Стиходактильный токсин
| ШК доменоподобный | |||
|---|---|---|---|
 Радужная мультипликационная диаграмма ( N-конец = синий, C-конец = красный) структуры раствора ЯМР токсина ShK. [ 1 ] Боковые цепи остатков цистеина , участвующих в дисульфидных связях, показаны в виде палочек, а атомы серы в этих связях окрашены в желтый цвет. | |||
| Идентификаторы | |||
| Символ | CS | ||
| Пфам | PF01549 | ||
| ИнтерПро | ИПР003582 | ||
| УМНЫЙ | SM00254 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1roo / SCOPe / СУПФАМ | ||
| TCDB | 8.Б.14 | ||
| Суперсемейство OPM | 296 | ||
| белок OPM | 2lg4 | ||
| |||
| Каппа-стихотоксин-She3a | |||
|---|---|---|---|
| Идентификаторы | |||
| Организм | |||
| Символ | ? | ||
| ЮниПрот | P29187 | ||
| |||
Токсин стиходактилии ( ShK , ShkT из 35 остатков ) представляет собой основной пептид из актинии Stichodactyla helianthus , который блокирует ряд калиевых каналов . Родственные пептиды образуют консервативное семейство белковых доменов, известное как домен ShkT . Другой хорошо изученный токсин семейства — BgK из Bunodosoma granulifera .
Аналог ШК под названием ШК-186 или Далазатид проходит испытания на людях в качестве средства лечения аутоиммунных заболеваний .
История
[ редактировать ]
Stichodactyla helianthus — вид актиний (тип: Cnidaria ), принадлежащий семейству Stichodactylidae . Helianthus происходит от греческих слов helios, означающих солнце, и anthos, означающих цветок, что соответствует общему названию вида «солнечный анемон». Он сидит сидя и использует мощные нейротоксины для защиты от своего основного хищника, лангуста . [ 2 ] Яд содержит, помимо других компонентов, многочисленные пептиды , блокирующие ионные каналы . В 1995 году группа под руководством Ольги Кастанеды и Эверта Карлссона выделила ShK, пептид из 35 остатков, блокирующий калиевые каналы, из S. helianthus . [ 3 ] В том же году Уильям Кем и его сотрудник Майкл Пеннингтон синтезировали и свернули ShK и показали, что он блокирует нейронов и лимфоцитов потенциал-зависимые калиевые каналы . [ 4 ] В 1996 году Рэй Нортон определил трехмерную структуру ШК. [ 1 ] В 2005–2006 годах Джордж Чанди, Кристин Битон и Майкл Пеннингтон разработали ШК-170 и ШК-186 (ШК-L5), селективные блокаторы K v 1.3 . [ 5 ] [ 6 ] ShK-186, теперь называемый Далазатид, был подвергнут испытаниям на людях в 2015-2017 годах Шоном Иадонато и Эриком Тарча как первый человеческий блокатор K v 1.3 для лечения аутоиммунных заболеваний. [ 7 ]
Структура
[ редактировать ]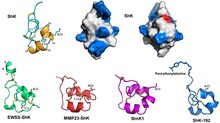
ShK сшит тремя дисульфидными мостиками : Cys3-Cys35, Cys12-Cys28 и Cys17-Cys32. В структуре раствора ShK обнаруживаются две короткие α-спирали, содержащие остатки 14–19 и 21–24; восемь N-концевых остатков принимают вытянутую конформацию, за которой следует пара переплетающихся витков, напоминающих 3 10 спираль ; С -концевой остаток Cys35 образует циклическую структуру почти «голова к хвосту» через дисульфидную связь с Cys3. [ 1 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ]

Филогенетические взаимоотношения доменов ShK и ShK
[ редактировать ]
База данных SMART в EMBL , по состоянию на май 2018 года, [ 13 ] перечисляет 3345 белковых доменов со структурным сходством с ShK в 1797 белках (от 1 до 8 доменов на белок), многие из которых есть у червей Caenorhabditis elegans и ядовитых змей . [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ] Большинство этих доменов находятся в металлопептидазах , тогда как другие находятся в пролил-4-гидроксилазах , тирозиназах , пероксидазах , оксидоредуктазах или белках, содержащих домены, подобные эпидермальному фактору роста , повторы типа тромбоспондина или трипсиноподобные домены сериновой протеазы . [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ] Единственными человеческими белками, содержащими ShK-подобные домены, являются MMP-23 (матриксная металлопротеаза 23) и MFAP-2 (связанный с микрофибриллами гликопротеин 2). [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ]
Цели канала
[ редактировать ]
Пептид ShK блокирует калий (K + ) ионные каналы K v 1.1 , K v 1.3 , K v 1.6 , K v 3.2 и K Ca 3.1 с наномолярной или пикомолярной активностью и не влияют на сердечный калиевый канал HERG ( K v 11.1 ). [ 19 ] [ 20 ] 1.1 нейронов Канал K v и канал K v 1.3 Т-лимфоцитов наиболее сильно ингибируются ShK. [ 8 ]
Конфигурация привязки в K + каналы
[ редактировать ]ШК и его аналоги являются блокаторами пор канала. Они связываются со всеми четырьмя субъединицами K + тетрамер канала путем взаимодействия с неглубоким «вестибюлем» у внешнего входа в пору канала. [ 5 ] [ 8 ] [ 9 ] [ 12 ] [ 21 ] [ 22 ] [ 19 ] Эти пептиды закрепляются во внешнем вестибюле посредством двух ключевых взаимодействий. Первым является Lys22, который выступает в пору канала и закупоривает ее, как «пробка в бутылке», и блокирует прохождение ионов калия через пору канала. [ 8 ] [ 23 ] [ 22 ] [ 19 ] Вторым является соседний Tyr23, который вместе с Lys22 образует «функциональную диаду», необходимую для блокировки каналов. [ 8 ] [ 9 ] [ 22 ] [ 23 ] [ 19 ] Многие К + пептиды, блокирующие каналы, содержат такую диаду лизина и соседнего ароматического или алифатического остатка. [ 19 ] [ 20 ] Немного К + пептиды, блокирующие каналы, лишены функциональной диады, но даже в этих пептидах лизин физически блокирует канал независимо от положения лизина в пептидной последовательности. [ 24 ] Дополнительные взаимодействия закрепляют ShK и его аналоги во внешнем преддверии и способствуют эффективности и селективности. [ 8 ] [ 9 ] [ 22 ] [ 23 ] [ 19 ] Например, Arg11 и Arg29 в ShK взаимодействуют с двумя остатками Asp386 в соседних субъединицах наружного преддверия K v 1.3 мыши (соответствует Asp433 в K v 1.3 человека). [ 8 ] [ 9 ] [ 22 ] [ 23 ] [ 19 ]
| Канал | ШК (ИК 50 ) | ШК-186 ( IC50 ) |
ШК-192 ( IC50 ) |
ШК-ЭВСС ( IC50 ) |
ШК-Ф6СА (ИК 50 ) | ШК-198 (ИК 50 ) | Домен ММП-23 ШК (IC 50 ) |
|---|---|---|---|---|---|---|---|
| К в 1.1 | 16-28 вечера | 7 нМ | 22 нМ | 5,4 нМ | 4 нМ | 159 вечера | 49 мкм |
| К в 1,2 | 10 нМ | 48 нМ | без даты | >100 нМ | >100 нМ | без даты | >100 мкм |
| К в 1,3 | 22-16 вечера | 70 вечера | 14:40 | 15:00 | 16:00 | 16:00 | 2,8 мкм |
| К в 1,6 | 14:00 | 18 нМ | 10,6 нМ | без даты | без даты | без даты | 400 нМ |
| К в 3,2 | 5 нМ | 20 нМ | 4,2 нМ | без даты | без даты | без даты | 49 мкм |
| К Са 3,1 | 30 нМ | 115 нМ | >100 нМ | >100 нМ | без даты | без даты | >100 мкм |
Аналоги, блокирующие канал Kv1.3
[ редактировать ]Было создано несколько аналогов ShK для повышения специфичности канала K v 1.3 по сравнению с нейронным каналом K v 1.1 и другими тесно связанными каналами.
- ШК-Дап 22 : Это был первый аналог, показавший некоторую степень специфичности для K v 1.3. Закрывающий поры лизин 22 ШК заменен на диаминопропионовую кислоту (Дап) в ШК-Дап 22 . [ 8 ] [ 22 ] [ 25 ] Dap представляет собой неприродный аналог лизина с более короткой длиной боковой цепи (2,5 Å от Cα ) , чем у лизина (6,3 Å). [ 26 ] Дап 22 взаимодействует с остатками, расположенными дальше во внешнем преддверии, в отличие от лизина 22 , который взаимодействует с фильтром избирательности канала. [ 22 ] Как следствие, ориентации ШК и ШК-Дап 22 в наружном вестибюле существенно различаются. [ 22 ] ШК-Дап 22 демонстрирует >20-кратную селективность в отношении K v 1,3 по близкородственным каналам в экспериментах с патч-клампом на целых клетках , [ 8 ] но в анализах равновесного связывания он связывает гетеротетрамеры K v 1.1-K v 1.2 почти с той же эффективностью, что и ShK, что не прогнозируется на основе изучения гомотетрамерных каналов K v 1.1 или K v 1.2. [ 25 ]
- ShK-F6CA: присоединение флуоресцеина к N-концу пептида через гидрофильный линкер AEEA (2-аминоэтокси-2-этоксиуксусная кислота; мини-ПЭГ ) привело к образованию пептида ShK-F6CA (флуоресцеин-6-карбоксил). , со 100-кратной специфичностью для K v 1,3 по сравнению с K v 1,1 и связанными каналами. [ 27 ] Присоединение тетраметилродамина или биотина через линкер AEEA к N-концу ShK не повышало специфичность для K v 1.3 по сравнению с K v 1.1. [ 27 ] Повышенную специфичность ShK-F6CA можно объяснить различиями в заряде: F6CA заряжен отрицательно; тетраметилродамин заряжен положительно; а биотин нейтрален. [ 27 ] Последующие исследования с другими аналогами показали, что отрицательно заряженный F6CA, вероятно, взаимодействует с остатками на турели канала K v 1.3, как показано для ShK-192 и ShK-EWSS. [ 9 ] [ 12 ]
- ШК-170, ШК-186, ШК-192 и ШК-ЭВСС: на базе ШК-Ф6СА были изготовлены дополнительные аналоги. Присоединение L- фосфотирозина к N-концу ShK через линкер AEEA привело к образованию пептида ShK-170 со 100-1000-кратной специфичностью к K v 1,3 по сравнению с родственными каналами. ШК-186 (он же SL5; он же Далазатид) идентичен ШК-170, за исключением того, что С-концевой карбоксил заменен амидом. ShK-186 блокирует K v 1,3 с IC 50 , равным 69 пМ, и проявляет ту же специфичность для K v 1,3 по близкородственным каналам, что и ShK-170. [ 19 ] L-фосфотирозин ShK-170 и ShK-186 быстро дефосфорилируется in vivo с образованием аналога ShK-198 со сниженной специфичностью к K v 1,3. [ 5 ] [ 28 ] [ 29 ] Для преодоления этой проблемы были разработаны ШК-192 и ШК-ЭВСС. В ShK-192 N-концевой L-фосфотирозин заменен негидролизуемым пара-фосфонофенилаланином (Ppa), а Met21 заменен неприродной аминокислотой норлейцином, чтобы избежать окисления метионина . [ 9 ] [ 29 ] В ShK-EWSS линкер AEEA и L-фосфотирозин заменены остатками глутаминовой кислоты (Е), триптофана (W) и двух серинов (S). [ 12 ] И ШК-192, и ШК-EWSS высокоспецифичны для K v 1.3 по родственным каналам.
