Дождевой червя
| Дождевой червя Временный диапазон:
| |
|---|---|

| |
| Неопознанный вид дождевого червя с хорошо развитым клителлум | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Phylum: | Annelida |
| Clade: | Pleistoannelida |
| Clade: | Sedentaria |
| Class: | Clitellata |
| Order: | Opisthopora |
| Suborder: | Lumbricina |
Земля дождевого червя -это земляный земляный , беспозвоночный который принадлежит к филам Аннелиде . Термин является общим названием для крупнейших членов класса ( или подкласса , в зависимости от автора) Oligochaeta . В классических системах они находились в порядке opisthopora , так как мужские поры открывались сзади к женским порам, хотя внутренние мужские сегменты находятся перед женщинами. Теоретические кладистические исследования поместили их в подряд Lumbricina порядка из гаплотаксиды , но это может измениться. [ нужно разъяснения ] Другие названия сленга для дождевых червей включают «Dew-Worm», «Rainworm», «Nightcrawler» и «Engleworm» (от его использования в качестве рыболовных крючков ). Большие земляные черви также называют мегадрилами (что переводится как «большие черви»), в отличие от микроодрилов («маленькие черви») в полуаватических семействах Tubificidae , Lumbricidae и Enchytraeidae . Мегадрилы характеризуются отчетливым Clitellum (более обширным, чем у микродарилов) и сосудистой системой с настоящими капиллярами . [ 2 ]
Земные черви обычно встречаются в влажной, компостной почве, есть широкий спектр органических вопросов , [ 3 ] который включает в себя детрит , простейшие простейшие , надеть , нематоды , бактерии , грибы и другие микроорганизмы . [ 4 ] An earthworm's digestive system runs the length of its body.[5] природы Они являются одним из самых важных детритоядных и копрофагов , а также служат в качестве пищи низкого уровня для многих потребителей в экосистемах.
Earthworms exhibit an externally segmented tube-within-a-tube body plan with corresponding internal segmentations, and usually have setae on all segments.[6] They have a cosmopolitan distribution wherever soil, water and temperature conditions allow.[7] They have a double transport system made of coelomic fluid that moves within the fluid-filled coelom and a simple, closed circulatory system, and respire (breathe) via cutaneous respiration. As soft-bodied invertebrates, they lack a true skeleton, but their structure is maintained by fluid-filled coelom chambers that function as a hydrostatic skeleton.[citation needed]
Earthworms have a central nervous system consisting of two ganglia above the mouth, one on either side, connected to an axial nerve running along its length to motor neurons and sensory cells in each segment. Large numbers of chemoreceptors concentrate near its mouth. Circumferential and longitudinal muscles edging each segment let the worm move. Similar sets of muscles line the gut tube, and their actions propel digested food toward the worm's anus.[8]
Earthworms are hermaphrodites: each worm carries male and female reproductive organs and genital pores. When mating, two individual earthworms will exchange sperm and fertilize each other's ova.
Anatomy
Form and function
Depending on the species, an adult earthworm can be from 10 mm (0.39 in) long and 1 mm (0.039 in) wide to 3 m (9.8 ft) long and over 25 mm (0.98 in) wide, but the typical Lumbricus terrestris grows to about 360 mm (14 in) long.[9] Probably the longest worm on confirmed records is Amynthas mekongianus that extends up to 3 m (10 ft) [10] in the mud along the banks of the 4,350 km (2,703 mi) Mekong River in Southeast Asia.
