио (1997). «Биологическая активность карпропамида (КТУ 3616): новый фунгицид против фунгицида риса» . Журнал пестицидной науки . Проверено 25 февраля 2014 г.
- ^ Фурментен, Софи; Крини, Грегорио; Лихтфаус, Эрик, ред. (2018). Экологическая химия для устойчивого мира . Том. 17. Чам, Швейцария : Springer International Publishing . дои : 10.1007/978-3-319-76162-6 . ISBN 978-3-319-76161-9 . ISSN 2213-7114 . S2CID 199492358 .
- ^ Jump up to: а б с Роза, Стефано; Пезарези, Паоло; Миццотти, Кьяра; Булоне, Винсент; Меццетти, Бруно; Баральди, Елена; Масьеро, Симона (2021). «Изменяющие правила игры альтернативы обычным фунгицидам: малые РНК и короткие пептиды». Тенденции в биотехнологии . 40 (3). Cell Press : 1–18. дои : 10.1016/j.tibtech.2021.07.003 . ISSN 0167-7799 . ПМИД 34489105 . S2CID 237433001 .
- ^ Rice Blast. Архивировано 31 июля 2010 г. в Wayback Machine в Cereal Knowledge Bank.
- ^ Бхадаурия, Виджай; Банниза, Сабина; Ванденберг, Альберт; Сельварадж, Гопалан; Вэй, Яндоу (27 апреля 2012 г.). «Пероксисомальный аланин: глиоксилатаминотрансфераза AGT1 незаменима для функции аппрессориума возбудителя рисового взрыва, Magnaporthe oryzae » . ПЛОС ОДИН . 7 (4): e36266. Бибкод : 2012PLoSO...736266B . дои : 10.1371/journal.pone.0036266 . ISSN 1932-6203 . ПМЦ 3338719 . ПМИД 22558413 .
- ^ Бхадаурия, Виджай; Банниза, Сабина; Ванденберг, Альберт; Сельварадж, Гопалан; Вэй, Яндоу (1 сентября 2012 г.). «Аланин» . Сигнализация и поведение растений . 7 (9): 1206–1208. Бибкод : 2012PlSiB...7.1206B . дои : 10.4161/psb.21368 . ПМЦ 3489663 . ПМИД 22899049 .
- ^ Сент-Клер, Дина (2010). «Количественная устойчивость к болезням и локусы количественной устойчивости в селекции». Ежегодный обзор фитопатологии . 48 : 247–268. doi : 10.1146/annurev-phyto-080508-081904 . ISSN 0066-4286 . ПМИД 19400646 .
- ^ Jump up to: а б с Мотояма, Такаюки; Юн, Чунг Су; Осада, Хироюки (2021). «Биосинтез и биологическая функция вторичных метаболитов рисового гриба Pyricalia oryzae » . Журнал промышленной микробиологии и биотехнологии . 48 (9–10). дои : 10.1093/jimb/kuab058 . ПМЦ 8788799 . ПМИД 34379774 .
- ^ Jump up to: а б с Маршаль, Клеманс; Михалопулу, Василики А.; Цзоу, Чжоу; Чевик, Волкан; Саррис, Панайотис Ф. (2022). «Покажите мне свой идентификатор: иммунные рецепторы NLR с интегрированными доменами в растениях» . Очерки по биохимии . 66 (5): 527–539. дои : 10.1042/ebc20210084 . ПМЦ 9528084 . ПМИД 35635051 .
- ^ Чжоу, Цзюньхуэй; Пэн, Чжао; Лонг, Джуинг; и др. (2015). «Нацеливание на ген с помощью эффектора TAL PthXo2 выявляет загадочный ген устойчивости к бактериальной болезни риса» . Заводской журнал . 82 (4): 632–643. дои : 10.1111/tpj.12838 . ПМИД 25824104 . S2CID 29633821 .
Дальнейшее чтение
[ редактировать ]- Калифорнийское агентство по охране окружающей среды . Заражение посевов риса в трех округах привело к заключению соглашения об экстренном сжигании , 11 февраля 1998 г.
- СИММИТ . Что такое пшеничный взрыв? , 2019.
- Кадлец, РП. Биологическое оружие для ведения экономической войны , Air & Space Power Chronicles
- НФС . Микробный геном помогает победить разрушительную болезнь риса , 21 апреля 2005 г.
- Конгресс США . Свидетельство доктора Кеннета Алибека , 1999 г.
- «Что такое пшеничный взрыв?» . СИММИТ . 11 декабря 2019 г. Проверено 21 декабря 2020 г.
- «Что такое пшеничный взрыв? Как мне с ним справиться?» (PDF) . СИММИТ .
- Като Като, Ямагути, Томио; Нишихара Нисихара, Нацуки (1976). Идеальное состояние Pyricleria oryzae Cav. в культуре» . журнал фитопатологии . 42 (4). Японский « doi : 10.3186/ jjphytopath.42.507 ISSN 1882-0484 .
- «Взрыв (узел и шейка)» . Банк знаний о рисе . IRRI ( Международный институт исследования риса ). 15 августа 2017 г. Проверено 4 марта 2021 г.