- ShK-K18A: Моделирование стыковки и молекулярной динамики Kv 1.3 и Kv 1.1 с последующим моделированием зонтичного отбора проб проложило путь к селективному ингибитору Kv1.3 ShK-K18A. [ 30 ]
- ShK-родственные пептиды у паразитических червей: AcK1, пептид из 51 остатка из анкилостомы Ancylostoma caninum и Ancylostoma ceylanicum , и BmK1, C-концевой домен металлопротеазы из филяриального червя Brugia malayi , имеют спиральные структуры, очень напоминающие ShK. [ 31 ] AcK1 и BmK1 блокируют K v каналы 1.3 в наномолярно-микромолярных концентрациях и подавляют эффекторные Т-клетки памяти крысы , не затрагивая подмножества наивных и центральных Т-клеток памяти. [ 31 ] Кроме того, они подавляют выработку IFN-g Т-клетками человека и ингибируют реакцию гиперчувствительности замедленного типа, вызываемую эффекторными Т-клетками памяти, возвращающимися к коже. [ 31 ] Teladorsagiacircumcincta — экономически важный паразит, поражающий овец и коз. TcK6, белок из 90 остатков с C-концевым доменом, родственным ShK, активируется во время личиночной стадии обитания этого паразита в слизистой оболочке. [ 32 ] TcK6 вызывает умеренное подавление запускаемой тапсигаргином продукции IFN-g Т-клетками овцы, что позволяет предположить, что паразит использует этот белок для уклонения от иммунитета путем модуляции Т-клеток слизистой оболочки. [ 32 ]
Увеличение периода полувыведения из обращения
[ редактировать ]Из-за своей низкой молекулярной массы ШК и его аналоги склонны к быстрой элиминации почками. У крыс период полувыведения составляет ~6 мин для ШК-186 и ~11 мин для ШК-198, скорость клиренса ~950 мл/кг·мин. [ 28 ] У обезьян период полувыведения составляет ~12 минут для ШК-186 и ~46 минут для ШК-198, скорость клиренса ~80 мл/кг·мин. [ 28 ]
ПЭГилирование ShK: конъюгация полиэтиленгликоля (ПЭГ) с ShK[Q16K], аналогом ShK, увеличивала его молекулярную массу и тем самым снижала почечный клиренс и увеличивала период полувыведения из плазмы до 15 часов у мышей и 64 часов у яванских макак. [ 11 ] ПЭГилирование также может снизить иммуногенность и защитить пептид от протеолиза и неспецифической адсорбции на инертных поверхностях. ПЭГилированный ShK[Q16K] предотвратил экспериментальный аутоиммунный энцефаломиелит с адоптивной передачей у крыс, модель рассеянного склероза. [ 11 ]
- Конъюгация ShK с более крупными белками. Период полувыведения пептидов из циркуляции можно продлить путем связывания их с более крупными белками или белковыми доменами. [ 19 ] [ 33 ] [ 34 ] Путем скрининга комбинаторной библиотеки пептидов ShK были идентифицированы новые аналоги, которые при слиянии с C-концами IgG1-Fc сохраняли пикомолярную активность, эффективно подавляли гиперчувствительность замедленного типа in vivo и демонстрировали длительный период полувыведения из крови. [ 35 ]
- Пролонгированные эффекты, несмотря на быстрый клиренс из плазмы: исследования ОФЭКТ/КТ с 111 В-ДОТА-конъюгат ШК-186 у крыс и беличьих обезьян выявил медленное высвобождение из места инъекции и уровень в крови выше дозы, блокирующей каналы, в течение 2 и 7 дней соответственно. [ 28 ] Исследования Т-клеток периферической крови человека показали, что кратковременного воздействия ShK-186 было достаточно для подавления цитокиновых ответов. [ 28 ] Эти данные позволяют предположить, что ShK-186, несмотря на короткий период полувыведения из крови, может оказывать пролонгированный терапевтический эффект. У крыс пептид эффективен при лечении аутоиммунных заболеваний на животных моделях при введении от одного раза в день до одного раза в 3 дня. [ 28 ] У людей подкожных инъекций два раза в неделю достаточно для облегчения заболевания у пациентов с бляшечным псориазом. [ 7 ]
Доставка пептидов
[ редактировать ]Низкая молекулярная масса ШК и его аналогов в сочетании с их высокими изоэлектрическими точками делает маловероятной абсорбцию этих пептидов из желудка или кишечника после перорального приема. Возможна сублингвальная доставка. Флуоресцентный аналог ShK абсорбировался в кровоток в фармакологических концентрациях после сублингвального введения с мукоадгезивным гелем на основе хитозана, с усилителем проникновения цетримидом или без него. [ 36 ] Возможна также доставка пептида в виде аэрозоля через легкие или через кожу или в виде глазных капель. [ 37 ] [ 38 ] [ 39 ]
Модуляция функции Т-клеток
[ редактировать ]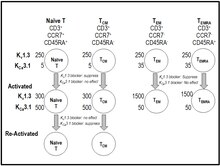
Во время активации Т-клеток кальций поступает в лимфоциты через депо-управляемые каналы CRAC (канал, активируемый высвобождением кальция), образованные в виде комплекса белков Orai и Stim. [ 40 ] [ 41 ] Повышение внутриклеточного кальция инициирует сигнальный каскад, завершающийся выработкой и пролиферацией цитокинов. [ 40 ] [ 41 ] К v 1,3 К + канал и активируемый кальцием K Ca 3,1 K + канал в Т-клетках способствует поступлению кальция в цитоплазму через CRAC, обеспечивая уравновешивающий отток катионов. [ 19 ] [ 40 ] [ 41 ] Блокада K v 1.3 деполяризует мембранный потенциал Т-клеток, подавляет передачу сигналов кальция и продукцию IL-2, но не экспрессию рецептора IL-2. [ 42 ] [ 43 ] [ 44 ] [ 45 ] [ 46 ] Блокаторы K v 1.3 не влияют на пути активации, независимые от повышения уровня внутриклеточного кальция (например, анти-CD28, IL-2). [ 42 ] [ 43 ] Экспрессия каналов K v 1.3 и K Ca 3.1 варьируется во время активации Т-клеток и дифференцировки в Т-клетки памяти. [ 19 ] [ 40 ] [ 41 ] [ 47 ] [ 48 ] Когда наивные Т-клетки и Т-клетки центральной памяти (TCM ) активируются, они повышают экспрессию KCa3.1 до ~500 на клетку без значительного изменения количества Kv1.3 . [ 19 ] [ 40 ] [ 41 ] [ 47 ] [ 48 ] Напротив, когда терминально дифференцированные подмножества эффекторной памяти ( TEM , T EMRA активируются [Т-эффекторная память, повторно экспрессирующая CD45RA]), они повышают уровень K v от 1,3 до 1500 на клетку без изменений в K Ca 3.1. [ 19 ] [ 40 ] [ 41 ] [ 47 ] [ 48 ] Количество каналов K v 1.3 увеличивается, а количество каналов K Ca 3.1 уменьшается по мере хронической активации Т-клеток. [ 40 ] [ 41 ] [ 47 ] [ 48 ] [ 49 ] В результате такой дифференциальной экспрессии блокаторы каналов K Ca 3.1 преимущественно подавляют функцию наивных и T CM клеток, тогда как ShK и его аналоги, избирательно ингибирующие каналы K v 1.3, преимущественно подавляют функцию хронически активированных эффекторных T клеток памяти (T ЭМ , ТЕМРА ). [ 19 ] [ 40 ] [ 41 ] [ 47 ] [ 48 ]
Особый интерес представляет большое количество аналогов ShK, разработанных в Amgen, которые подавляют выработку интерлейкина-2 и гамма-интерферона Т-клетками. [ 11 ] Этот ингибирующий эффект блокаторов K v 1.3 является частичным и зависит от силы стимуляции, при этом ингибирующая эффективность Т-клеток снижается при усиленной стимуляции анти-CD3/CD28. [ 50 ] Хронически активированный CD28 нулевой эффекторные Т-клетки памяти участвуют в аутоиммунных заболеваниях (например, волчанке, болезни Крона, ревматоидном артрите, рассеянном склерозе). [ 51 ] [ 52 ] [ 53 ]