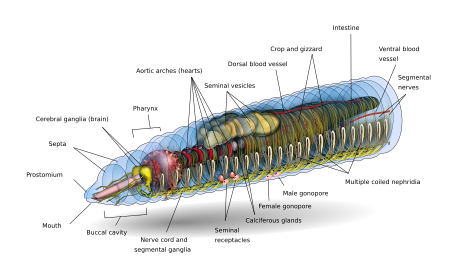
Спереди к спине основная форма дождевой червя представляет собой цилиндрическую трубку в трубке, разделенную на серию сегментов (называемые метамерами ), которые разделяют тело. Пороты обычно [11] Внешне видны на теле, пожимая сегменты; Спинные поры и нефридиопоры источают жидкость, которая увлажняет и защищает поверхность червя, позволяя ему дышать. За исключением рта и анальных сегментов, каждый сегмент носит гристлевые волосы, называемые боковыми щетинками [ 12 ] используется для закрепления частей тела во время движения; [ 13 ] Виды могут иметь четыре пары щетинок на каждом сегменте или более восьми, иногда образуя полный круг щетинок на сегмент. [ 12 ] Специальные вентральные щетинки используются для закрепления спаривающихся дождевых червей путем их проникновения в тела своих товарищей. [ 14 ]
Как правило, внутри вида количество обнаруженных сегментов согласуется между образцами, и люди рождаются с количеством сегментов, которые они будут иметь на протяжении всей своей жизни. Первый сегмент тела (сегмент № 1) оснащен как ртом дождевого червя, так и, нависающего над ртом, мясистой доли, называемой простомиумом , который закрывает вход, когда червь находится в состоянии покоя, но также используется для ощущения и химического ощущения окружающей среды червя. Некоторые виды дождевого червя могут даже использовать преусильный простомиум, чтобы захватить и перетаскивать такие предметы, как травы и листья в их нору.
У взрослого дождевого червя появляется поясная железистая железа в форме пояса, называемый Clitellum , который охватывает несколько сегментов в направлении передней части животного. Это является частью репродуктивной системы и производит яичные капсулы. Задняя часть чаще всего является цилиндрическим, как и остальная часть тела, но в зависимости от вида, это также может быть четырехугольной, восьмиугольной, трапеции или сплющенной. Последний сегмент называется перипроктом ; Анус дождевого червя, короткая вертикальная щель, находится в этом сегменте. [ 12 ]

Внешняя внешность отдельного сегмента представляет собой тонкую кутикулу над кожей, обычно пигментированная от красного до коричневого цвета, которая имеет специализированные клетки, которые выделяют слизь над кутикулой, чтобы организовало тело влажным и легким движением через почву. Под кожей находится слой нервной ткани и два слоя мышц - тонкий внешний слой круговой мышцы и гораздо более толстый внутренний слой продольной мышцы. [ 15 ] Интерьер на мышечный слой-это камера, заполненная жидкостью, называемая Coelom [ 16 ] Это по его давлению обеспечивает структуру для тела без костей червя. Сегменты отделены друг от друга септа (множественное число «перегородки») [ 17 ] которые представляют собой перфорированные поперечные стенки, позволяющие целомической жидкости проходить между сегментами. [ 18 ] Пара структур, называемых нефростомами , расположены в задней части каждой перегородки; Нефрическая канальца ведет из каждого нефростома через перегородку и в следующий сегмент. Затем этот канал приводит к основному органу фильтрации жидкости, нефридий или метанефридиуму, который удаляет метаболические отходы из целомической жидкости и вытесняет его через поры, называемые нефридиопоры по бокам червя; Обычно в большинстве сегментов встречаются две нефридии (иногда больше). [ 19 ] В центре червя находится пищеварительный тракт , который проходит прямо из устья к задницам без анета, и окружено над и внизу кровеносными сосудами (дорсальный кровеносный сосуд и вентральный кровеносный сосуд, а также субнеральные кровеносные сосуды) и вентральный нервный шнур и окружен в каждом сегменте парой кровлиальных васелей, которые соединяют дельсоры.
Многие дождевые черви могут выбросить целомическую жидкость через поры сзади в ответ на стресс; Австралийский Didymogaster Sylvaticus (известный как «голубой дождевой червя») может впрыгивать жидкостью до 30 см (12 дюймов). [ 20 ] [ 18 ]
Нервная система

Центральная нервная система
CNS состоит из двулобированного мозга (церебральные ганглии , или супрафарингеальные ганглии), субфарингеальных ганглиев, окружных соединительных связей и вентрального нервного шнура .
Мозги дождевых червей состоят из пары грушевидных головных ганглиев. Они расположены в дорсальной стороне пищеварческого канала в третьем сегменте, в канавке между полостью щека и глотки .
Пара окружных фарингеальных соединений из мозга окружает глотку, а затем соединяется с парой подфарингеальных ганглий, расположенных под глоткой в четвертом сегменте. Такое расположение означает мозг, субфарингеальные ганглии и окружных связующих соединений образуют нервное кольцо вокруг глотки.
