PA клан протеаз
| PA клан протеаз | |
|---|---|
 | |
| Идентификаторы | |
| Символ | Н/Д |
| Пфам Клан | CL0124 |
| ИнтерПро | ИПР009003 |
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 50494 / SCOPe / СУПФАМ |
| Мембраном | 319 |
Клан PA ( протеазы ) представляет собой смешанных нуклеофилов, суперсемейство А крупнейшую группу протеаз с общим происхождением, определяемым по структурной гомологии . Члены имеют химотрипсиноподобную складку и схожие механизмы протеолиза , но могут иметь идентичность <10%. Клан содержит как цистеиновые , так и сериновые протеазы (разные нуклеофилы ). [ 1 ] [ 2 ] Протеазы клана PA можно обнаружить в растениях . [ 3 ] животные , [ 3 ] грибы , [ 3 ] эубактерии , [ 4 ] архея [ 5 ] [ 6 ] и вирусы . [ 2 ]
Общее использование каталитической триады для гидролиза несколькими кланами протеаз, включая клан PA, представляет собой пример конвергентной эволюции . [ 7 ] Различия в каталитической триаде внутри клана ПА также являются примером дивергентной эволюции активных центров ферментов. [ 2 ]
История
[ редактировать ]В 1960-х годах сходство последовательностей нескольких протеаз указывало на их эволюционное родство. [ 8 ] Они были сгруппированы в химотрипсиноподобные сериновые протеазы. [ 9 ] (сейчас называется семейством S1 ). Когда структуры этих и других протеаз были определены с помощью рентгеновской кристаллографии в 1970-х и 80-х годах, было замечено, что несколько вирусных протеаз, таких как протеаза вируса травления табака, продемонстрировали структурную гомологию, несмотря на отсутствие заметного сходства последовательностей и даже другой нуклеофил. [ 2 ] [ 10 ] [ 11 ] На основе структурной гомологии было выделено надсемейство , позднее названное кланом ПА (по системе классификации MEROPS ). По мере того, как решено больше структур, к суперсемейству клана PA добавляется больше семейств протеаз. [ 12 ] [ 13 ]
Этимология
[ редактировать ]Буква P относится к пропротеазам смешанного нуклеофила. Буква A указывает на то, что это был первый такой клан, который был идентифицирован (существуют также кланы PB, PC, PD и PE). [ 1 ]
Структура
[ редактировать ]

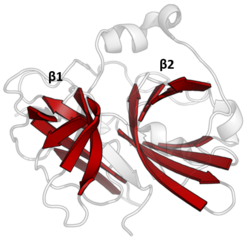
Несмотря на сохранение всего лишь 10% идентичности последовательностей, члены клана PA, выделенные из вирусов, прокариот и эукариот, демонстрируют структурную гомологию и могут быть выровнены по структурному сходству (например, с помощью DALI ). [ 3 ]
Двойная β-стволка
[ редактировать ]Все протеазы клана PA имеют общий основной мотив из двух β-цилиндров с ковалентным катализом, осуществляемым мотивом каталитической триады кислота-гистидин-нуклеофил . Стволы расположены перпендикулярно друг другу, а гидрофобные остатки удерживают их вместе в качестве основного каркаса для фермента. Остатки триады распределяются между двумя цилиндрами, так что катализ происходит на их границе. [ 14 ]
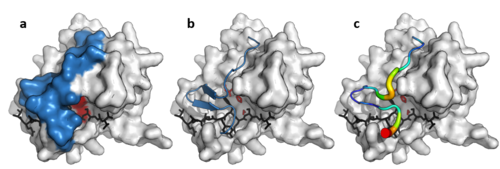
Вирусная протеазная петля
[ редактировать ]В дополнение к двойному β-цилиндрическому ядру некоторые вирусные протеазы (например, протеаза TEV ) имеют длинную гибкую С-концевую петлю, которая образует крышку, полностью закрывающую субстрат и создающую туннель связывания. Этот туннель содержит набор карманов прочного связывания, так что каждая боковая цепь пептида-субстрата (от P6 до P1') связана в комплементарном сайте (от S6 до S1'), а специфичность обеспечивается большой площадью контакта между ферментом и субстратом. [ 11 ] И наоборот, клеточные протеазы, у которых нет этой петли, такие как трипсин, обладают более широкой специфичностью .
