Гигантопитек
| Гигантопитек Временной диапазон: ранний – средний плейстоцен.
~ | |
|---|---|

| |
| Реконструированная гигантопитека нижняя челюсть в Кливлендском музее естественной истории , Огайо. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Симииформы |
| Семья: | Гоминиды |
| Племя: | † Сивапитецины |
| Род: | † Гигантопитек |
| Разновидность: | † Г. Блэки
|
| Биномиальное имя | |
| † Гигантопитек черный фон Кенигсвальд , 1935 год. [ 1 ]
| |
Гигантопитек ( / d ʒ aɪ ˌ ɡ æ n t oʊ p ɪ ˈ θ i k ə s , ˈ p ɪ θ ɪ k ə s , d ʒ ɪ - / jahy-gan-toh-pi-thee-kuhs, pith-i -кух, джи- ; [ 2 ] горит. ' гигантская обезьяна ' ) — вымерший род обезьян , обитавший на юге Китая от 2 миллионов до примерно 300 000—200 000 лет назад в период раннего и среднего плейстоцена , представленный одним видом, Gigantopithecus blacki . [ 3 ] [ 4 ] Потенциальные идентификации также были сделаны в Таиланде , Вьетнаме и Индонезии . Первые останки гигантопитека , два третьих коренных зуба , были обнаружены в аптеке антропологом Ральфом фон Кенигсвальдом в 1935 году, который впоследствии описал обезьяну. была найдена первая нижняя челюсть и более 1000 зубов В 1956 году в Лючэне , а с тех пор еще множество останков было обнаружено как минимум в 16 местах. В настоящее время известны только зубы и четыре челюсти, а другие элементы скелета, вероятно, были съедены дикобразами до того, как они смогли окаменеть. [ 5 ] Гигантопитек когда-то считался гоминином , представителем человеческой линии, но теперь считается, что он тесно связан с орангутанами , отнесенными к подсемейству Ponginae .
Гигантопитек традиционно восстанавливался как массивная гориллоподобная обезьяна, потенциально весившая при жизни 200–300 кг (440–660 фунтов), но из-за нехватки останков оценки общего размера весьма спекулятивны. Этот вид мог иметь половой диморфизм : самцы были намного крупнее самок. Резцы редуцированы, а клыки функционируют как щечные зубы ( премоляры и коренные зубы). Премоляры имеют высокую коронку , четвертый премоляр очень похож на моляр. Коренные зубы являются самыми большими из всех известных обезьян и имеют относительно плоскую поверхность. У гигантопитеков была самая толстая эмаль по абсолютным меркам среди всех обезьян, в некоторых местах до 6 мм (четверть дюйма), хотя это достаточно толстая лишь с учетом размера зубов.
Гигантопитек, по-видимому, был универсальным травоядным, питающимся лесными растениями C 3 , с челюстями, приспособленными для измельчения, дробления и разрезания жестких волокнистых растений, а толстая эмаль защищала от продуктов с абразивными частицами, такими как стебли, корни и клубни. с грязью. На некоторых зубах имеются следы плодов семейства инжира , которые, возможно, были важными диетическими компонентами. В основном он обитал в субтропических и тропических лесах и вымер около 300 000 лет назад, вероятно, из-за сокращения предпочтительной среды обитания из-за изменения климата и потенциально архаичной деятельности человека. Гигантопитек стал популярен в кругах криптозоологов как тибетский йети или американский снежный человек , обезьяноподобные существа в местном фольклоре.
Открытие
[ редактировать ]История исследований
[ редактировать ]
Gigantopithecus blacki был назван антропологом Ральфом фон Кенигсвальдом в 1935 году на основании двух третьих нижних коренных зубов , которые, как он отмечал, имели огромные размеры (первый был « Ein gewaltig Grosser (...) Molar », второй описывался как « der enorme Grösse besitzt "), размером 20 мм × 22 мм (0,79 дюйма × 0,87 дюйма). [ 1 ] Видовое название blacki дано в честь канадского палеоантрополога Дэвидсона Блэка , который изучал эволюцию человека в Китае и умер в прошлом году. Фон Кенигсвальд, работавший в Минералогической службе Голландской Ост-Индии на Яве, нашел зубы в аптеке в Гонконге , где они продавались как « кости дракона » для использования в традиционной китайской медицине . К 1939 году, купив еще несколько зубов, он определил, что они возникли где-то в Гуандуне или Гуанси . Он не мог официально описать типовой образец до 1952 года из-за его интернирования японскими войсками во время Второй мировой войны . Первоначально обнаруженные зубы являются частью коллекции Утрехтского университета . [ 5 ] [ 6 ]
исследовательской группе, которую возглавил китайский палеонтолог Пей Вэньчжун, (IVPP) поручил В 1955 году Китайский институт палеонтологии и палеоантропологии позвоночных найти первоначальное местонахождение гигантопитеков . Они собрали 47 зубов среди партий «костей дракона» в Гуандуне и Гуанси. В 1956 году команда обнаружила первые останки in situ , третий моляр и премоляр , в пещере (впоследствии названной « Пещера гигантопитеков ») на горе Нюсуй в Гуанси. Также в 1956 году фермер Лючэн еще больше зубов и первую нижнюю челюсть Сюхуай Цинь обнаружил на своем поле . С 1957 по 1963 год исследовательская группа ИВПП проводила раскопки на этом участке и обнаружила еще две челюсти и более 1000 зубов. [ 5 ] [ 7 ] [ 8 ] [ 6 ]