Блокада каналов K v 1.3 в этих хронически активированных Т-клетках подавляет передачу сигналов кальция, выработку цитокинов (интерферон гамма, интерлейкин-2, интерлейкин 17) и пролиферацию клеток. [ 6 ] [ 19 ] [ 29 ] [ 30 ] [ 40 ] [ 41 ] [ 47 ] [ 48 ] Эффекторные Т-клетки памяти, являющиеся CD28. + невосприимчивы к подавлению блокаторами K v 1.3, когда они совместно стимулируются антителами против CD3 и против CD28, но чувствительны к подавлению при стимуляции только антителами против CD3. [ 48 ] In vivo ShK-186 парализует эффекторные Т-клетки памяти в месте воспалительной реакции гиперчувствительности замедленного типа и предотвращает активацию этих Т-клеток в воспаленной ткани. [ 54 ] Напротив, ShK-186 не влияет на хоминг и подвижность наивных и Т- CM- клеток в лимфатических узлах и внутри них, скорее всего, потому, что эти клетки экспрессируют канал K Ca 3.1 и, следовательно, защищены от эффекта блокады K v 1.3. [ 54 ]
Влияние на микроглию
[ редактировать ]K v 1.3 играет важную роль в активации микроглии. [ 55 ] [ 56 ] [ 57 ] [ 58 ] ShK-223, аналог ShK-186, уменьшал индуцированное липополисахаридом (ЛПС) образование фокальных спаек микроглией, обращал вспять индуцированное ЛПС ингибирование миграции микроглии и ингибировал индуцированную ЛПС активацию белка 1, содержащего домен EH (EHD1), белка участвуют в торговле микроглией. [ 59 ] Повышенная экспрессия K v 1.3 была зарегистрирована в микроглии бляшек Альцгеймера. [ 60 ] Ингибиторы K v 1.3 могут быть использованы при лечении болезни Альцгеймера, как сообщалось в исследовании, подтверждающем концепцию, в котором небольшая молекула блокатора K v 1.3 (PAP-1) облегчала симптомы, подобные болезни Альцгеймера, на мышиной модели AD. . [ 61 ]
Эффективность аналогов на животных моделях заболеваний человека
[ редактировать ]Экспериментальный аутоиммунный энцефаломиелит (ЭАЭ), модель рассеянного склероза
[ редактировать ]ШК, ШК-Дап 22 , ShK-170 и ПЭГилированный ShK-Q16K предотвращают ЭАЭ адоптивного переноса у крыс Льюиса, модели рассеянного склероза. [ 5 ] [ 11 ] Поскольку рассеянный склероз является рецидивирующим ремиттирующим заболеванием, ShK-186 и ShK-192 оценивали на модели ремиттирующего EAE на крысах DA (Dark Agouti). И предотвращали, и лечили заболевание при введении от одного раза в день до одного раза в три дня. [ 54 ] [ 28 ] [ 29 ] Таким образом, ингибиторы K v 1.3 эффективны при лечении заболеваний на крысиных моделях рассеянного склероза при введении отдельно. [ 11 ] [ 28 ] [ 62 ] [ 63 ] и терапевтическая эффективность, по-видимому, не подвергается риску из-за компенсаторной сверхэкспрессии каналов K Ca 3.1. [ 49 ] [ 64 ]
Пристан-индуцированный артрит (ПИА), модель ревматоидного артрита
[ редактировать ]ShK-186 был эффективен при лечении ПИА при ежедневном введении или через день. [ 5 ] [ 6 ] [ 54 ] [ 28 ] Ингибитор токсина скорпиона K V 1.3 также оказался эффективным в этой модели. [ 65 ] В обоих этих исследованиях одной блокады K v 1.3 было достаточно для облегчения заболевания, и одновременная блокада K Ca 3.1 не была необходимой, как предполагалось. [ 49 ] [ 64 ]
Крысиные модели атопического дерматита
[ редактировать ]Большинство инфильтрирующих Т-клеток в поражениях кожи у пациентов с атопическим дерматитом (АД) средней и тяжелой степени экспрессируют высокие уровни K v 1,3, что позволяет предположить, что ингибиторы K v 1,3 могут быть эффективны при лечении АД. [ 66 ] Моделью атопического дерматита считаются овальбумин-индуцированная гиперчувствительность замедленного типа и оксазолоновый дерматит. [ 66 ] [ 67 ] [ 68 ] [ 69 ] [ 70 ] ShK, ShK-170, ShK-186, ShK-192 и ShK-IgG-Fc были эффективны в модели гиперчувствительности замедленного типа, индуцированной овальбумином. [ 5 ] [ 6 ] [ 9 ] [ 54 ] [ 28 ] [ 29 ] [ 35 ] [ 66 ] [ 69 ] [ 70 ] [ 71 ] [ 72 ] [ 73 ] [ 74 ] [ 75 ] в то время как местный состав ShK-198 был эффективен при лечении дерматита, вызванного оксазолоном. [ 66 ] Даже там, где сообщалось, что компенсация с помощью каналов K Ca 3.1 подавляет блокировку K V 1.3, введение только ShK значительно подавляло гиперчувствительность замедленного типа в 2 из 3 исследований, хотя и умеренно. [ 49 ]
Псориаз
[ редактировать ]Псориаз — тяжелое аутоиммунное заболевание кожи, от которого страдают многие люди во всем мире. Несмотря на успех последних биологических препаратов в облегчении заболевания, поиск безопасных и эффективных лекарств от псориаза все еще ведется. Сообщалось, что ингибиторы K V 1.3 (ShK, PAP-1) лечат заболевание на мышиной модели псориазоформной (псориазоподобной) SCID (тяжелый комбинированный иммунодефицит). [ 76 ] В плацебо-контролируемом клиническом исследовании фазы 1b у пациентов с бляшечным псориазом ShK-186, вводимый два раза в неделю (30 или 60 мг/доза/пациент) путем подкожной инъекции, вызывал улучшение со статистически значимым снижением PASI (площадь и тяжесть псориаза). Индекс) балл между исходным уровнем и 32-м днем. [ 7 ] У этих пациентов также наблюдалось снижение уровней в плазме нескольких маркеров воспаления и снижение экспрессии маркеров активации Т-клеток на Т-клетках памяти периферической крови. [ 7 ]
Ожирение, вызванное диетой, и жировая болезнь печени
[ редактировать ]Ожирение и диабет являются основными проблемами здравоохранения во всем мире. Существует потребность в безопасных лекарствах от этих метаболических заболеваний. В мышиной модели ожирения, вызванного диетой, ShK-186 противодействовал негативным эффектам повышенного потребления калорий. Это уменьшило увеличение веса, ожирение и ожирение печени; снижение уровня холестерина, сахара, HbA1c, инсулина и лептина в крови; и повышенная периферическая чувствительность к инсулину. [ 77 ] Генетическая делеция гена K v 1.3 имеет тот же эффект, указывая на то, что эффект ShK-186 обусловлен блокадой K v 1.3. [ 78 ] [ 77 ] [ 79 ] [ 80 ] По крайней мере, два механизма способствуют терапевтическому эффекту ШК-186. Высококалорийная диета индуцировала экспрессию K v 1.3 в бурых жировых тканях. [ 77 ] Блокируя K v 1.3, ShK-186 удвоил поглощение глюкозы и увеличил β-окисление жирных кислот, гликолиз, синтез жирных кислот и разобщение экспрессии белка 1 бурым жиром. [ 77 ] В результате активации бурого жира потребление кислорода и расход энергии увеличились. [ 77 ] Диета при ожирении также индуцировала экспрессию K v 1.3 в печени, а ShK-186 вызывал глубокие изменения в энергетическом и липидном обмене в печени. ШК, его аналоги или другие блокаторы K v 1.3 могут быть использованы для контроля негативных последствий высококалорийных диет.
Возбуждение и анестезия
[ редактировать ]Механизмы общей анестезии включают множество молекулярных мишеней и путей, которые до конца не изучены. [ 81 ] Севофлуран — распространенный анестетик, используемый для индукции общей анестезии во время операции. [ 81 ] Крысы, постоянно подвергавшиеся воздействию севофлурана, теряют рефлекс выпрямления, что является показателем потери сознания. У этих крыс микроинфузия ShK в центральное медиальное ядро таламуса (ЦМТ) обратила вспять анестезию, вызванную севофлураном, у грызунов. [ 81 ] Крысы, получавшие ШК, полностью выздоровели (восстановили сознание), несмотря на постоянное воздействие севофлурана. [ 81 ] ШК-микроинфузия в соседние отделы мозга не оказала такого эффекта. [ 81 ] Севофлуран усиливал калиевые токи в ШМТ, а ШК и ШК-186 противодействовали этому эффекту. [ 81 ] Эти исследования показывают, что ShK-чувствительный K + каналы ШМТ важны для подавления возбуждения во время анестезии.