Вентральный нервный шнур (образованный нервными клетками и нервными волокнами) начинается в субфарингеальных ганглиях и простирается под пищевым каналом до наиболее заднего сегмента тела. Вентральный нервный шнур имеет отек, или ганглион, в каждом сегменте, то есть сегментальный ганглион, который происходит от пятого до последнего сегмента тела. Есть также три гигантских аксона , один медиальный гигантский аксон (MGA) и два боковых гигантских аксона (LGA) на средней части вентрального нервного шнура. MGA имеет диаметр 0,07 мм и передает в переднем ходовом направлении со скоростью 32,2 м/с. LGA немного более узкие диаметром 0,05 мм и передают в заднем дворчащем направлении при 12,6 м/с. Два LGA связаны через регулярные промежутки времени вдоль тела и поэтому считаются одним гигантским аксоном. [ 21 ] [ 22 ]
Периферическая нервная система
- Восемь -десять нервов возникают из головных ганглиев, чтобы поставить простомий , пировую камеру и глотку .
- Три пары нервов возникают из подражающих ганглиев для предоставления второго, третьего и четвертого сегмента.
- Три пары нервов простираются от каждого сегментарного ганглия для обеспечения различных структур сегмента.
Симпатическая нервная система состоит из нервных сплетений в эпидермисе и пищеварительном канале. (Сплетение - это сеть соединенных нервных клеток.) Нервы, которые проходят вдоль прохода стенки тела между внешним круговым и внутренним продольным мышечным слоями стенки. Они выделяют ветви, которые образуют межмышечное сплетение и субэпидермальное сплетение. Эти нервы связаны с соединением CricoPharyNgeal.
Движение

На первый взгляд скорость ползания варьируется как внутри, так и среди людей. Земные червя ползают быстрее, прежде всего, делая более длинные «шаги» и большую частоту шагов. Большие черви Lumbricus Tarrestris ползают с большей абсолютной скоростью, чем меньшие черви. Они достигают этого, делая немного более длинные шаги, но с немного более низкими частотами шага. [ 23 ]
Прикосновение к дождевым червям, который вызывает реакцию «давления», а также (часто) реакцию на обезвоживающее качество соли на коже человека (токсичные с дождевыми червями), стимулирует субэпидермальный нервный плекс, который соединяется с межмышечным сплетением и вызывает сокращение продольных мышц. Это приводит к тому, что корстающие движения наблюдаются, когда человек поднимает дождевой червя. Такое поведение является рефлексом и не требует ЦНС; Это происходит, даже если нервный шнур удаляется. Каждый сегмент дождевого червя имеет собственное нервное сплетение. Сплетение одного сегмента не связано непосредственно с соседним сегментами. Для соединения нервных систем сегментов необходим нервный шнур. [ 24 ]
Гигантские аксоны несут самые быстрые сигналы вдоль нервного шнура. Это аварийные сигналы, которые инициируют поведение рефлекса. Большой дорсальный гигантский аксон проводит сигналы самых быстрых, от задней части до передней части животного. Если задняя часть червя касается, сигнал быстро отправляется вперед, вызывая сжимание продольных мышц в каждом сегменте. Это приводит к тому, что червь очень быстро сокращается как попытка убежать от хищника или другой потенциальной угрозы. Два медиальных гигантских аксонов соединяются друг с другом и отправляют сигналы спереди к задней части. Стимуляция их заставляет дождевой червя очень быстро отступить (возможно, сжимаясь в его норе, чтобы избежать птицы).