Эволюция и функции
[ редактировать ]Каталитическая активность
[ редактировать ]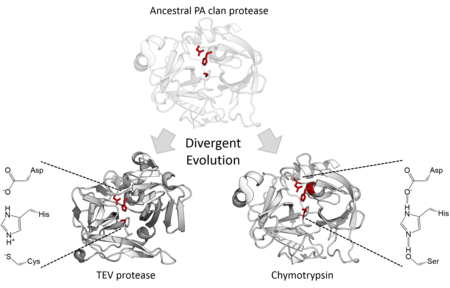
Структурная гомология указывает на то, что члены клана ПА происходят от общего предка одной и той же складки. Хотя протеазы клана PA используют каталитическую триаду, осуществляющую двухэтапный нуклеофильный катализ . [ 7 ] некоторые семьи используют серин в качестве нуклеофила, тогда как другие используют цистеин . [ 2 ] Таким образом, суперсемейство является крайним примером дивергентной эволюции ферментов , поскольку в ходе эволюционной истории основной каталитический остаток фермента менялся в разных семействах. [ 15 ] Было показано , что помимо их структурного сходства направленная эволюция способна превращать цистеиновую протеазу в активную сериновую протеазу. [ 16 ] Все клеточные протеазы клана PA являются сериновыми протеазами , однако существуют как сериновых, так и цистеиновых протеаз . семейства вирусных протеаз [ 7 ] Большинство из них представляют собой эндопептидазы , за исключением семейства экзопептидаз S46 . [ 17 ] [ 18 ]
Биологическая роль и субстратная специфичность
[ редактировать ]Помимо различий в их основных каталитических механизмах, протеазы клана PA также демонстрируют широкую дивергентную эволюцию в функциях. Члены клана PA встречаются у эукариот , прокариотов и вирусов и выполняют широкий спектр функций. У млекопитающих некоторые из них участвуют в свертывании крови (например, тромбин ) и поэтому обладают высокой субстратной специфичностью, а также пищеварением (например, трипсин ) с широкой субстратной специфичностью. Некоторые змеиные яды также являются протеазами клана PA, такими как гадюки гемотоксин , и мешают каскаду свертывания крови жертвы. Кроме того, такие бактерии, как золотистый стафилококк, выделяют эксфолиативный токсин , который переваривает и повреждает ткани хозяина. Многие вирусы экспрессируют свой геном в виде одного массивного полипротеина и используют протеазу клана PA для расщепления его на функциональные единицы (например, полиомиелита , норовируса и протеазы TEV ). [ 19 ] [ 20 ]
В суперсемействе также есть несколько псевдоферментов , остатки каталитической триады которых мутированы и поэтому функционируют как связывающие белки. [ 21 ] Например, гепаринсвязывающий белок азуроцидин содержит глицин вместо нуклеофила и серин вместо гистидина. [ 22 ]
Семьи
[ редактировать ]Внутри клана ПА (P=протеазы смешанных нуклеофилов ) семейства обозначаются каталитическим нуклеофилом (C= цистеиновые протеазы , S= сериновые протеазы ). Несмотря на отсутствие гомологии последовательностей для клана ПА в целом, отдельные семейства внутри него можно идентифицировать по сходству последовательностей.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Роулингс Н.Д., Барретт А.Дж., Бейтман А. (январь 2012 г.). «MEROPS: база данных протеолитических ферментов, их субстратов и ингибиторов» . Исследования нуклеиновых кислот . 40 (Проблема с базой данных): D343-50. дои : 10.1093/nar/gkr987 . ПМК 3245014 . ПМИД 22086950 .