Предотвращение повреждения головного мозга после терапевтического облучения головного мозга
[ редактировать ]Облучение головного мозга используется для лечения опухолей головы, шеи и головного мозга, но это лечение сопряжено со значительным риском неврологического повреждения. Повреждение частично происходит из-за активации микроглии и микроглиально-опосредованного повреждения нейронов. Нейропротекторная терапия радиационно-индуцированного повреждения головного мозга по-прежнему ограничена. В мышиной модели облучения мозга ShK-170 обратил вспять неврологический дефицит и защитил нейроны от радиационно-индуцированного повреждения головного мозга путем подавления микроглии. [ 82 ]
Токсичность ШК и его аналогов
[ редактировать ]ШК и ШК-Дап 22
[ редактировать ]Пептид ShK имеет профиль низкой токсичности для мышей. ШК эффективен при лечении аутоиммунных заболеваний в дозе от 10 до 100 мг/кг массы тела. Его средняя паралитическая доза составляет примерно 25 мг/кг массы тела (на 250–2500 выше фармакологической дозы). У крыс индекс терапевтической безопасности превышает 75 раз. ШК-Дап 22 продемонстрировал более низкий профиль токсичности. [ 8 ] Доза 1,0 мг не вызывала гиперактивности, судорог или смертности у крыс. Средняя паралитическая доза ШК-Дап 22 составляет около 200 мг/кг массы тела (на 2000-20000 выше фармакологической дозы). [ 8 ] ПЭГилированный ShK[Q16K] не показал неблагоприятной токсичности у обезьян в течение нескольких месяцев. [ 11 ]
ShK-186/Dalazatide
[ редактировать ]ШК-186 также демонстрирует низкую токсичность у крыс. Ежедневное введение ШК-170 или ШК-186 (100 мкг/кг/день) путем подкожной инъекции в течение 4 недель крысам не вызывает каких-либо изменений показателей крови, биохимического состава крови или гистопатологии. [ 5 ] [ 6 ] [ 28 ] Благодаря подавлению только клеток TEM и TEMRA , ShK-186 не нарушал защитные иммунные реакции на вирус гриппа и хламидийную инфекцию у крыс, скорее всего, потому, что наивные клетки и клетки TCM, не затронутые блокадой K v 1.3, вызывали эффективные иммунные ответы. [ 54 ] ShK-186 обладает низкой иммуногенностью и не вызывает антитела против ShK у крыс, которым повторно вводили пептид. [ 6 ] Возможно, это связано с тем, что структура пептида с дисульфидными связями препятствует процессингу и презентации антигена антигенпрезентирующими клетками. ShK-186 также имеет сходство последовательности и структуры с ShK-подобным доменом матриксной металлопротеазы 23. [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ] что может заставить иммунную систему считать, что это нормальный белок в организме. ШК-186 был безопасен для приматов. В исследованиях фазы 1a и 1b на здоровых добровольцах ShK-186 хорошо переносился, не было отмечено никаких побочных эффектов 3 или 4 степени или лабораторных отклонений, а прогнозируемый диапазон воздействия препарата был достигнут. [ 7 ] Наиболее частыми нежелательными явлениями были временная легкая (степень 1) гипестезия и парестезия, затрагивающая руки, ноги или периоральную область. Также наблюдались легкие мышечные спазмы, чувствительность зубов и боль в месте инъекции. [ 7 ]
Функции ShK-подобных белков
[ редактировать ]ММП-23
[ редактировать ]ММП-23 принадлежит к семейству цинк- и кальций-зависимых матриксных металлопротеаз. Он закреплен в клеточной мембране с помощью N-концевого продомена и содержит три внеклеточных домена: домен каталитической металлопротеазы, домен ShK и домен иммуноглобулиноподобной молекулы клеточной адгезии ( Ig-CaM ). [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ] Продомен захватывает потенциалзависимый калиевый канал K V 1.3, но не близкородственный канал K V 1.2, в эндоплазматическом ретикулуме. [ 14 ] [ 15 ] [ 16 ] [ 17 ] Исследования с химерами показывают, что продомен взаимодействует с областью K V 1.3 от трансмембранного сегмента S5 до С-конца. [ 14 ] [ 15 ] [ 16 ] [ 17 ] ЯМР-исследования продомена выявили одну трансмембранную альфа-спираль, соединенную коротким линкером с околомембранной альфа-спиралью, которая связана с поверхностью мембраны. [ 14 ] [ 15 ] [ 16 ] [ 17 ] Продомен имеет топологическое сходство с белками (KCNE1, KCNE2, KCNE4), которые, как известно, захватывают калиевые каналы в секреторном пути, что указывает на общий механизм регуляции каналов. [ 14 ] [ 15 ] [ 16 ] [ 17 ] Каталитический домен MMP-23 демонстрирует структурную гомологию с каталитическими доменами других металлопротеаз и, вероятно, функционирует как эндопептидаза. Домен ShK MMP-23 расположен сразу после каталитического домена и соединен с доменом IgCAM коротким линкером, богатым пролином. Он имеет филогенетическое родство с токсинами морского анемона и доменами ICR-CRISP , будучи наиболее похожим на токсин BgK из морского анемона Bunodosoma granulifera . [ 14 ] [ 15 ] [ 16 ] [ 17 ] Этот домен ShK блокирует потенциалзависимые калиевые каналы (K V 1,6 > K V 1,3 > K V 1,1 = K V 3,2 > K v 1,4, с уменьшением эффективности) в наномолярном и низкомикромолярном диапазоне. [ 14 ] K V 1.3 необходим для поддержания передачи сигналов кальция во время активации Т-клеток человека. [ 19 ] [ 40 ] [ 47 ] Захватывая K V 1.3 в эндоплазматическом ретикулуме через продомен и блокируя канал K V 1.3 доменом ShK, MMP-23 может служить иммунной контрольной точкой для снижения чрезмерной активации Т-клеток во время иммунного ответа. В подтверждение, повышенная экспрессия MMP-23 в раковых клетках меланомы снижает количество инфильтрирующих опухоль лимфоцитов и связано с рецидивом рака и более короткими периодами выживаемости без прогрессирования. [ 17 ] Однако в меланомах экспрессия MMP-23 не коррелирует с экспрессией K v 1.3, что позволяет предположить, что вредный эффект MMP-23 при меланомах может быть не связан с ее K v 1.3. функцией модуляции каналов [ 17 ] С-концевой домен IgCAM MMP-23 имеет сходство последовательности с доменами IgCAM в белках, которые, как известно, опосредуют белок-белковые и белок-липидные взаимодействия (например , CDON , человеческий брат CDO , ROBO1-4 , гемицентин , NCAM1 и NCAM2 ). [ 14 ] [ 15 ] [ 16 ] [ 17 ] Таким образом, четыре домена MMP-23 могут действовать синергически, модулируя иммунные ответы in vivo .
Маб7
[ редактировать ]У самцов червей Caenorhabditis elegans отсутствие белка под названием Mab7 ( Q95Q39 ) приводит к уродливым сенсорным лучам, необходимым для спаривания. [ 63 ] Введение Mab7 этим самцам червей восстанавливает нормальное развитие нормальных сенсорных лучей. [ 63 ] Введение белков Mab7, лишенных ShK-домена, не корректирует дефект сенсорных лучей, что указывает на роль ShK-домена Mab7 в развитии сенсорных лучей. [ 63 ]
HMP2 и PMP1
[ редактировать ]HMP2 и PMP-1 представляют собой астациновые металлопротеиназы Cnidarian Hydra vulgaris и медузы Podocoryne carnea , которые содержат ShK-подобные домены на своих С-концах. [ 83 ] [ 17 ] Оба этих ShK-домена содержат критический лизин, закупоривающий поры, необходимый для K + блок каналов. [ 14 ] HMP2 играет решающую роль в регенерации стоп гидры . [ 83 ] в то время как PMP-1 обнаружен в органе питания медузы, а ShK-домен может парализовать добычу после ее проглатывания. [ 17 ]
CRISP
[ редактировать ]
Более отдаленным родством являются секреторные белки, богатые цистеином (CRISP), которые содержат ShK-подобный «богатый цистином домен», а также более крупный CAP-подобный «домен, связанный с патогенезом 1». [ 84 ] Эти белки участвуют в репродукции млекопитающих. [ 85 ] а также в ядах некоторых змей. [ 86 ] Считается, что в обоих случаях механизм включает ингибирование активности ионных каналов. [ 85 ]
Ссылки
[ редактировать ]![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2018 г. ) ( отчеты рецензента ):
Ши Чи Чанг; Саумья Баджадж; К. Джордж Чанди (1 июня 2018 г.). «Токсин ШК: история, структура и терапевтическое применение при аутоиммунных заболеваниях» (PDF) . Викижурнал науки . 1 (1): 3. дои : 10.15347/WJS/2018.003 . ISSN 2470-6345 . Викиданные Q55120296 .
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2018 г. ) ( отчеты рецензента ):
Ши Чи Чанг; Саумья Баджадж; К. Джордж Чанди (1 июня 2018 г.). «Токсин ШК: история, структура и терапевтическое применение при аутоиммунных заболеваниях» (PDF) . Викижурнал науки . 1 (1): 3. дои : 10.15347/WJS/2018.003 . ISSN 2470-6345 . Викиданные Q55120296 .
- ^ Перейти обратно: а б с д ПДБ : 1РОО ; Тюдор Дж. Э., Паллаги П. К., Пеннингтон М. В., Нортон Р. С. (апрель 1996 г.). «Структура раствора токсина ШК, нового ингибитора калиевых каналов из актинии». Структурная биология природы . 3 (4): 317–20. дои : 10.1038/nsb0496-317 . ПМИД 8599755 . S2CID 9180663 .
- ^ Нортон Р.С., Пеннингтон М.В., Вульф Х. (декабрь 2004 г.). «Блокада калиевых каналов токсином актинии ШК для лечения рассеянного склероза и других аутоиммунных заболеваний». Современная медицинская химия . 11 (23): 3041–52. дои : 10.2174/0929867043363947 . ПМИД 15578998 .
- ^ Кастаньеда О, Сотолонго В, Амор А.М., Стеклин Р., Андерсон А.Дж., Харви А.Л., Энгстрем А., Вернстедт К., Карлссон Е (май 1995 г.). «Характеристика токсина калиевого канала карибской актинии Stichodactyla helianthus». Токсикон . 33 (5): 603–13. дои : 10.1016/0041-0101(95)00013-C . ПМИД 7660365 .
- ^ Пеннингтон М.В., Бирнс М.Е., Зайденберг И., Хайтин И., де Частоне Дж., Крафте Д.С., Хилл Р., Махнир В.М., Вольберг В.А., Горчица В. (ноябрь 1995 г.). «Химический синтез и характеристика токсина ШК: мощного ингибитора калиевых каналов из актинии». Международный журнал исследований пептидов и белков . 46 (5): 354–8. дои : 10.1111/j.1399-3011.1995.tb01068.x . ПМИД 8567178 .
- ^ Перейти обратно: а б с д и ж г Битон С., Пеннингтон М.В., Вульф Х., Сингх С., Ньюджент Д., Кроссли Г., Хайтин И., Калабрези П.А., Чен С.И., Гутман Г.А., Чанди К.Г. (апрель 2005 г.). «Нацеливание на эффекторные Т-клетки памяти с помощью селективного пептидного ингибитора каналов Kv1.3 для терапии аутоиммунных заболеваний» . Молекулярная фармакология . 67 (4): 1369–81. дои : 10.1124/моль.104.008193 . ПМЦ 4275123 . ПМИД 15665253 .