Присутствие нервной системы необходимо для того, чтобы животное могло испытать ноцицепцию или боль . Тем не менее, также требуются другие физиологические возможности, такие как опиоидная чувствительность и центральная модуляция ответов анальгетиками. [ 25 ] Энкефалин и α -эндорфиновые вещества были обнаружены в дождевых червях. Инъекции налоксона (опиоидный антагонист) ингибируют экономные реакции дождевых червей. Это указывает на то, что опиоидные вещества играют роль в сенсорной модуляции, аналогично тем, которая встречается у многих позвоночных. [ 26 ]
Сенсорный прием
Светочувствительность
Хотя у некоторых червей есть глаза , дождевые черви нет. Тем не менее, они имеют специализированные фоточувствительные клетки, называемые «светлыми клетками Hess». Эти фоторецепторные клетки имеют центральную внутриклеточную полость ( фасома ), заполненную микроворками . Наряду с микроворками, в фаосоме есть несколько сенсорных ресничек, которые структурно не зависят от микроворсинок. [ 27 ] Фоторецепторы распределяются в большинстве частей эпидермиса, но более концентрируются на спине и на сторонах червя. Относительно небольшое число происходит на вентральной поверхности первого сегмента. Они наиболее многочисленны в простомии и снижают плотность в первых трех сегментах; Их очень мало в течение третьего сегмента. [ 24 ]
Эпидермальный рецептор (орган чувств)
Эти рецепторы обильны и распределены по всему эпидермису . Каждый рецептор показывает слегка повышенную кутикулу, которая охватывает группу высоких, тонких и столбчатых рецепторных клеток. Эти клетки несут небольшие родовые процессы на их внешних концах, а их внутренние концы связаны с нервными волокнами. Эпидермальные рецепторы тактильны в функции. Они также обеспокоены изменениями температуры и реагируют на химические стимулы. Земные черви чрезвычайно чувствительны к прикосновению и механической вибрации.
Buccal (Sense Organ) Рецептор
Эти рецепторы расположены только в эпителии чистой камеры. Эти рецепторы вкусовые и обонятельные (связанные со вкусом и запахом). Они также реагируют на химические стимулы. (Хеморецептор)
Пищеварительная система
дождевого В кишечнике червя - прямая трубка, которая простирается от уст червя до его ануса . Он дифференцируется в пищевой канал и связанные железы, которые встроены в стену самого пищевого канала. Элементарный канал состоит из рта, щекальной полости (обычно проходит через первый или два сегмента дождевого червя), глотки (в целом около четырех сегментов), пищевода, урожая, живота (обычно) и кишечника. [ 28 ]
Еда входит в рот. Главная глотка действует как всасывающий насос; Его мышечные стены привлекают пищу. В глотке глоточные железы секретируют слизь . Пища движется в пищевод , где кальций (от крови и проглатывается из предыдущих приемов пищи), чтобы поддерживать правильный уровень кальция в крови в крови и pH пищи . Оттуда еда проходит в урожай и желудок. В желудце сильные мышечные сокращения размалывают пищу с помощью минеральных частиц, проглатываемых вместе с пищей. Оказавшись через желудку, еда продолжается через кишечник для пищеварения. Кишечный кишечник выделяет пепсин для переваривания белков, амилазы для переваривания полисахаридов, целлюлазы для переваривания целлюлозы и липазы для переваривания жиров. [ 8 ] Значительные черви используют, в дополнение к пищеварительным белкам, класс активных поверхностных соединений, называемых дрилодефенсинами , которые помогают переваривать растительный материал. [ 29 ] Вместо того, чтобы быть свернутым, как кишечник млекопитающего, в кишечнике дождевого червя присутствует большая средняя дорсальная складка, называемая тифлозолом , причем многие складки бегут по ее длине, увеличивая площадь поверхности, чтобы увеличить поглощение питательных веществ. В кишечнике есть своя пара мышечных слоев, таких как тело, но в обратном порядке - внутренний круглый слой во внешнем продольном слое. [ 30 ]
Система кровообращения
Земные черви имеют двойную циркуляторную систему, в которой как целомическая жидкость, так и замкнутая система кровообращения несут пищевые продукты, отходы и дыхательные газы. Закрытая система кровообращения имеет пять основных кровеносных сосудов: дорсальный (верхний) сосуд, который работает над пищеварительным трактом; вентральный (нижний) сосуд, который работает под пищеварительным трактом; субневральный сосуд, который проходит ниже вентрального нервного шнура; и два поздних сосуда по обе стороны от нервного шнура. [ 31 ]
Спинной сосуд является в основном структурой сбора в кишечной области. Он получает пару комиссуральных и дорсальных кишечников в каждом сегменте. Вентральный сосуд разветвляется до пары вентротегментарных и вентро-заводов в каждом сегменте. Субневральное судно также выдает пару комиссуров, проходящих вдоль задней поверхности перегородки.