- ^ Jump up to: а б с д и Базан Дж. Ф., Флеттерик Р. Дж. (ноябрь 1988 г.). «Вирусные цистеиновые протеазы гомологичны трипсиноподобному семейству сериновых протеаз: структурные и функциональные значения» . Труды Национальной академии наук Соединенных Штатов Америки . 85 (21): 7872–6. Бибкод : 1988PNAS...85.7872B . дои : 10.1073/pnas.85.21.7872 . ПМЦ 282299 . ПМИД 3186696 .
- ^ Jump up to: а б с д Ласкар А., Роджер Э.Дж., Чаттерджи А., Мандал С. (май 2012 г.). «Моделирование и структурный анализ сериновых протеаз клана PA» . Исследовательские заметки BMC . 5 : 256. дои : 10.1186/1756-0500-5-256 . ПМЦ 3434108 . ПМИД 22624962 .
- ^ Барбоса Дж.А., Салданья Дж.В., Гарратт Р.С. (июль 1996 г.). «Новые особенности активных центров и карманов специфичности сериновой протеазы: анализ последовательностей и моделирование исследований глутамат-специфических эндопептидаз и эпидермолитических токсинов» . Белковая инженерия . 9 (7): 591–601. дои : 10.1093/белок/9.7.591 . ПМИД 8844831 .
- ^ «МЕРОПС - архейные протеазы S01» .
- ^ Руис-Перес Ф., Натаро Дж.П. (март 2014 г.). «Бактериальные сериновые протеазы, секретируемые аутотранспортным путем: классификация, специфичность и роль в вирулентности» . Клеточные и молекулярные науки о жизни . 71 (5): 745–70. дои : 10.1007/s00018-013-1355-8 . ПМК 3871983 . ПМИД 23689588 .
- ^ Jump up to: а б с Буллер А.Р., Таунсенд, Калифорния (февраль 2013 г.). «Внутренние эволюционные ограничения на структуру протеазы, ацилирование ферментов и идентичность каталитической триады» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (8): Е653-61. Бибкод : 2013PNAS..110E.653B . дои : 10.1073/pnas.1221050110 . ПМК 3581919 . ПМИД 23382230 .
- ^ де Хаэн С., Нейрат Х., Теллер округ Колумбия (февраль 1975 г.). «Филогения родственных трипсину сериновых протеаз и их зимогенов. Новые методы исследования отдаленных эволюционных связей». Журнал молекулярной биологии . 92 (2): 225–59. дои : 10.1016/0022-2836(75)90225-9 . ПМИД 1142424 .
- ^ Леск А.М., Фордхэм У.Д. (май 1996 г.). «Консервативность и изменчивость структур сериновых протеиназ семейства химотрипсинов». Журнал молекулярной биологии . 258 (3): 501–37. дои : 10.1006/jmbi.1996.0264 . ПМИД 8642605 .
- ^ Горбаленя А.Е., Блинов В.М., Донченко А.П. (январь 1986 г.). «Протеиназа 3C, кодируемая полиовирусом: возможная эволюционная связь между семействами клеточных сериновых и цистеиновых протеиназ» . Письма ФЭБС . 194 (2): 253–7. дои : 10.1016/0014-5793(86)80095-3 . ПМИД 3000829 . S2CID 23268152 .
- ^ Jump up to: а б Фан Дж, Зданов А, Евдокимов А.Г., Тропеа Дж.Э., Петерс Х.К., Капуст Р.Б., Ли М., Влодавер А., Во Д.С. (декабрь 2002 г.). «Структурная основа субстратной специфичности протеазы вируса травления табака» . Журнал биологической химии . 277 (52): 50564–72. дои : 10.1074/jbc.M207224200 . ПМИД 12377789 .
- ^ Аллер М., Черная М.М., Малкольм Б.А., Джеймс М.Н. (май 1994 г.). «Пикорнавирусные цистеиновые протеиназы 3C имеют складку, аналогичную химотрипсиноподобным сериновым протеиназам». Природа . 369 (6475): 72–6. Бибкод : 1994Natur.369...72A . дои : 10.1038/369072a0 . ПМИД 8164744 . S2CID 4312593 .