- ^ Перейти обратно: а б с д и ж Битон С., Вульф Х., Стандифер Н.Э., Азам П., Маллен К.М., Пеннингтон М.В. и др. (ноябрь 2006 г.). «Каналы Kv1.3 являются терапевтической мишенью для аутоиммунных заболеваний, опосредованных Т-клетками» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (46): 17414–9. Бибкод : 2006PNAS..10317414B . дои : 10.1073/pnas.0605136103 . ПМК 1859943 . ПМИД 17088564 .
- ^ Перейти обратно: а б с д и ж Тарча Э.Дж., Олсен К.М., Пробст П., Пекхэм Д., Муньос-Элиас Э.Дж., Крюгер Дж.Г., Иадонато С.П. (июль 2017 г.). «Безопасность и фармакодинамика далазатида, ингибитора каналов Kv1.3, при лечении бляшечного псориаза: рандомизированное исследование фазы 1b» . ПЛОС ОДИН . 12 (7): e0180762. Бибкод : 2017PLoSO..1280762T . дои : 10.1371/journal.pone.0180762 . ПМК 5516987 . ПМИД 28723914 .
- ^ Перейти обратно: а б с д и ж г час я дж к Калман К., Пеннингтон М.В., Ланиган М.Д., Нгуен А., Рауэр Х., Махнир В., Пашетто К., Кем В.Р., Гриссмер С., Гутман Г.А., Кристиан Э.П., Кахалан М.Д., Нортон Р.С., Чанди К.Г. (декабрь 1998 г.). «ShK-Dap22, мощный Kv1.3-специфичный иммуносупрессивный полипептид» . Журнал биологической химии . 273 (49): 32697–707. дои : 10.1074/jbc.273.49.32697 . ПМИД 9830012 .
- ^ Перейти обратно: а б с д и ж г час Пеннингтон М.В., Битон С., Галеа К.А., Смит Б.Дж., Чи В., Монаган К.П., Гарсия А., Рангараджу С., Джуффрида А., Планк Д., Кроссли Г., Ньюджент Д., Хайтин И., Лефьевр Ю., Пещенко И., Диксон С., Чаухан С. , Орзель А., Иноуэ Т., Ху X, Мур Р.В., Нортон Р.С., Чанди К.Г. (апрель 2009 г.). «Разработка стабильного и селективного пептидного блокатора канала Kv1.3 в Т-лимфоцитах» . Молекулярная фармакология . 75 (4): 762–73. дои : 10.1124/моль.108.052704 . ПМЦ 2684922 . ПМИД 19122005 .
- ^ Пеннингтон М.В., Харунур Рашид М., Таджья Р.Б., Битон С., Куюкак С., Нортон Р.С. (ноябрь 2012 г.). «Амидированный по С-концу аналог ShK является мощным и селективным блокатором потенциалзависимого калиевого канала Kv1.3» . Письма ФЭБС . 586 (22): 3996–4001. Бибкод : 2012FEBSL.586.3996P . дои : 10.1016/j.febslet.2012.09.038 . ПМК 3496055 . ПМИД 23063513 .
- ^ Перейти обратно: а б с д и ж г Мюррей Дж.К., Цянь YX, Лю Б., Эллиотт Р., Арал Дж., Пак С., Чжан Х., Стенкилссон М., Сальерс К., Роуз М., Ли Х., Ю С., Эндрюс К.Л., Коломберо А., Вернер Дж., Гайда К., Сикмиер Э.А. , Миу П., Итано А., Макгиверн Дж., Гегг К.В., Салливан Дж.К., Миранда Л.П. (сентябрь 2015). «Фармацевтическая оптимизация пептидных токсинов для мишеней ионных каналов: мощные, селективные и долгоживущие антагонисты Kv1.3». Журнал медицинской химии . 58 (17): 6784–802. doi : 10.1021/acs.jmedchem.5b00495 . ПМИД 26288216 . S2CID 28397762 .
- ^ Перейти обратно: а б с д Чанг С.К., Хук Р., Чхабра С., Битон С., Пеннингтон М.В., Смит Б.Дж., Нортон Р.С. (июнь 2015 г.). «Удлиненные по N-концу аналоги токсина канала K⁺ из Stichodactyla helianthus как мощные и селективные блокаторы потенциалзависимого калиевого канала Kv1.3» . Журнал ФЭБС . 282 (12): 2247–59. дои : 10.1111/февраль 13294 . ПМЦ 4472561 . ПМИД 25864722 .
- ^ «SMART: аннотация домена ШКТ» . smart.embl-heidelberg.de . Проверено 16 мая 2018 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Рангараджу С., Ху К.К., Фенг З.П., Кроссли Г., Ньюджент Д., Хайтин И., Чи В., Фам С., Калабрези П., Пеннингтон М.В., Нортон Р.С., Чанди К.Г. (март 2010 г.). «Модуляция калиевого канала с помощью токсинового домена в матриксной металлопротеазе 23» . Журнал биологической химии . 285 (12): 9124–36. дои : 10.1074/jbc.M109.071266 . ПМЦ 2838332 . ПМИД 19965868 .
- ^ Перейти обратно: а б с д и ж г час я дж к Нгуен Х.М., Галеа К.А., Шмунк Г., Смит Б.Дж., Эдвардс Р.А., Нортон Р.С., Чанди К.Г. (март 2013 г.). «Внутриклеточный транспорт калиевого канала KV1.3 регулируется продоменом матриксной металлопротеазы» . Журнал биологической химии . 288 (9): 6451–64. дои : 10.1074/jbc.M112.421495 . ПМК 3585079 . ПМИД 23300077 .
- ^ Перейти обратно: а б с д и ж г час я дж к Галеа К.А., Нгуен Х.М., Джордж Чанди К., Смит Б.Дж., Нортон Р.С. (апрель 2014 г.). «Доменная структура и функция матриксной металлопротеазы 23 (MMP23): роль в транспортировке калиевых каналов» . Клеточные и молекулярные науки о жизни . 71 (7): 1191–210. дои : 10.1007/s00018-013-1431-0 . ПМЦ 11113776 . ПМИД 23912897 . S2CID 8113220 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Мугк Д., да Силва И.П., Ма М.В., Фридман Э.Б., де Миера Э.В., Дарвишян Ф., Скэнлон П., Перес-Гарсия А., Павлик А.С., Бхардвадж Н., Христос П.Дж., Осман И., Крогсгаард М. (декабрь 2014 г.). «Экспрессия матриксной металлопротеиназы-23 в меланоме связана с притупленным опухолевым иммунитетом и плохой реакцией на иммунотерапию» . Журнал трансляционной медицины . 12 : 342. дои : 10.1186/s12967-014-0342-7 . ПМЦ 4272770 . ПМИД 25491880 .
- ^ Перейти обратно: а б с д и Пан Т., Грегер Х., Шмид В., Спринг Дж. (июль 1998 г.). «Домен гомологии токсина в астацин-подобной металлопротеиназе медузы Podocoryne carnea, играющий двойную роль в пищеварении и развитии». Гены развития и эволюция . 208 (5): 259–66. дои : 10.1007/s004270050180 . ПМИД 9683741 . S2CID 13562676 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Чанди К.Г., Нортон РС (июнь 2017 г.). «Каналы v1.3 в Т-клетках как средство лечения аутоиммунных заболеваний». Современное мнение в области химической биологии . 38 : 97–107. дои : 10.1016/j.cbpa.2017.02.015 . ПМИД 28412597 .
- ^ Перейти обратно: а б Гилкин Б., Брауд С., Эрикссон М.А., Ру Б., Бейли Т.Д., Прист Б.Т., Гарсия М.Л., Менез А., Гаспарини С. (июль 2005 г.). «Вариабельный остаток в порах каналов Kv1 имеет решающее значение для высокого сродства блокаторов из морских анемонов и скорпионов» . Журнал биологической химии . 280 (29): 27093–102. дои : 10.1074/jbc.M413626200 . ПМИД 15890656 .
- ^ Пеннингтон М.В., Махнир В.М., Хайтин И., Зайденберг И., Бирнс М.Е., Кем В.Р. (декабрь 1996 г.). «Необходимая поверхность связывания для взаимодействия токсина ShK с калиевыми каналами мозга крысы». Биохимия . 35 (51): 16407–11. дои : 10.1021/bi962463g . ПМИД 8987971 .
- ^ Перейти обратно: а б с д и ж г час Ланиган, доктор медицинских наук, Калман К., Лефьевр Ю., Пеннингтон М.В., Чанди К.Г., Нортон Р.С. (октябрь 2002 г.). «Мутация критического лизина в токсине ShK меняет конфигурацию его связывания в области преддверия пор потенциалзависимого калиевого канала, Kv1.3». Биохимия . 41 (40): 11963–71. дои : 10.1021/bi026400b . ПМИД 12356296 .
- ^ Перейти обратно: а б с д Рауэр Х., Пеннингтон М., Кахалан М., Чанди К.Г. (июль 1999 г.). «Структурная консервация пор кальций-активируемых и потенциалзависимых калиевых каналов, определяемая токсином морского анемона» . Журнал биологической химии . 274 (31): 21885–92. дои : 10.1074/jbc.274.31.21885 . ПМИД 10419508 .
- ^ Стелинг Э.Г., Сфорса М.Л., Занчин Н.И., Ояма С., Пиньятелли А., Беллуцци О., Полверини Е., Корсини Р., Списни А., Пертинез Т.А. (март 2012 г.). «Рассмотрение взаимодействия каналов токсин-K(+). Подсказки из структурной и функциональной характеристики токсина α-KTx Tc32, блокатора каналов Kv1.3». Биохимия . 51 (9): 1885–94. дои : 10.1021/bi201713z . hdl : 11381/2435205 . ПМИД 22332965 .
- ^ Перейти обратно: а б Миддлтон Р.Э., Санчес М., Линде А.Р., Бугианеси Р.М., Дай Дж., Феликс Дж.П., Копрак С.Л., Старух М.Дж., Бругера М., Кокс Р., Гош А., Хванг Дж., Джонс С., Колер М., Слотер Р.С., Макманус О.Б., Качоровски Г.Дж. , Гарсия М.Л. (ноябрь 2003 г.). «Замена одного остатка в пептиде Stichodactyla helianthus, ShK-Dap22, открывает новый фармакологический профиль». Биохимия . 42 (46): 13698–707. дои : 10.1021/bi035209e . ПМИД 14622016 .