Подтвержденные останки гигантопитека с тех пор были обнаружены в 16 различных местах на юге Китая. Самые северные места — Лунгупо и Лунгудун , расположенные к югу от реки Янцзы , а самые южные — на острове Хайнань в Южно-Китайском море . Изолированный клык из пещеры Тхум Хуен , Вьетнам, и четвертый премоляр из Фабонга , Таиланд, возможно, могли быть отнесены к гигантопитеку , хотя они также могли представлять собой вымершего орангутанга Понго вейденрейчи . [ 5 ] Два фрагмента нижней челюсти, каждый из которых сохраняет два последних коренных зуба из Семоно в Центральной Яве , Индонезия, описанные в 2016 году, могут представлять собой гигантопитека . [ 9 ] Самые старые останки датируются 2 миллионами лет назад из пещеры Байконг , а самые молодые - 380 000–310 000 лет назад из пещеры Хэй . [ 5 ] В 2014 году четвертая подтвержденная нижняя челюсть была обнаружена в Яньляне , Центральный Китай. [ 10 ] Зубы , о которых свидетельствуют обширные грызунов следы , скопились в пещерах, вероятно, из-за деятельности дикобразов . Дикобразы грызут кости, чтобы получить питательные вещества, необходимые для роста игл, а также могут затаскивать большие кости в свои подземные норы и поглощать их целиком, за исключением твердых, покрытых эмалью коронок зубов. Это может объяснить, почему зубы обычно находят в большом количестве и почему останки, кроме зубов, встречаются так редко. [ 5 ]
Классификация
[ редактировать ]Г. Блэки
[ редактировать ]
В 1935 году фон Кенигсвальд считал гигантопитеков близким родственником позднемиоценовых сивапитеков из Индии. [ 1 ] В 1939 году южноафриканский палеонтолог Роберт Брум выдвинул гипотезу, что он был тесно связан с австралопитеком и последним общим предком человека и других обезьян. [ 11 ] В 1946 году немецкий еврейский антрополог Франц Вайденрайх описал гигантопитека как предка человека как « гигантантропа », полагая, что человеческое происхождение прошло через гигантскую фазу. Он заявил, что зубы более похожи на зубы современного человека и человека прямоходящего (в то время « питекантропа » для ранних яванских экземпляров ), и предположил, что зубы происходят от гигантопитека к яванской обезьяне -мегантропу (тогда считавшейся предком человека) и « Питекантроп », « Явантроп » и, наконец, австралийские аборигены . Это было частью его межрегиональной гипотезы , согласно которой все современные расы и этнические группы развились независимо от местного архаичного человеческого вида, а не имели общего более недавнего и полностью современного общего предка. [ 12 ] В 1952 году фон Кенигсвальд согласился с тем, что гигантопитек был человеком , но считал, что это ответвление, а не предок человека. [ 13 ] В течение следующих трех десятилетий последовало много споров о том, был ли гигантопитек гоминином или нет, пока гипотеза «из Африки» не опровергла гипотезу «из Азии» и мультирегиональную гипотезу, твердо установив происхождение человечества в Африке. [ 5 ] [ 6 ]
Gigantopithecus теперь классифицируется в подсемействе Ponginae , тесно связанном с Sivapithecus и Indopithecus . Это сделало бы его ближайшими ныне живущими родственниками орангутанов . Тем не менее, существует несколько схожих черт ( синапоморфий ), связывающих гигантопитеков и орангутанов из-за фрагментарных останков, при этом основным морфологическим аргументом является его близкое сходство с сивапитеком , которого лучше определить как понгина на основании особенностей черепа. В 2017 году китайский палеоантрополог Инци Чжан и американский антрополог Терри Харрисон предположили, что гигантопитек наиболее тесно связан с китайским люфэнпитеком , который вымер за 4 миллиона лет до гигантопитека . [ 5 ]
В 2019 году пептидное секвенирование белков дентина и коренного зуба эмали гигантопитека из пещеры Чуйфэн показало, что гигантопитеки действительно были тесно связаны с орангутанами и, если предположить, что текущая скорость мутаций у орангутанов остается постоянной, имели общего предка примерно 12–10 миллионов лет назад. назад, в среднем и позднем миоцене. Их последний общий предок мог быть частью миоценовой радиации обезьян. В том же исследовании было рассчитано время расхождения между Ponginae и африканскими человекообразными обезьянами около 26–17,7 миллионов лет назад. [ 14 ]
Кладограмма по Чжану и Харрисону, 2017 г.: [ 5 ]
| Гоминоид (обезьяны) | |





" Г. биласпуренсис "
[ редактировать ]нижнюю челюсть возрастом 8,6 миллиона лет с холмов Сивалик классифицировали на севере Индии как « G. bilaspurensis В 1969 году палеонтологи Элвин Л. Саймонс и Шив Радж Кумар Чопра » , которые полагали, что она была предком G. blacki . [ 5 ] [ 6 ] Этот зуб напоминал коренной зуб, обнаруженный в 1915 году на пакистанском плато Потохар , который тогда классифицировали как « Dryopithecus giganteus ». Фон Кенигсвальд реклассифицировал « D. giganteus » в 1950 году в отдельный род Indopithecus снова изменили его на « G. giganteus ». , но в 1979 году американские антропологи Фредерик Салай и Эрик Дельсон [ 15 ] пока Индопитек не был воскрешен в 2003 году австралийским антропологом Дэвидом Кэмероном . [ 16 ] [ 5 ] « G. bilaspurensis » теперь считается синонимом Indopithecus giganteus , оставляя Gigantopithecus монотипическим (только с одним видом), G. blacki . [ 17 ] [ 5 ]
Описание
[ редактировать ]
Размер