- ^ Снейдер Э.Дж., Вассенаар А.Л., ван Динтен Л.К., Спаан В.Дж., Горбаленя А.Е. (март 1996 г.). «Протеаза nsp4 артеривируса является прототипом новой группы химотрипсиноподобных ферментов, 3C-подобных сериновых протеаз» . Журнал биологической химии . 271 (9): 4864–71. дои : 10.1074/jbc.271.9.4864 . ПМИД 8617757 .
- ^ Догерти В.Г., Паркс Т.Д., Кэри С.М., Базан Дж.Ф., Флеттерик Р.Дж. (сентябрь 1989 г.). «Характеристика каталитических остатков протеиназы 49-кДа вируса травления табака». Вирусология . 172 (1): 302–10. дои : 10.1016/0042-6822(89)90132-3 . ПМИД 2475971 .
- ^ Ласкар А., Роджер Э.Дж., Чаттерджи А., Мандал С. (май 2012 г.). «Моделирование и структурный анализ сериновых протеаз клана PA» . Исследовательские заметки BMC . 5 (1): 256. дои : 10.1186/1756-0500-5-256 . ПМЦ 3434108 . ПМИД 22624962 .
- ^ Шафи Т., Гатти-Лафранкони П., Минтер Р., Холлфельдер Ф. (сентябрь 2015 г.). «Эволюция восстановления инвалидности ведет к химически универсальной протеазе, допускающей нуклеофилы» . ХимБиоХим . 16 (13): 1866–1869. дои : 10.1002/cbic.201500295 . ПМЦ 4576821 . ПМИД 26097079 .
- ^ Сузуки Ю, Сакамото Ю, Танака Н, Окада Х, Морикава Ю, Огасавара В (март 2014 г.). «Идентификация каталитической триады экзопептидаз семейства S46, тесно связанных с эндопептидазами клана PA» . Научные отчеты . 4 : 4292. Бибкод : 2014NatSR...4E4292S . дои : 10.1038/srep04292 . ПМЦ 3944710 . ПМИД 24598890 .
- ^ Сакамото Ю, Сузуки Ю, Иидзука И, Татеока С, Роппонги С, Фудзимото М, Инака К, Танака Х, Масаки М, Охта К, Окада Х, Нонака Т, Морикава Ю, Накамура К. Т., Огасавара В, Танака Н ( май 2014). «Пептидазы S46 являются первыми экзопептидазами, входящими в клан PA» . Научные отчеты 4 : 4977. Бибкод : 2014NatSR...4E4977S . дои : 10.1038/srep04977 . ПМК 4021333 . ПМИД 24827749 .
- ^ Салвесен Г (2013). Роулингс Н. (ред.). Справочник по протеолитическим ферментам . Бостон: Академическая пресса. ISBN 9780123822192 .
- ^ Полгар Л. (октябрь 2005 г.). «Каталитическая триада сериновых пептидаз» . Клеточные и молекулярные науки о жизни . 62 (19–20): 2161–72. дои : 10.1007/s00018-005-5160-x . ПМЦ 11139141 . ПМИД 16003488 . S2CID 3343824 .
- ^ Тодд А.Е., Оренго, Калифорния, Торнтон Дж.М. (октябрь 2002 г.). «Последовательность и структурные различия между ферментными и неферментными гомологами» . Структура . 10 (10): 1435–51. дои : 10.1016/s0969-2126(02)00861-4 . ПМИД 12377129 .
- ^ Иверсен Л.Ф., Каструп Дж.С., Бьёрн С.Е., Расмуссен П.Б., Виберг ФК, Флодгаард Х.Дж., Ларсен И.К. (апрель 1997 г.). «Структура HBP, многофункционального белка со складкой сериновой протеиназы». Структурная биология природы . 4 (4): 265–8. дои : 10.1038/nsb0497-265 . ПМИД 9095193 . S2CID 19949043 .
Внешние ссылки
[ редактировать ]- MEROPS - Полная база данных протеаз
- Суперсемейство. Архивировано 24 июня 2016 г. в Wayback Machine — база данных белковых складок.