- ^ Айяр Дж., Рицци Дж.П., Гутман Г.А., Чанди К.Г. (декабрь 1996 г.). «Сигнатурная последовательность потенциалзависимых калиевых каналов выступает во внешний вестибюль» . Журнал биологической химии . 271 (49): 31013–6. дои : 10.1074/jbc.271.49.31013 . ПМИД 8940091 .
- ^ Перейти обратно: а б с Битон С., Вульф Х., Сингх С., Боцко С., Кроссли Г., Гутман Г.А., Кахалан М.Д., Пеннингтон М., Чанди К.Г. (март 2003 г.). «Новый флуоресцентный токсин для обнаружения и исследования активации канала Kv1.3 в хронически активированных Т-лимфоцитах» . Журнал биологической химии . 278 (11): 9928–37. дои : 10.1074/jbc.M212868200 . ПМИД 12511563 .
- ^ Перейти обратно: а б с д и ж г час я дж к Тарча Э.Дж., Чи В., Муньос-Элиас Э.Дж., Бейли Д., Лондоно Л.М., Упадьяй С.К., Нортон К., Бэнкс А., Чонг И., Нгуен Х., Ху Х., Рупперт Г.В., Боли С.Е., Слотер Р., Сэмс Дж., Кнапп Б., Кентала Д., Хансен З., Пеннингтон М.В., Битон С., Чанди К.Г., Иадонато С.П. (сентябрь 2012). «Долговечный фармакологический ответ от пептида ShK-186, специфического ингибитора канала Kv1.3, который подавляет медиаторы Т-клеток аутоиммунных заболеваний» . Журнал фармакологии и экспериментальной терапии . 342 (3): 642–53. дои : 10.1124/jpet.112.191890 . ПМЦ 3422530 . ПМИД 22637724 .
- ^ Перейти обратно: а б с д и Чи В., Пеннингтон М.В., Нортон Р.С., Тарча Э.Дж., Лондоно Л.М., Симс-Фэи Б., Упадьяй С.К., Лейки Дж.Т., Иадонато С., Вульф Х., Битон С., Чанди К.Г. (март 2012 г.). «Разработка токсина морского анемона как иммуномодулятора для терапии аутоиммунных заболеваний» . Токсикон . 59 (4): 529–46. дои : 10.1016/j.токсикон.2011.07.016 . ПМЦ 3397671 . ПМИД 21867724 .
- ^ Перейти обратно: а б Рашид М.Х., Хайнцельманн Г., Хук Р., Таджья Р.Б., Чанг С.К., Чхабра С., Пеннингтон М.В., Битон С., Нортон Р.С., Куюкак С. (ноябрь 2013 г.). «Мощный и селективный пептидный блокатор канала Kv1.3: прогноз на основе моделирования свободной энергии и экспериментальное подтверждение» . ПЛОС ОДИН . 8 (11): е78712. Бибкод : 2013PLoSO...878712R . дои : 10.1371/journal.pone.0078712 . ПМК 3820677 . ПМИД 24244345 .
- ^ Перейти обратно: а б с Чхабра С., Чанг С.К., Нгуен Х.М., Хук Р., Таннер М.Р., Лондоно Л.М., Эстрада Р., Дхаван В., Чаухан С., Упадхьяй С.К., Гиндин М., Хотез П.Дж., Валенсуэла Дж.Г., Моханти Б., Сворбрик Дж.Д., Вульф Х., Иадонато С.П. , Гутман Г.А., Битон С., Пеннингтон М.В., Нортон Р.С., Чанди К.Г. (сентябрь 2014 г.). «Имуномодулирующие пептиды, блокирующие каналы Kv1.3, из паразитических червей: значение для аутоиммунных заболеваний» . Журнал ФАСЭБ . 28 (9): 3952–64. дои : 10.1096/fj.14-251967 . ПМК 4139903 . ПМИД 24891519 .
- ^ Перейти обратно: а б Макнилли Т.Н., Фрю Д., Берджесс С.Т., Райт Х., Бартли DJ, Бартли Ю., Нисбет А.Дж. (август 2017 г.). «Специфическая для ниши экспрессия генов у паразитических нематод; повышенная экспрессия иммуномодуляторов в личинках Teladorsagiacircumcincta, полученных из слизистой оболочки хозяина» . Научные отчеты . 7 (1): 7214. Бибкод : 2017НатСР...7.7214М . дои : 10.1038/s41598-017-07092-0 . ПМЦ 5543109 . ПМИД 28775251 .
- ^ Эдвардс В., Фунг-Леунг В.П., Хуан С., Чи Е, Ву Н., Лю Й., Махер М.П., Бонстил Р., Коннор Дж., Феллоуз Р., Гарсия Е., Ли Дж., Лу Л., Нго К., Скотт Б., Чжоу Х., Суонсон Р.В., Викенден, AD (август 2014 г.). «Нацеливание на ионный канал Kv1.3 с помощью пептидов яда скорпиона, обеспечивающих эффективность, селективность и период полураспада» . Журнал биологической химии . 289 (33): 22704–14. дои : 10.1074/jbc.M114.568642 . ПМЦ 4132777 . ПМИД 24939846 .
- ^ Ван Р.Э., Ван И., Чжан И., Габрелоу С., Чжан И., Чи В., Фу К., Луо Икс, Ван Д., Джозеф С., Джонсон К., Чаттерджи А.К., Райт Т.М., Нгуен-Тран В.Т., Тейхаро Дж., Теофилопулос А.Н., Шульц П.Г., Ван Ф. (октябрь 2016 г.). «Рациональный дизайн антитела, блокирующего каналы Kv1.3, в качестве селективного иммунодепрессанта» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (41): 11501–11506. Бибкод : 2016PNAS..11311501W . дои : 10.1073/pnas.1612803113 . ПМК 5068325 . ПМИД 27663736 .
- ^ Перейти обратно: а б Чжан Х., Ду М., Се Дж., Лю Х., Сунь Дж., Ван В., Синь Х., Поссани Л.Д., Йеа К., Лернер Р.А. (август 2016 г.). «Аутокринный выбор лекарств, воздействующих на ионные каналы, из комбинаторных библиотек пептидов яда». Ангеванде Хеми . 55 (32): 9306–10. дои : 10.1002/anie.201603052 . ПМИД 27197631 .
- ^ Джин Л., Бойд Б.Дж., Уайт П.Дж., Пеннингтон М.В., Нортон Р.С., Николаццо Дж.А. (февраль 2015 г.). «Доставка мощного пептида в слизистую оболочку щеки приводит к терапевтически значимым концентрациям в плазме для лечения аутоиммунных заболеваний». Журнал контролируемого выпуска . 199 : 37–44. дои : 10.1016/j.jconrel.2014.12.001 . ПМИД 25482338 .
- ^ Джин Л., Чжоу К.Т., Чан Х.К., Ларсон И.С., Пеннингтон М.В., Моралес Р.А., Бойд Б.Дж., Нортон Р.С., Николаццо Дж.А. (февраль 2016 г.). «Легочная доставка пептида, блокирующего Kv1.3 HsTX1 [R14A] для лечения аутоиммунных заболеваний» . Журнал фармацевтических наук . 105 (2): 650–656. дои : 10.1016/j.xphs.2015.10.025 . ПМИД 26869426 .
- ^ WO 2015013330 , Iadonato & Munoz, «Офтальмологическое применение терапевтических пептидов на основе токсинов и их фармацевтических композиций»
- ^ WO 2016112208 , Иадонато; Тарча и Люстиг, «Местное применение пептидов, блокирующих каналы kv1.3, для лечения воспалений кожи»
- ^ Перейти обратно: а б с д и ж г час я дж Кахалан, доктор медицинских наук, Чанди К.Г. (сентябрь 2009 г.). «Функциональная сеть ионных каналов в Т-лимфоцитах» . Иммунологические обзоры . 231 (1): 59–87. дои : 10.1111/j.1600-065x.2009.00816.x . ПМК 3133616 . ПМИД 19754890 .
- ^ Перейти обратно: а б с д и ж г час я Феске С., Вульф Х., Скольник Е.Ю. (март 2015 г.). «Ионные каналы во врожденном и адаптивном иммунитете» . Ежегодный обзор иммунологии . 33 (1): 291–353. doi : 10.1146/annurev-immunol-032414-112212 . ПМЦ 4822408 . ПМИД 25861976 .
- ^ Перейти обратно: а б Лин К.С., Болц Р.К., Блейк Дж.Т., Нгуен М., Таленто А., Фишер П.А., Спрингер М.С., Сигал Н.Х., Слотер Р.С., Гарсия М.Л. (март 1993 г.). «Потенциал-управляемые калиевые каналы регулируют кальций-зависимые пути, участвующие в активации Т-лимфоцитов человека» . Журнал экспериментальной медицины . 177 (3): 637–45. дои : 10.1084/jem.177.3.637 . ПМК 2190940 . ПМИД 7679705 .
- ^ Перейти обратно: а б Чанди К.Г., ДеКурси Т.Э., Кахалан, доктор медицинских наук, Маклафлин С., Гупта С. (август 1984 г.). «Для активации Т-лимфоцитов человека необходимы потенциал-управляемые калиевые каналы» . Журнал экспериментальной медицины . 160 (2): 369–85. дои : 10.1084/jem.160.2.369 . ПМК 2187449 . ПМИД 6088661 .
- ^ Леонард Р.Дж., Гарсия М.Л., Слотер Р.С., Рубен Дж.П. (ноябрь 1992 г.). «Селективные блокаторы потенциал-зависимых К+-каналов деполяризуют Т-лимфоциты человека: механизм антипролиферативного действия харибдотоксина» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (21): 10094–8. Бибкод : 1992PNAS...8910094L . дои : 10.1073/pnas.89.21.10094 . ПМЦ 50284 . ПМИД 1279670 .