Насосное действие на дорсальном сосуде движется кровью вперед, в то время как остальные четыре продольных сосуда несут кровь назад. В сегментах седьмого до одиннадцати пара артов аорты звонит в Coelom и действует как сердца, перекачивая кровь в вентральный сосуд, который действует как аорта. Кровь состоит из амебоидных клеток и гемоглобина, растворенного в плазме. Вторая система кровообращения вытекает из ячеек пищеварительной системы, которые выравнивают Coelom. Когда пищеварительные клетки становятся полными, они высвобождают неживущие клетки жира в заполненный жидкостью Coelom, где они свободно плавают, но могут проходить через стены, разделяющие каждый сегмент, перемещая пищу в другие части и помогают в заживлении ран. [ 32 ]
Экскреторная система
Экспериторная система содержит пару нефридий в каждом сегменте, за исключением первых трех и последних. [ 33 ] Три типа нефридий: покровные, перегородки и глоточный. Интенсивная нефридия лежит прикрепленной к внутренней стороне стены тела во всех сегментах, кроме первых двух. Септальная нефридия прикреплена к обеим сторонам септы за 15 -м сегментом. Нефридия глотки прикреплена к четвертому, пятому и шестому сегментам. [ 33 ] жидкости Coelom от переднего сегмента привлечены избиением ресничек нефростома . Отходы в Оттуда он переносится через перегородку (стену) через трубку, которая образует серию петель, переплетенные капиллярами крови, которые также переносят отходы в канальца нефростома. Экскреторные отходы затем наконец выгружаются через пор на стороне червя. [ 34 ]
Дыхание
Землю не имеют особых респираторных органов. Газы обмениваются через влажную кожу и капилляры, где кислород поднимается гемоглобином, растворенным в плазме крови и выпускается углекислый газ. Вода, а также соли, также можно перемещать через кожу активным транспортом.
Жизнь и физиология
При рождении дождевые черви появляются небольшими, но полностью сформированными, не имея только своих половых структур, которые развиваются примерно через 60-90 дней. Они достигают полного размера примерно за год. Ученые предсказывают, что средняя продолжительность жизни в полевых условиях составляет от четырех до восьми лет, в то время как большинство сортов сада проживают только один -два года.
Размножение



Несколько распространенных видов дождевых червей в основном партеногенетические , что означает, что рост и развитие эмбрионов происходит без оплодотворения . Среди ламбрицид дождевых червей партеногенез возникал из -за сексуальных родственников много раз. [ 35 ] Партеногенез у некоторых линий Trapezoides Aprectodea выступил от 6,4 до 1,1 миллиона лет назад от сексуальных предков. [ 36 ] Несколько видов демонстрируют псевдогамный партогенез, что означает, что спаривание необходимо для стимуляции размножения, хотя ни один мужской генетический материал не передает потомству. [ 37 ]
Спечный спаривание дождевого червя происходит на поверхности, чаще всего ночью. Землемы - гермафродиты ; То есть у них есть как мужские, так и женские сексуальные органы. Сексуальные органы расположены в сегментах с 9 по 15. Престоносные черви имеют одну или две пары яичек, содержащихся в мешках. Две или четыре пары семенных пузырьков производят, хранят и выпускают сперму через мужские поры. Яичники и яйцеклетки в сегменте 13 высвобождают яйца через женские поры на сегменте 14, в то время как сперматозоиды удаляются из сегмента 15. Одна или несколько пар сперматеков присутствует в сегментах 9 и 10 (в зависимости от видов), которые являются внутренними мешками, которые получают и хранят сперму от другого червя во время совокупления. В результате 15 -й сегмент одного червя источает сперму в сегменты 9 и 10 с его везикулами хранилища своего партнера. Некоторые виды используют внешние сперматофоры для переноса сперматозоидов.
У Hormogaster Samnitica и Hormogaster Elisae Transcriptome DNA библиотеки ДНК были секвенированы, а два половых феромона , притяжение и искушение, были обнаружены во всех образцах ткани обоих видов . [ 38 ] Половые феромоны, вероятно, важны у дождевых червей, потому что они живут в среде, где химическая сигнализация может играть решающую роль в привлечении партнера и в содействии скрещиваниям. Скротительство обеспечит преимущество маскировки экспрессии вредных рецессивных мутаций в потомстве [ 39 ] (См. комплементацию ).