[ редактировать ]Оценки общего размера весьма спекулятивны, поскольку известны только элементы зубов и челюстей, а размер коренных зубов и общая масса тела не всегда коррелируют, как, например, в случае постсобачьих мегадонтных гоминидов (маленьких приматов с массивными коренными зубами и толстой эмалью). [ 18 ] В 1946 году Вайденрайх выдвинул гипотезу, что гигантопитеки вдвое крупнее самцов горилл. [ 12 ] В 1957 году Пей оценил общую высоту около 3,7 м (12 футов). В 1970 году Саймонс и американский палеонтолог Питер Эттель достигли высоты почти 2,7 м (9 футов) и веса до 270 кг (600 фунтов), что примерно на 40% тяжелее среднего самца гориллы. В 1979 году американский антрополог Альфред Э. Джонсон-младший использовал размеры горилл, чтобы оценить длину бедренной кости 54,4 см (1 фут 9 дюймов) и плечевой кости длину 62,7 см (2 фута 1 дюйм) для гигантопитеков , около 20–25%. длиннее, чем у горилл. [ 19 ] В 2017 году китайский палеоантрополог Инци Чжан и американский антрополог Терри Харрисон предположили, что масса тела составляет 200–300 кг (440–660 фунтов), хотя признали, что невозможно получить надежную оценку массы тела без более полных останков. [ 5 ]
Средняя максимальная длина верхних клыков у предполагаемых мужчин и женщин составляет 21,1 мм (0,83 дюйма) и 15,4 мм (0,61 дюйма) соответственно, а нижняя челюсть III (предполагаемая мужская) на 40% больше, чем нижняя челюсть I (предполагаемая женская). Это подразумевает половой диморфизм : самцы крупнее самок. Столь высокая степень диморфизма уступает только гориллам среди современных обезьян по размеру собак, и никто не превосходит ее по несоответствию нижней челюсти. [ 5 ]
Зубы и челюсти
[ редактировать ]Как и другие обезьяны, гигантопитек имел зубную формулу 2.1.2.3 2.1.2.3 с двумя резцами , одним клыком, двумя премолярами и тремя коренными зубами в каждой половине челюсти обеих челюстей. [ 5 ] Было высказано предположение, что из-за отсутствия заточки фасеток (которые сохраняют их острыми) и их общей толщины клыки функционировали как премоляры и коренные зубы (щечные зубы). Как и у других обезьян с увеличенными коренными зубами, резцы у гигантопитека редуцированы. [ 20 ] [ 21 ] Ношение резцов на язычной стороне (язычной поверхности), которая может доходить до корня зуба , предполагает недостаточный прикус . [ 5 ] Общая анатомия нижней челюсти и стираемость зубов предполагают движение челюсти из стороны в сторону во время жевания (боковая экскурсия). [ 22 ] Резцы и клыки имеют чрезвычайно длинные корни зубов, по крайней мере вдвое длиннее коронки зуба (видимой части зуба). Эти зубы были плотно прижаты друг к другу. [ 5 ]
На верхней челюсти первый премоляр (P 3 ) составляет в среднем 20,3 мм × 15,2 мм (0,8 дюйма × 0,6 дюйма) по площади поверхности, второй премоляр (P 4 ) 15,2 мм × 16,4 мм (0,60 дюйма × 0,65 дюйма), первые и/или вторые моляры (M 1/2 , которые трудно различить) 19,8 мм × 17,5 мм (0,78 × 0,69 дюйма) и третий моляр (М 3 ) 20,3 × 17,3 мм (0,80 × 0,68 дюйма). В нижней челюсти P 3 составляет в среднем 15,1 × 20,3 мм (0,59 × 0,80 дюйма), P 4 13,7 × 20,3 мм (0,54 × 0,80 дюйма), M 1/2 18,1 × 20,8 мм (0,71 × 0,82 дюйма). дюймов) и M 3 16,9 мм × 19,6 мм (0,67 × 0,77 дюйма). Коренные зубы — самые большие из всех известных обезьян. [ 5 ] Зубы постоянно развивались, становясь все больше и больше. [ 23 ] Премоляры имеют высокую коронку, нижние имеют два корня зубов, а верхние — три. Нижние коренные зубы имеют низкую коронку, длинные и узкие, с талией по средней линии, которая более выражена у нижних коренных зубов, с низко расположенными выпуклыми бугорками и закругленными гребнями. [ 5 ]

Зубная эмаль на коренных зубах в абсолютном измерении самая толстая среди всех известных обезьян: в среднем 2,5–2,9 мм (0,098–0,114 дюйма) на трех разных коренных зубах и более 6 мм (0,24 дюйма) на бугорках со стороны языка (лингвальных). верхнего моляра. [ 22 ] Это вызвало сравнения с вымершими гомининами Paranthropus , которые имели чрезвычайно большие коренные зубы и толстую эмаль для своего размера. [ 22 ] [ 20 ] Однако, что касается размера зубов, толщина эмали гигантопитеков совпадает с толщиной эмали некоторых других ныне живущих и вымерших человекообразных обезьян. Подобно орангутанам и, возможно, всем понгинам (хотя и в отличие от африканских обезьян), коренной зуб гигантопитека имел большую и плоскую (табличную) шлифовальную поверхность с ровным эмалевым покрытием и короткие дентинные рога (участки слоя дентина, выступающие вверх в верхнюю часть эмали). слой). [ 18 ] Коренные зубы — самые гипсодонтные (эмаль которых выходит за пределы десен) у любой обезьяны. [ 5 ]
Палеобиология
[ редактировать ]Диета
[ редактировать ]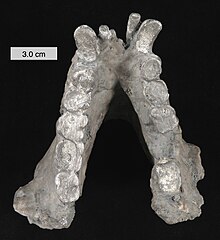
Масштаб 3 см (1,2 дюйма)
Гигантопитек считается травоядным . Анализ изотопа углерода-13 предполагает потребление C 3 растений , таких как фрукты, листья и другие лесные растения. [ 24 ] Крепкая нижняя челюсть гигантопитека указывает на то, что он был способен выдерживать большие нагрузки при пережевывании жесткой или твердой пищи. Однако та же анатомия нижней челюсти обычно наблюдается у современных обезьян, которые в основном едят мягкие листья ( folivores ) или семена ( granivores ). Зубы гигантопитеков имеют заметно более низкую скорость образования ямок (вызванных употреблением в пищу маленьких твердых предметов), чем у орангутанов, что больше похоже на уровень, наблюдаемый у шимпанзе , что может указывать на аналогичную универсальную диету . [ 5 ]