- ^ Дефариас Ф.П., Стивенс С.П., Леонард Р.Дж. (1995). «Стабильная экспрессия калиевых каналов Kv1.3 человека сбрасывает мембранный потенциал покоя культивируемых клеток млекопитающих». Рецепторы и каналы . 3 (4): 273–81. ПМИД 8834000 .
- ^ Верхойген Дж. А., Вийверберг Х. П., Оортгисен М., Кахалан, доктор медицинских наук (июнь 1995 г.). «Вольт-управляемые и Ca(2+)-активируемые K+-каналы в интактных Т-лимфоцитах человека. Неинвазивные измерения мембранных токов, мембранного потенциала и внутриклеточного кальция» . Журнал общей физиологии . 105 (6): 765–94. дои : 10.1085/jgp.105.6.765 . ПМК 2216960 . ПМИД 7561743 .
- ^ Перейти обратно: а б с д и ж г Вульф Х., Калабрези П.А., Элли Р., Юн С., Пеннингтон М., Битон С., Чанди К.Г. (июнь 2003 г.). «Потенциал-управляемый канал Kv1.3 K(+) в эффекторных Т-клетках памяти как новая мишень для рассеянного склероза» . Журнал клинических исследований . 111 (11): 1703–13. дои : 10.1172/jci16921 . ПМК 156104 . ПМИД 12782673 .
- ^ Перейти обратно: а б с д и ж г Ху Л., Пеннингтон М., Цзян К., Уортенби К.А., Калабрези П.А. (октябрь 2007 г.). «Характеристика функциональных свойств потенциалзависимого калиевого канала Kv1.3 в CD4+ Т-лимфоцитах человека» . Журнал иммунологии . 179 (7): 4563–70. дои : 10.4049/jimmunol.179.7.4563 . ПМИД 17878353 .
- ^ Перейти обратно: а б с д Чан Э.Ю., Ли Т., Джит С., Пэн И., Чжан Дж., Ли В.П., ДеВосс Дж., Каплази П., Чен Дж., Варминг С., Хакос Д.Х., Мукунд С., Кот СМ, Гроган Дж.Л. (март 2017 г.). «Калиевые каналы Kv1.3 и KCa3.1 совместно и компенсаторно регулируют антигенспецифические функции Т-клеток памяти» . Природные коммуникации . 8 : 14644. Бибкод : 2017NatCo...814644C . дои : 10.1038/ncomms14644 . ПМЦ 5337993 . ПМИД 28248292 .
- ^ Фунг-Люнг В.П., Эдвардс В., Лю Й., Нго К., Ангсана Дж., Кастро Дж., Ву Н., Лю Икс, Суонсон Р.В., Викенден А.Д. (20 января 2017 г.). «Подмножество Т-клеток и зависящая от силы стимуляции модуляция активации Т-клеток блокаторами Kv1.3» . ПЛОС ОДИН . 12 (1): e0170102. Бибкод : 2017PLoSO..1270102F . дои : 10.1371/journal.pone.0170102 . ПМК 5249144 . ПМИД 28107393 .
- ^ Намекава Т., Снайдер М.Р., Йен Дж.Х., Геринг Б.Е., Лейбсон П.Дж., Вейанд К.М., Горонзи Дж.Дж. (июль 2000 г.). «Рецепторы, активирующие киллерные клетки, действуют как костимулирующие молекулы на CD4+CD28null Т-клетках, клонально размножающихся при ревматоидном артрите» . Журнал иммунологии . 165 (2): 1138–45. дои : 10.4049/jimmunol.165.2.1138 . ПМИД 10878393 .
- ^ Маркович-Плесе С., Кортезе И., Вандингер К.П., МакФарланд Х.Ф., Мартин Р. (октябрь 2001 г.). «CD4+CD28-независимые от костимуляции Т-клетки при рассеянном склерозе» . Журнал клинических исследований . 108 (8): 1185–94. дои : 10.1172/jci12516 . ПМК 209525 . ПМИД 11602626 .
- ^ Гарсиа де Тена Х, Мансано Л., Леал Х.С., Сан-Антонио Э., Суальдеа В., Альварес-Мон М. (март 2004 г.). «У пациентов с активной болезнью Крона наблюдается характерное увеличение циркулирующих CD4+CD45RO+CD28null Т-клеток памяти». Журнал клинической иммунологии . 24 (2): 185–96. дои : 10.1023/B:JOCI.0000019784.20191.7f . ПМИД 15024186 . S2CID 12368149 .
- ^ Перейти обратно: а б с д и ж Матеу М.П., Битон С., Гарсия А., Чи В., Рангараджу С., Сафрина О., Монаган К., Уэмура М.И., Ли Д., Пал С., де ла Маза Л.М., Монуки Е., Флюгель А., Пеннингтон М.В., Паркер И., Чанди К.Г., Кахалан, доктор медицинских наук (октябрь 2008 г.). «Визуализация эффекторных Т-клеток памяти во время реакции гиперчувствительности замедленного типа и подавления блокировкой канала Kv1.3» . Иммунитет . 29 (4): 602–14. doi : 10.1016/j.immuni.2008.07.015 . ПМЦ 2732399 . ПМИД 18835197 .
- ^ Ханна Р., Рой Л., Чжу X, Шлихтер LC (апрель 2001 г.). «К+-каналы и микроглиальный респираторный взрыв». Американский журнал физиологии. Клеточная физиология . 280 (4): C796–806. дои : 10.1152/ajpcell.2001.280.4.C796 . ПМИД 11245596 . S2CID 12094438 .
- ^ Фордайс CB, Джагасия Р., Чжу X, Шлихтер LC (август 2005 г.). «Каналы микроглии Kv1.3 способствуют их способности убивать нейроны» . Журнал неврологии . 25 (31): 7139–49. doi : 10.1523/jneurosci.1251-05.2005 . ПМК 6725234 . ПМИД 16079396 .
- ^ Нгуен Х.М., Грёсингер Э.М., Хориучи М., Дэвис К.В., Джин Л.В., Маэзава И., Вульф Х. (январь 2017 г.). «Дифференциальная экспрессия Kv1.3, KCa3.1 и Kir2.1 в «классически» и «альтернативно» активированной микроглии» . Глия . 65 (1): 106–121. дои : 10.1002/glia.23078 . ПМК 5113690 . ПМИД 27696527 .
- ^ Нгуен Х.М., Бломстер Л.В., Кристоферсен П., Вульф Х. (июль 2017 г.). «Экспрессия и функция калиевых каналов в микроглии: пластичность и возможные видовые вариации» . Каналы . 11 (4): 305–315. дои : 10.1080/19336950.2017.1300738 . ПМЦ 5555259 . ПМИД 28277939 .
- ^ Рангараджу С., Раза С.А., Пеннати А, Денг К., Даммер Э.Б., Дуонг Д., Пеннингтон М.В., Тэнси М.Г., Ла Дж.Дж., Бетарбет Р., Сейфрид Н.Т., Леви А.И. (июнь 2017 г.). «Системный фармакологический подход к выявлению новых механизмов, зависящих от канала Kv1.3, при активации микроглии» . Журнал нейровоспаления . 14 (1): 128. дои : 10.1186/s12974-017-0906-6 . ПМЦ 5485721 . ПМИД 28651603 .
- ^ Рангараджу С., Гиринг М., Джин Л.В., Леви А. (01.01.2015). «Калиевый канал Kv1.3 высоко экспрессируется микроглией при болезни Альцгеймера у человека» . Журнал болезни Альцгеймера . 44 (3): 797–808. дои : 10.3233/jad-141704 . ПМК 4402159 . ПМИД 25362031 .
- ^ Маэзава И., Нгуен Х.М., Ди Лусенте Дж., Дженкинс Д.П., Сингх В., Хилт С., Ким К., Рангараджу С., Леви А.И., Вульф Х., Джин Л.В. (февраль 2018 г.). «Ингибирование Kv1.3 как потенциальная микроглиальная терапия болезни Альцгеймера: доклиническое доказательство концепции» . Мозг . 141 (2): 596–612. дои : 10.1093/brain/awx346 . ПМЦ 5837198 . ПМИД 29272333 .
- ^ Битон С., Вульф Х., Барбария Дж., Клот-Фейбесс О., Пеннингтон М., Бернард Д., Кахалан М.Д., Чанди К.Г., Беро Э. (ноябрь 2001 г.). «Селективная блокада K(+)-каналов Т-лимфоцитов улучшает экспериментальный аутоиммунный энцефаломиелит, модель рассеянного склероза» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (24): 13942–7. Бибкод : 2001PNAS...9813942B . дои : 10.1073/pnas.241497298 . ПМК 61146 . ПМИД 11717451 .
- ^ Перейти обратно: а б с д Цанг С.В., Нгуен CQ, Холл Д.Х., Чоу К.Л. (декабрь 2007 г.). «mab-7 кодирует новый трансмембранный белок, который управляет морфогенезом сенсорных лучей у C. elegans» (PDF) . Биология развития . 312 (1): 353–66. дои : 10.1016/j.ydbio.2007.09.037 . ПМИД 17959165 .
- ^ Перейти обратно: а б Харрис Р.А. (январь 1984 г.). «Дифференциальное влияние мембранных возмущающих веществ на потенциал-активируемые натриевые и кальциевые каналы и кальций-зависимые калиевые каналы» . Биофизический журнал . 45 (1): 132–4. Бибкод : 1984BpJ....45..132H . дои : 10.1016/s0006-3495(84)84137-5 . ПМЦ 1435273 . ПМИД 19431539 .
- ^ Таннер М.Р., Тайхья Р.Б., Хук Р., Германн Э.Дж., Родарте К.Е., Атик М.А., Нортон Р.С., Пеннингтон М.В., Битон С. (июль 2017 г.). «Пролонгированная иммуномодуляция при воспалительном артрите с использованием селективного блокатора каналов Kv1.3 HsTX1 [R14A] и его ПЭГилированного аналога» . Клиническая иммунология . 180 : 45–57. дои : 10.1016/j.clim.2017.03.014 . ПМК 5484050 . ПМИД 28389388 .