Копуляция и размножение являются отдельными процессами в дождевых червях. Пара спаривания перекрывает передние концы вентрально, а каждый обменивает сперму с другой. Клителлум . становится очень красноватым до розоваточного цвета Через некоторое время после совокупления, спустя долгое время после того, как черви расстались, Clitellum (позади сперматеков) выделяет материал, который образует кольцо вокруг червя. Червь затем выходит из кольца, и, как и это, он вводит свои собственные яйца и сперму другого червя. Таким образом, каждый червь становится генетическим отцом некоторых их потомков (из -за его собственной спермы, перенесенной на другие дождевые червя) и генетической матери (потомства из своих собственных яичных клеток) остальных. Когда червь выдвигается из кольца, концы коконского уплотнения образуют смутно луковичный инкубатор ( кокон ), в котором развиваются эмбриональные черви. Следовательно, оплодотворение является внешним. Затем кокон осаждается в почве. Через три недели от 2 до 20 потомства люк со средним уровнем 4. Разработка является прямой т.е. без формирования личинки.
Репарация ДНК
Воздействие дождевой червя Эйзения Фетда на ионизирующее излучение, индуцированное разрывами ДНК и окисленными основаниями ДНК . [ 40 ] Эти повреждения ДНК могут затем быть восстановлены в соматических и сперматогенных клетках. [ 40 ] Клетки яичков дождевых червей также способны восстанавливать перекисью водорода . индуцированные окислительные ДНК -аддукты, индуцированные [ 41 ]
Локомоция

Дождевые черви проходят под землей посредством волн мышечных сокращений, которые попеременно сокращают и удлиняют тело ( перистальтис ). Укороченная часть привязана к окружающей почве крошечными когтями щетинками ( щетинками ), установленными вдоль ее сегментированной длины. Во всех сегментах тела, за исключением первого, последнего и клинеллума, в эпидермальную яму каждого сегмента есть кольцо с S-образными щетинками. Весь процесс обрывания помогает секреция смазочной слизи. В результате их движения через их смазываемые туннели, черви могут издавать булькающие звуки под землей при нарушении. Дождевые черви движутся по почве, расширяя расщелины с силой; Когда силы измеряются в зависимости от массы тела, мыслиние могут увеличить их собственную массу тела в 500 раз, тогда как крупные взрослые могут продвигать только в 10 раз их собственную массу тела. [ 42 ]
Регенерация
Земные черви обладают способностью регенерировать утраченные сегменты, но эта способность варьируется между видами и зависит от степени ущерба. Стивенсон (1930) посвятил главу своей монографии этой теме, в то время как Ge Gates потратил 20 лет, изучая регенерацию у различных видов. Но «потому что был проявлен небольшой интерес», Гейтс (1972) опубликовал лишь несколько из его выводов. Тем не менее, они показывают, что теоретически возможно выращивать двух целых червей из раздела пополам в определенных видах.
Отчеты Гейтса включали:
- Eisenia fetida ( Savigny , 1826) с регенерацией головы в переднем направлении, возможно, на каждом межсегментарном уровне назад к и включению 23/24, в то время как хвосты регенерировали на любых уровнях за 20/21; Таким образом, два черви могут расти из одного. [ 43 ]
- Lumbricus Tarrestris ( Linnaeus , 1758 ) заменил передние сегменты с 13/14 и 16/17, но регенерация хвоста никогда не была найдена.
- Perionyx Excavatus (Perrier, 1872) легко регенерировал утраченные части тела, в переднем направлении от 17/18 и в заднем направлении до 20/21.
- Lampito Mauritii (Kinberg, 1867) с регенерацией в переднем направлении на всех уровнях назад до 25/26 и регенерации хвоста с 30/31; Иногда считалось, что регенерация головы вызвана внутренней ампутацией, возникающей в результате Sarcophaga sp. заражение личинками.