Премоляры, похожие на коренные, большие коренные зубы и щечные зубы с длинными корнями могут указывать на жевание, дробление и измельчение объемных и волокнистых материалов. [ 25 ] [ 26 ] Толстая эмаль предполагает наличие абразивных предметов, таких как частицы грязи с еды, собранной рядом или на земле (например, побеги бамбука ). [ 22 ] Точно так же анализ изотопов кислорода показывает, что гигантопитеки потребляли больше низколежащих растений, таких как стебли, корни и травы, чем орангутанги. Зубной камень указывает на употребление клубней . [ 27 ] Гигантопитеки , по-видимому, не потребляли обычные травы саванны ( растения C4 ). [ 24 ] Тем не менее, в 1990 году было установлено, что несколько фитолитов опала, прилипших к четырем зубам из пещеры Гигантопитек , произошли от трав; однако большинство фитолитов напоминают волоски плодов семейства инжирных , к которым относятся инжир , шелковица , хлебное дерево и баньян . Это говорит о том, что фрукты были важным компонентом питания, по крайней мере, для этой популяции гигантопитеков . [ 26 ]
возрастом 400 000–320 000 лет Зубы среднего плейстоцена из пещеры Хэцзян на юго-востоке Китая (недалеко от времени вымирания) демонстрируют некоторые отличия от материала раннего плейстоцена из других мест, что потенциально может указывать на то, что гигантопитеки Хэцзян были специализированной формой, адаптирующейся к изменяющейся окружающей среде. с различными пищевыми ресурсами. Зубы Хэцзян имеют менее ровную (более зубчатую) внешнюю поверхность эмали из-за наличия вторичных гребней, исходящих из паракона и протоконуса на стороне моляра ближе к средней линии (медиально), а также более острых основных гребней. То есть зубы не такие плоские. [ 10 ] [ 28 ] [ 29 ]
В 1957 году, основываясь на останках копытных животных в пещере, расположенной в, казалось бы, недоступной горе, Пей предположил, что гигантопитек был пещерным хищником, и принес этих животных внутрь. [ 30 ] Эта гипотеза больше не считается жизнеспособной, поскольку ее зубная анатомия соответствует травоядным. [ 24 ] В 1975 году американский палеоантрополог Тим Д. Уайт обнаружил сходство между челюстями и зубными рядами гигантопитека и гигантской панды и предположил, что они оба занимают ту же нишу , что и специалисты по бамбуку . [ 31 ] Это получило поддержку со стороны некоторых последующих исследователей, но более толстая эмаль и гипсодонтность у гигантопитеков могли указывать на различную функциональность этих зубов. [ 22 ]
Зависимость этого вида от коры и веток в качестве источника питания привела к его вымиранию. [ 32 ]
Рост
[ редактировать ], По оценкам, формирование постоянного третьего моляра Gigantopithecus исходя из примерно 600–800 дней, необходимых для формирования эмали на бугорках (что довольно долго), заняло четыре года, что находится в пределах диапазона (хотя и гораздо выше диапазон) того, что наблюдается у людей и шимпанзе. Как и у многих других ископаемых человекообразных обезьян, скорость образования эмали вблизи соединения эмали и дентина (дентин — это слой, заполненный нервами под эмалью), по оценкам, начинается примерно с четырех мкм в день; это наблюдается только у молочных зубов современных обезьян. [ 20 ]
Секвенирование белков эмали гигантопитека выявило альфа-2-HS-гликопротеин (AHSG), который у современных обезьян играет важную роль в минерализации костей и дентина. Поскольку AHSG был обнаружен в эмали, а не в дентине, он мог быть дополнительным компонентом у Gigantopithecus , который способствовал биоминерализации эмали во время длительного амелогенеза (роста эмали). [ 14 ]
Патология
[ редактировать ]Коренные зубы гигантопитека имеют высокую степень кариеса (11%), что может означать, что в его рацион обычно включались фрукты. [ 5 ] [ 26 ] На коренных зубах из пещеры Гигантопитеков часто наблюдается точечная гипоплазия эмали , при которой эмаль неправильно формируется с ямками и бороздками. Это может быть вызвано недоеданием в годы роста, что может указывать на периодическую нехватку продовольствия, хотя это также может быть вызвано другими факторами. [ 26 ] У образца PA1601-1 из пещеры Яньлян имеются признаки потери правого второго моляра до прорезывания соседнего третьего моляра (который рос наклонно), что позволяет предположить, что этот человек смог выжить в течение длительного времени, несмотря на нарушения жевательной способности. [ 10 ]
Общество
[ редактировать ]Высокий уровень полового диморфизма может указывать на относительно интенсивную конкуренцию между самцами, хотя, учитывая, что верхние клыки выступают лишь немного дальше щечных зубов, демонстрация клыков, вероятно, не очень важна в агонистическом поведении, в отличие от современных человекообразных обезьян. [ 5 ]
Палеоэкология
[ редактировать ]
Останки гигантопитеков обычно находят в субтропических вечнозеленых широколиственных лесах Южного Китая, за исключением Хайнаня , где есть тропические тропические леса . Анализ изотопов углерода и кислорода эмали раннего плейстоцена позволяет предположить, что гигантопитеки населяли густой, влажный лес с закрытым пологом. В пещере Кеке был смешанный лиственный и вечнозеленый лес, в котором преобладали березы , дубы и чинкапины , а также несколько низменных трав и папоротников . [ 5 ]