- ^ Перейти обратно: а б с д Олсен С., Люстиг К., Иадонато С., Тарча Е., Гуттман-Ясский Е. (2017). «695 Местное применение KPI-150, мощного ингибитора каналов Kv1.3 на эффекторных Т-клетках памяти, демонстрирует доклиническую эффективность на модели атопического дерматита» . Журнал исследовательской дерматологии . 137 (5): С119. дои : 10.1016/j.jid.2017.02.718 .
- ^ Эвальд Д.А., Нода С., Олива М., Литман Т., Накадзима С., Ли Х, Сюй Х., Уоркман К.Т., Шайперс П., Свитачева Н., Лабуда Т., Крюгер Дж.Г., Суарес-Фариньяс М., Кабашима К., Гуттман-Ясский Е (февраль). 2017). «Основные различия между атопическим дерматитом человека и мышиными моделями, определенные с помощью глобального транскриптомного профилирования» . Журнал аллергии и клинической иммунологии . 139 (2): 562–571. дои : 10.1016/j.jaci.2016.08.029 . ПМИД 27702671 .
- ^ Мартель БК, Ловато П., Боймер В., Оливри Т. (сентябрь 2017 г.). «Трансляционные модели атопического дерматита на животных для доклинических исследований» . Йельский журнал биологии и медицины . 90 (3): 389–402. ПМК 5612183 . ПМИД 28955179 .
- ^ Перейти обратно: а б Азам П., Шанкаранараянан А., Гомерик Д., Гриффи С., Вульф Х. (июнь 2007 г.). «Нацеливание на эффекторные Т-клетки памяти с помощью небольшой молекулы блокатора Kv1.3 PAP-1 подавляет аллергический контактный дерматит» . Журнал исследовательской дерматологии . 127 (6): 1419–29. дои : 10.1038/sj.jid.5700717 . ЧВК 1929164 . ПМИД 17273162 .
- ^ Перейти обратно: а б Уэяма А., Имура К., Касаи-Ямамото Э., Тай Н., Нагира М., Ситидзё М., Ясуи К. (декабрь 2013 г.). «Блокаторы Kv1.3 облегчают аллергический контактный дерматит путем преимущественного подавления эффекторных Т-клеток памяти на модели крыс». Клиническая и экспериментальная дерматология . 38 (8): 897–903. дои : 10.1111/ced.12097 . ПМИД 24252082 .
- ^ Ку Г.К., Блейк Дж.Т., Шах К., Старух М.Дж., Дюмон Ф., Вундерлер Д., Санчес М., Макманус О.Б., Сиротина-Мейшер А., Фишер П., Больц Р.К., Гетц М.А., Бейкер Р., Бао Дж., Кайзер Ф., Рупрехт К.М., Парсонс В.Х., Тонг Х.К., Ита И.Е., Пивничный Дж., Винсент С., Каннингем П., Хора Д., Фини В., Качоровски Г. (ноябрь 1999 г.). «Корреолид и его производные представляют собой новые иммунодепрессанты, блокирующие калиевые каналы Kv1.3 лимфоцитов». Клеточная иммунология . 197 (2): 99–107. дои : 10.1006/cimm.1999.1569 . ПМИД 10607427 .
- ^ Хаббол А.В., Ланг Б., Соуза М.А., Карран О.Д., Мартин Дж.Э., Ноулз CH (август 2012 г.). «Аутоантитела к вольт-зависимому калиевому каналу (K(v) 1) у пациентов с нарушением моторики кишечника и распределение каналов K(v) 1 в энтеральной нервно-мышечной системе человека (аутоантитела при нарушении моторики желудочно-кишечного тракта)». Нейрогастроэнтерология и моторика . 24 (8): 719–28, e344. дои : 10.1111/j.1365-2982.2012.01924.x . ПМИД 22591165 . S2CID 24100663 .
- ^ Битон С., Барбария Дж., Жиро П., Дево Дж., Бенольель А.М., Гола М., Сабатье Ж.М., Бернар Д., Крест М., Беро Э. (январь 2001 г.). «Селективная блокировка потенциалзависимых K+-каналов улучшает экспериментальный аутоиммунный энцефаломиелит и подавляет активацию Т-клеток» . Журнал иммунологии . 166 (2): 936–44. дои : 10.4049/jimmunol.166.2.936 . ПМИД 11145670 .
- ^ Варга З., Гуррола-Брионес Г., Папп Ф., Родригес де ла Вега Р.К., Педраса-Альва Г., Тайхья Р.Б., Гаспар Р., Карденас Л., Розенштейн Ю., Битон С., Поссани Л.Д., Паньи Г. (сентябрь 2012 г.). «Vm24, природный иммуносупрессивный пептид, мощно и избирательно блокирует калиевые каналы Kv1.3 Т-клеток человека» . Молекулярная фармакология . 82 (3): 372–82. дои : 10.1124/моль.112.078006 . ПМК 3422703 . ПМИД 22622363 .
- ^ Кунду-Райчаудхури С., Чен Ю.Дж., Вульф Х., Райчаудхури С.П. (декабрь 2014 г.). «Kv1.3 при псориатическом заболевании: PAP-1, низкомолекулярный ингибитор Kv1.3, эффективен при псориазе у мышей SCID - модель ксенотрансплантата» . Журнал аутоиммунитета . 55 : 63–72. дои : 10.1016/j.jaut.2014.07.003 . ПМК 4398058 . ПМИД 25175978 .
- ^ Гилхар А., Бергман Р., Эссей Б., Ульманн Ю., Эциони А. (январь 2011 г.). «Положительный эффект блокирования Kv1.3 на модели псориазоформных мышей SCID» . Журнал исследовательской дерматологии . 131 (1): 118–24. дои : 10.1038/jid.2010.245 . ПМИД 20739949 .
- ^ Перейти обратно: а б с д и Упадхьяй С.К., Экель-Махан К.Л., Мирболуки М.Р., Чонг И, Гриффи С.М., Шмунк Г., Кене А., Халбут Б., Иадонато С., Педерсен Б., Боррелли Е., Ван П.Х., Мукерджи Дж., Сассон-Корси П., Чанди К.Г. (июнь) 2013). «Селективный блокатор каналов Kv1.3 как средство лечения ожирения и резистентности к инсулину» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (24): E2239–48. Бибкод : 2013PNAS..110E2239U . дои : 10.1073/pnas.1221206110 . ПМЦ 3683782 . ПМИД 23729813 .
- ^ Такер К., Овертон Дж. М., Фадул Д. А. (август 2008 г.). «Направленная на ген Kv1.3 делеция изменяет продолжительность жизни и уменьшает ожирение за счет увеличения локомоции и метаболизма у мышей с нулевым рецептором меланокортина-4» . Международный журнал ожирения . 32 (8): 1222–32. дои : 10.1038/ijo.2008.77 . ПМЦ 2737548 . ПМИД 18542083 .
- ^ Сюй Дж., Кони П.А., Ван П., Ли Г., Качмарек Л., Ву Ю., Ли Ю., Флавелл Р.А., Дезир Г.В. (март 2003 г.). «Потенциал-управляемый калиевый канал Kv1.3 регулирует энергетический гомеостаз и массу тела» . Молекулярная генетика человека . 12 (5): 551–9. дои : 10.1093/hmg/ddg049 . ПМИД 12588802 .
- ^ Сюй Дж, Ван П., Ли Ю, Ли Г, Качмарек Л.К., Ву Ю, Кони П.А., Флавелл Р.А., Дезир Г.В. (март 2004 г.). «Потенциал-зависимый калиевый канал Kv1.3 регулирует периферическую чувствительность к инсулину» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (9): 3112–7. Бибкод : 2004PNAS..101.3112X . дои : 10.1073/pnas.0308450100 . ПМЦ 365752 . ПМИД 14981264 .
- ^ Перейти обратно: а б с д и ж Людино М.И., Берч А.М., Танака Б.С., Соколов Ю., Голдин А.Л., Чанди К.Г., Холл Дж.Э., Алкире М.Т. (октябрь 2013 г.). «Калиевые каналы, связанные с шейкером, в центральном медиальном ядре таламуса являются важными молекулярными мишенями для подавления возбуждения с помощью летучих общих анестетиков» . Журнал неврологии . 33 (41): 16310–22. doi : 10.1523/jneurosci.0344-13.2013 . ПМЦ 3792466 . ПМИД 24107962 .
- ^ Пэн Ю, Лу К, Ли З, Чжао Ю, Ван Ю, Ху Б, Сюй П, Ши Х, Чжоу Б, Пеннингтон М, Чанди К.Г., Тан Ю (апрель 2014 г.). «Блокада каналов Kv1.3 облегчает радиационное повреждение головного мозга» . Нейроонкология . 16 (4): 528–39. дои : 10.1093/neuonc/not221 . ПМЦ 3956348 . ПМИД 24305723 .
- ^ Перейти обратно: а б Ян Л., Фей К., Чжан Дж., Декстер С., член парламента Сарраса (январь 2000 г.). «Идентификация и характеристика гидра-металлопротеиназы 2 (HMP2): меприноподобной астациновой металлопротеиназы, которая участвует в морфогенезе стопы» . Разработка . 127 (1): 129–41. дои : 10.1242/dev.127.1.129 . ПМИД 10654607 .
- ^ Перейти обратно: а б Го М, Тэн М, Ню Л, Лю Ц, Хуан Ц, Хао Ц (апрель 2005 г.). «Кристаллическая структура богатого цистеином секреторного белка stecrisp показывает, что богатый цистеином домен имеет складку, подобную ингибитору K+-канала» . Журнал биологической химии . 280 (13): 12405–12. дои : 10.1074/jbc.M413566200 . ПМИД 15596436 .
- ^ Перейти обратно: а б Копперс А.Дж., Редди Т., О'Брайан М.К. (январь 2011 г.). «Роль секреторных белков, богатых цистеином, в мужской фертильности» . Азиатский журнал андрологии . 13 (1): 111–7. дои : 10.1038/aja.2010.77 . ПМЦ 3739402 . ПМИД 20972450 .
- ^ Ямазаки Ю., Морита Т. (сентябрь 2004 г.). «Структура и функция секреторных белков змеиного яда, богатых цистеином». Токсикон . 44 (3): 227–31. doi : 10.1016/j.токсикон.2004.05.023 . ПМИД 15302528 .