« гигантопитеков Фауна », одна из важнейших групп фауны млекопитающих раннего плейстоцена южного Китая, включает тропические и субтропические лесные виды. Эта группа была разделена на три этапа: 2,6–1,8 миллиона лет назад, 1,8–1,2 миллиона лет назад и 1,2–0,8 миллиона лет назад. Для ранней стадии характерны более древние неогеновые животные, такие как гомфотериид хоботный (родственник слонов) Sinomastodon , халикотер Hesperotherium , суид Hippopotamodon , трагулид Dorcabune и олень Cervavitus . На среднюю стадию указывает появление панды Ailuropoda wulingshanensis , волка Cuon antiquus и тапира Tapirus sinensis . На поздней стадии представлены более типичные животные среднего плейстоцена, такие как панда Ailuropoda bconi и стегодонтидный хоботный Stegodon . [ 33 ] К другим классическим животным обычно относятся орангутанги, макаки , носороги , вымершие свиньи Sus xiaozhu и Sus peii , мунтжак , цервус (олень), гаур (корова), бык Megalovis и, реже, большой саблезубый кот Megantereon . [ 34 ] В 2009 году американский палеоантрополог Рассел Чохон выдвинул гипотезу о том, что неописанная обезьяна размером с шимпанзе, которую он идентифицировал по нескольким зубам, сосуществовала с гигантопитеком . [ 35 ] который в 2019 году был идентифицирован как близкородственный Мегантроп . [ 36 ] Пещера Лунгудонг, возможно, представляла собой переходную зону между Палеарктическим и Восточным царствами, где, наряду с типичной фауной гигантопитеков , обитали более бореальные животные, такие как ежи , гиены , лошади , бычьи лептобо и пищухи . [ 33 ]
Вымирание
[ редактировать ]В течение плейстоцена гигантопитеки , специализирующиеся на окружающей среде , демонстрировали признаки диетических изменений, основанных на морфологии зубов, в ответ на экологическое давление. Другие гоминиды, сосуществовавшие с ним, по сравнению с ним демонстрируют мало серьезных морфологических изменений. [ 5 ] В конце среднего плейстоцена и начале среднего плейстоцена глобальные тенденции указывают на сильные циклы оледенения, более холодный и/или более влажный климат, усиление сезонности и снижение C 4 . численности растений [ 37 ] Усиление зимних и летних муссонов также считается еще одной важной экологической силой в среднем плейстоцене Китая. [ 23 ] Стратиграфические данные свидетельствуют о том, что его диапазон сильно сократился от провинций Гуанси, Гуйчжоу и Хайнань до региона Гуанси на 420–330 тыс. лет назад. Популяция гигантской обезьяны сократилась еще на 300 тыс. лет назад в ответ на сокращение лесных рефугиумов в результате расширения пастбищ. По оценкам, окно вымирания гигантопитеков приходится на период от 295 000 до 215 000 лет назад, что связано с усилением сезонности и значительными изменениями в окружающей среде. [ 4 ] Саванны оставались доминирующей средой обитания Юго-Восточной Азии до позднего плейстоцена . [ 38 ] [ 37 ]
Считается, что вымирание специализированных Gigantopithecus и выживание Pongo weidenreichi plus Homo в среднем плейстоцене было результатом того, что первые изо всех сил пытались адаптироваться к серьезным изменениям окружающей среды по сравнению с двумя последними. P. weidenreichi, вероятно, был более гибким в сезонных пищевых привычках, тогда как Homo был универсальным и, следовательно, был способен адаптироваться к смешанным средам обитания, возникающим в результате изменений окружающей среды в среднем плейстоцене. [ 4 ] Некоторые авторы предположили, что H. erectus был возможным фактором исчезновения Gigantopithecus . [ 39 ] Человеческая деятельность на юге Китая известна еще 800 000 лет назад, но не стала преобладающей до исчезновения гигантопитеков , поэтому неясно, было ли фактором такое давление, как конкуренция за ресурсы или чрезмерная охота. [ 35 ] Чжан и др. ал. в 2024 году предположили, что нет никаких доказательств участия архаичных гомининов в ранних вымираниях плейстоцена на юге Китая. [ 4 ]
Некоторые авторы предполагают, что гигантопитеки , возможно, дожили до раннего позднего плейстоцена , хотя доказательства этого ограничены. [ 40 ]
Криптозоология
[ редактировать ]Гигантопитек использовался в криптозоологических кругах как олицетворение тибетского йети или американского снежного человека , обезьяноподобных монстров в местном фольклоре. Это началось в 1960 году с того, что зоолог Владимир Чернецкий кратко описал в журнале Nature фотографию предполагаемых следов йети 1951 года, сделанную гималайскими альпинистами Майклом Уордом и Эриком Шиптоном . Чернецкий пришел к выводу, что йети ходил как человек и был похож на гигантопитека . Впоследствии йети на короткое время привлек научное внимание, и еще несколько авторов опубликовали свои публикации в журналах Nature and Science , но это также спровоцировало популярную охоту на монстров как за йети, так и за аналогичным американским снежным человеком, которая сохраняется и по сей день. Единственным ученым, который продолжал пытаться доказать существование таких монстров, был антрополог Гровер Кранц , который продолжал настаивать на связи между гигантопитеком и снежным человеком с 1970 года до своей смерти в 2002 году. Среди биномиальных названий, которые он придумал для снежного человека, было «Gigantopithecus canadensis». Ученые и охотники на монстров-любители отвергли аргументы Кранца, заявив, что он с готовностью принял явно ложные доказательства. [ 41 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с фон Кенигсвальд, GHR (1935). «Ископаемая фауна млекопитающих с обезьянами из южного Китая» (PDF) . Труды Королевской академии наук в Амстердаме . 38 (8): 874–879. Архивировано (PDF) из оригинала 12 декабря 2017 г. Проверено 12 декабря 2017 г.
- ^ «Определение гигантопитека | Dictionary.com» . www.dictionary.com . Архивировано из оригинала 3 октября 2022 г. Проверено 2 октября 2022 г.
- ^ Тамисия, Джек (10 января 2024 г.). «Самая большая обезьяна, которая когда-либо жила, не была слишком большой, чтобы потерпеть неудачу — ископаемые зубы показывают, что гигантопитеки были обречены из-за меняющейся окружающей среды и жесткой диеты» . Нью-Йорк Таймс . Архивировано из оригинала 10 января 2024 года . Проверено 11 января 2024 г.
- ^ Jump up to: а б с д Чжан, Инци; Вестэуэй, Кира Э.; Хаберле, Саймон; Любек, Жюльен К.; Бейли, Мэриан; Чохон, Рассел; Морли, Майк В.; Робертс, Патрик; Чжао, Цзянь-синь; Дюваль, Матье; Доссето, Энтони; Пан, Юэ; Правило, Сью; Ляо, Вэй; Галли, Грант А.; Лукас, Мэри; Мо, Джинью; Ян, Лиюнь; Цай, Яньцзюнь; Ван, Вэй; Жоан-Бойо, Рено (10 января 2024 г.). «Гибель гигантской обезьяны Gigantopithecus blacki» . Природа . 625 (7995): 535–539. Бибкод : 2024Natur.625..535Z . дои : 10.1038/s41586-023-06900-0 . ПМЦ 10794149 . ПМИД 38200315 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Чжан, Ю.; Харрисон, Т. (2017). « Gigantopithecus blacki : повторное посещение гигантской обезьяны из плейстоцена Азии» . Американский журнал физической антропологии . 162 (С63): 153–177. дои : 10.1002/ajpa.23150 . ПМИД 28105715 .
- ^ Jump up to: а б с д Хартвиг, WC (2002). Летопись окаменелостей приматов . Издательство Кембриджского университета. стр. 371–372. ISBN 978-0-521-66315-1 .
- ^ Пуарье, FE; Макки, Дж. К. (1999). Понимание эволюции человека (четвертое изд.). Река Аппер-Сэддл, Нью-Джерси: Прентис-Холл. п. 119. ИСБН 0-13-096152-3 .
- ^ Койшон, Р. (1991). «Обезьяна, которая была – азиатские окаменелости показывают гигантского кузена человечества» . Естественная история . 100 : 54–62. ISSN 0028-0712 . Архивировано из оригинала 25 мая 2015 года.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Софван, Н.; и др. (2016). «Гигантский примат Явы: новый экземпляр гигантопитека из Семедо» [Гигантский примат Явы: новый экземпляр гигантопитека из Семедо]. Археологические периодические издания . 36 (2): 141–160. дои : 10.30883/jba.v36i2.241 .
- ^ Jump up to: а б с Чжан, Ю.; Джин, К.; и др. (2015). «Четвертая нижняя челюсть и связанные с ней зубные останки Gigantopithecus blacki из пещеры Яньлян раннего плейстоцена, Фусуй, Гуанси, Южный Китай». Историческая биология . 28 (1–2): 95–104. дои : 10.1080/08912963.2015.1024115 . S2CID 130928802 .
- ^ Брум, Р. (1939). «Зубной ряд антропоидов плейстоцена Трансвааля, плезиантропов и парантропов » (PDF) . Летопись музея Трансвааля . 19 (3): 303–314.
- ^ Jump up to: а б Вайденрайх, Ф. (1946). Обезьяны, гиганты и человек . Издательство Чикагского университета. стр. 58–66 .
- ^ фон Кенигсвальд, GHR (1952). « Gigantopithecus blacki von Koenigswald, гигантский ископаемый гуманоид из плейстоцена южного Китая». Антропологические документы Американского музея естественной истории . 43 : 292–325. HDL : 2246/298 .
- ^ Jump up to: а б Велкер, Ф.; Рамос-Мадригал, Дж.; Кульвильм, М.; и др. (2019). «Протеом эмали показывает, что гигантопитеки были ранними дивергентными понгами» . Природа . 576 (7786): 262–265. Бибкод : 2019Natur.576..262W . дои : 10.1038/s41586-019-1728-8 . ПМК 6908745 . ПМИД 31723270 .
- ^ Салай, Ф.; Делсон, Э. (1979). Эволюционная история приматов . Академическая пресса. стр. 493–494. ISBN 978-1-4832-8925-0 .
- ^ Кэмерон, Д. (2003). «Функциональная и филогенетическая интерпретация позднемиоценового сиваликского гоминида Indopithecus и китайского плейстоценового гоминида Gigantopithecus ». Гималайская геология . 24 :19–28.
- ^ Кэмерон, Д. (2001). «Таксономический статус сиваликского гоминида позднего миоцена Indopithecus (= Gigantopithecus)». Гималайская геология . 22 : 29–34.
- ^ Jump up to: а б Олейничак, AJ; и др. (2008). «Толщина молярной эмали и высота рогов дентина у Gigantopithecus blacki » (PDF) . Американский журнал физической антропологии . 135 (1): 85–91. дои : 10.1002/ajpa.20711 . ПМИД 17941103 . Архивировано из оригинала (PDF) 03 марта 2016 г.
- ^ Джонсон, А.Э. младший (1979). «Оценки скелета гигантопитека на основе аналогии с гориллой». Журнал эволюции человека . 8 (6): 585–587. Бибкод : 1979JHumE...8..585J . дои : 10.1016/0047-2484(79)90111-8 .
- ^ Jump up to: а б с Дин, MC; Шренк, Ф. (2003). «Толщина и развитие эмали третьего постоянного моляра Gigantopithecus blacki ». Журнал эволюции человека . 45 (5): 381–388. Бибкод : 2003JHumE..45..381D . дои : 10.1016/j.jhevol.2003.08.009 . ПМИД 14624748 .
- ^ Чочон, Р. ; и др. (1996). «Датированное совместное появление человека прямоходящего и гигантопитека из пещеры Тхам Хуен, Вьетнам» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (7): 3016–3020. Бибкод : 1996PNAS...93.3016C . дои : 10.1073/pnas.93.7.3016 . ПМЦ 39753 . ПМИД 8610161 .
- ^ Jump up to: а б с д и Коно, РТ; Чжан, Ю.; Джин, К.; Такай, М.; Сува, Г. (2014). «Трехмерная оценка толщины и характера распределения эмали коренных зубов у Gigantopithecus blacki ». Четвертичный интернационал . 354 : 46–51. Бибкод : 2014QuInt.354...46K . дои : 10.1016/j.quaint.2014.02.012 .
- ^ Jump up to: а б Шао, кв.; Ван, Ю.; и др. (2017). «Датирование фауны стегодонов и айлуроподов серией U и ESR / U в Черной пещере, Гуанси, южный Китай, с учетом сроков исчезновения Gigantopithecus blacki ». Четвертичный интернационал . 434 : 65–74. Бибкод : 2017QuInt.434...65S . дои : 10.1016/j.quaint.2015.12.016 .
- ^ Jump up to: а б с Бохеренс, Х.; Шренк, Ф.; Чаймани, Ю.; Куллмер, О.; Мёрике, Д.; Пушкина, Д.; Йегер, Ж.-Ж. (2017). «Гибкость диеты и среды обитания плейстоценовых млекопитающих Южной Азии: последствия для судьбы гигантской ископаемой обезьяны Gigantopithecus ». Четвертичный интернационал . 434 : 148–155. Бибкод : 2017QuInt.434..148B . дои : 10.1016/j.quaint.2015.11.059 .
- ^ Купчик, К.; Дин, MC (2008). «Сравнительные наблюдения за морфологией корней зубов Gigantopithecus blacki ». Журнал эволюции человека . 54 (2): 196–204. Бибкод : 2008JHumE..54..196K . дои : 10.1016/j.jhevol.2007.09.013 . ПМИД 18045651 .
- ^ Jump up to: а б с д Чочон, Р. ; Пиперно, ДР; Томпсон, Р.Г. (1990). «Опаловые фитолиты, обнаруженные на зубах вымершей обезьяны Gigantopithecus blacki : значение для палеодиетических исследований» . Труды Национальной академии наук . 87 (20): 8120–8124. Бибкод : 1990PNAS...87.8120C . дои : 10.1073/pnas.87.20.8120 . ПМК 54904 . ПМИД 2236026 .
- ^ Цюй, Ю.; Джин, К.; Чжан, Ю.; и др. (2014). «Оценка сохранности и анализ изотопов углерода и кислорода зубной эмали Gigantopithecus blacki и современных животных из пещеры Санхэ, Чунцзуо, Южный Китай, во время раннего плейстоцена». Четвертичный интернационал . 354 : 52–58. Бибкод : 2014QuInt.354...52Q . дои : 10.1016/j.quaint.2013.10.053 .
- ^ Чжан, Ю.; Коно, РТ; Джин, К.; Ван, В.; Харрисон, Т. (2014). «Возможное изменение морфологии зубов у Gigantopithecus blacki незадолго до его исчезновения: данные по стыку эмали и дентина верхних премоляров». Журнал эволюции человека . 75 : 166–171. Бибкод : 2014JHumE..75..166Z . дои : 10.1016/j.jhevol.2014.06.010 . ПМИД 25063565 .
- ^ Чжан, Ю.; Коно, РТ; Джин, К.; Ван, В.; и др. (2014). «Новые останки Gigantopithecus blacki возрастом 400–320 тыс. лет из пещеры Хэцзян, город Чунцзуо, Гуанси, Южный Китай». Четвертичный интернационал . 354 : 35–45. Бибкод : 2014QuInt.354...35Z . дои : 10.1016/j.quaint.2013.12.008 .
- ^ Пей, В. (1957). «В Китае обнаружена челюстная кость гигантской обезьяны» . Американский антрополог . 59 (5): 834–838. дои : 10.1525/aa.1957.59.5.02a00080 .
- ^ Уайт, Т.Д. (1975). «Геоморфология и палеоэкология: переоценка гигантопитеков ». Журнал эволюции человека . 4 (3): 219–233. Бибкод : 1975JHumE...4..219W . дои : 10.1016/0047-2484(75)90009-3 .
- ^ Чжан, Инци; Вестэуэй, Кира Э.; Хаберле, Саймон; Любек, Жюльен К.; Бейли, Мэриан; Чохон, Рассел; Морли, Майк В.; Робертс, Патрик; Чжао, Цзянь-синь; Дюваль, Матье; Доссето, Энтони; Пан, Юэ; Правило, Сью; Ляо, Вэй; Галли, Грант А.; Лукас, Мэри; Мо, Джинью; Ян, Лиюнь; Цай, Яньцзюнь; Ван, Вэй; Жоан-Бойо, Рено (10 января 2024 г.). «Гибель гигантской обезьяны Gigantopithecus blacki » . Природа . 625 (7995): 535–539. Бибкод : 2024Natur.625..535Z . дои : 10.1038/s41586-023-06900-0 . ПМЦ 10794149 . ПМИД 38200315 .
- ^ Jump up to: а б Джин, К.; Ван, Ю.; и др. (2014). «Хронологическая последовательность фаун гигантопитеков раннего плейстоцена из пещер в Чунцзо, район реки Цзоцзян, Южный Китай». Четвертичный интернационал . 354 : 4–14. Бибкод : 2014QuInt.354....4J . дои : 10.1016/j.quaint.2013.12.051 .
- ^ Чжу, М.; Шуберт, Б.В.; Лю, Дж.; Уоллес, Южная Каролина (2014). «Новая находка саблезубого кота Мегантереона (Felidae, Machairodontinae) из фауны гигантопитеков раннего плейстоцена , пещера Яньлян, Фусуй, Гуанси, Южный Китай». Четвертичный интернационал . 354 : 100–109. Бибкод : 2014QuInt.354..100Z . дои : 10.1016/j.quaint.2014.06.052 .
- ^ Jump up to: а б Чочон, Р. (2009). «Таинственная обезьяна плейстоценовой Азии» . Природа . 459 (7249): 910–911. Бибкод : 2009Natur.459..910C . дои : 10.1038/459910а . ПМИД 19536242 .
- ^ Занолли, Клеман; Куллмер, Оттмар; Келли, Джей; Бэкон, Анн-Мари; Деметра, Фабрис; Дюмонсель, Жан; Фиоренца, Лука; Грин, Фредерик Э.; Юблин, Жан-Жак; Нгуен, Ань Туан; Нгуен, Ти Май Хуонг (май 2019 г.). «Доказательства увеличения разнообразия гоминид в раннем и среднем плейстоцене Индонезии» (PDF) . Экология и эволюция природы . 3 (5): 755–764. Бибкод : 2019NatEE...3..755Z . дои : 10.1038/s41559-019-0860-z . ISSN 2397-334X . ПМИД 30962558 . S2CID 102353734 . Проверено 17 ноября 2022 г.
- ^ Jump up to: а б Сан, Фаджун; Ван, Ян; Ван, Юань; Цзинь, Чан-чжу; Дэн, Тао; Вольф, Берт (15 июня 2019 г.). «Палеоэкология млекопитающих плейстоцена и палеоклиматические изменения в Южном Китае: данные по стабильным изотопам углерода и кислорода» . Палеогеография, Палеоклиматология, Палеоэкология . 524 : 1–12. Бибкод : 2019PPP...524....1S . дои : 10.1016/j.palaeo.2019.03.021 . S2CID 134558136 . Архивировано из оригинала 14 ноября 2022 года . Проверено 14 ноября 2022 г.
- ^ Луис, Дж.; Робертс, П. (2020). «Экологические факторы вымирания мегафауны и гомининов в Юго-Восточной Азии». Природа . 586 (7829): 402–406. Бибкод : 2020Natur.586..402L . дои : 10.1038/s41586-020-2810-y . hdl : 10072/402368 . ПМИД 33029012 . S2CID 222217295 .
- ^ Чжао, LX; Чжан, LZ (12 февраля 2013 г.). «Новые ископаемые свидетельства и анализ диеты Gigantopithecus blacki , а также его распространение и исчезновение в Южном Китае». Четвертичный интернационал . 286 : 69–74. Бибкод : 2013QuInt.286...69Z . дои : 10.1016/j.quaint.2011.12.016 . ISSN 1040-6182 .
- ^ Лопатин А.В.; Мащенко Е.Н.; Дак, Ле Суан (июнь 2022 г.). «Gigantopithecus blacki (Primates, Ponginae) из пещеры Ланг Транг (Северный Вьетнам): последний гигантопитек в позднем плейстоцене?» . Доклады биологических наук . 502 (1): 6–10. дои : 10.1134/S0012496622010069 . ISSN 0012-4966 . PMID 35298746 . S2CID 254413457 .
- ^ Регал, Б. (2008). «Любитель против профессионала: поиски снежного человека». Стараться . 32 (2): 53–57. doi : 10.1016/j.endeavour.2008.04.005 . ПМИД 18514914 .



| Базы данных органов управления : Национальные |
|---|
- Понгине
- Доисторические обезьяны
- Роды доисторических приматов
- Плейстоценовые приматы
- Плейстоценовые млекопитающие Азии
- Окаменелости Китая
- Вымершие животные Китая
- Окаменелости Индии
- Вымершие животные Индии
- Окаменелости Вьетнама
- Вымершие животные Вьетнама
- Окаменелости Индонезии
- Вымершие животные Индонезии
- Первые выступления Тортониана
- Вымирание родов плейстоцена
- Ископаемые таксоны описаны в 1935 г.
- Таксоны, названные Густавом Генрихом Ральфом фон Кенигсвальдом
- Млекопитающие описаны в 1935 г.