- Criodrilus lacuum (Hoffmeister, 1845) также обладает потрясающей регенеративной способностью с регенерацией «головы» с 40/41. [ 44 ]
Сообщалось о неопознаваемом тасманском дождевом червях. [ 45 ]
Таксономия и распределение
В мире таксономии стабильная «классическая система» Майклсена (1900) и Стивенсона (1930) была постепенно разрушена спорами о том, как классифицировать дождевых червей, такие что Fender и McKey Fender (1990) зашли так далеко, чтобы сказать, «классификация семейных уровней мегасколетических земляных червей находится в chaos». [ 46 ] За эти годы многие ученые разработали свои собственные системы классификации для дождевых червей, что привело к путанице, и эти системы и по -прежнему продолжаются пересмотрены и обновляются. Система классификации, используемая здесь, которая была разработана Blakemore (2000), представляет собой современную возврат к классической системе, которая исторически доказана и широко принята. [ 47 ]
Категоризация мегадрильного дождевого червя в одну из его таксономических семей под подчиненными Lumbricina и Moniligastrida основана на таких особенностях, как состав Clitellum, местоположение и расположение пола (поры, простатические железы и т. Д.), Количество Gizzards и форму тела. [ 47 ] В настоящее время названо более 6000 видов земных червей, как указано в базе данных названия вида, [ 48 ] Но количество синонимов неизвестно.
Семьи, с их известными распределениями или происхождением: [ 47 ]
- Acanthodrilidae
- Ailoscolecidae - Пиренеи и Юго -Восточный США
- Almidae -тропический экваториальный (Южная Америка, Африка, Индо-Азия)
- Benhamiinae - Эфиопская, неотропическая (возможная подсемейство Octochaetidae)
- Criodrilidae - юго -западный палеарктический: Европа, Ближний Восток, Россия и Сибири до Тихоокеанского побережья; Япония (Biwadrilus); в основном водный
- Diplocardily /-ideae-гендуань или Laurance? (поднофиляция актодриального)
- Enchytraeidae - космополитическое, но необычное в тропиках (обычно классифицируется с микродарилами)
- Eudrilidae - тропическая Африка к югу от Сахары
- Exxidae - Neotropical: Центральная Америка и Карибский бассейн
- Glossoscolecidae - Neotropical: Центральная и Южная Америка, Карибский бассейн
- Haplotaxidae - Cosmopolitan Distribution (обычно классифицируется с микродарилами)
- Hormogastridae - Средиземноморье
- Kynotidae - Малагасиан: Мадагаскар
- Lumbricidae - Holarctic: Северная Америка, Европа, Ближний Восток, Центральная Азия в Японию
- Lutodrilidae - Луизиана Юго -Восточная США
- Megascolecidae
- Microchaetidae - наземные в Африке, особенно южноафриканские луга
- Moniguilia - восточные и индийские раны
- Ocnerodrilidae - Neotropics, Африка; Индия
- Octochaetidae - австралийский, индийский, восточный, эфиопский, неотропический
- Octochaetinae - австралийская, индийская, восточная (подсемейство, если принимается Benhamiinae)
- Sparganophilidae - Nearctic, Neotropical: North и Central America
- Tumakidae - Колумбия, Южная Америка
Как инвазивный вид
В общей сложности около 7000 видов только около 150 видов широко распространены по всему миру. Это перогрин или космополитические дождевые черви. [ 49 ] Из 182 таксонов дождевых червей, найденных в Соединенных Штатах и Канаде, 60 (33%) являются введенными видами.
Экология

Земные черви классифицируются по трем основным экофизиологическим категориям: (1) листовой мусорец -или компост-брюки, которые не становятся нежирующими, живут на границе раздела почвы и питаются разлагающимися органическими веществами ( эпигеей ), например, Eisenia fetida ; (2) червей, содержащих верхний слой или подсылки, которые питаются (на почве), норы и отливают в почве, создавая горизонтальные норы в верхних 10–30 см почвы ( эндогея ); и (3) черви, которые строят постоянные глубокие вертикальные норы, которые они используют для посещения поверхности для получения растительного материала для пищи, таких как листья ( анеция , что означает «достижение»), например, Lumbricus Tarrestris . [ 50 ]
Популяции дождевых червей зависят как от физических, так и химических свойств почвы, таких как температура, влажность, pH, соли, аэрация и текстура, а также доступная пищевая продукция, а также способность вида воспроизводить и рассеивать. Одним из наиболее важных факторов окружающей среды является рН , но дождевые черви варьируются по их предпочтениям. Большинство предпочитают нейтральные до слегка кислотных почв. Lumbricus Tarrestris все еще присутствует в PH 5,4, Dendrobaena Octaedra на PH 4,3, а некоторые мегасколециды присутствуют в чрезвычайно кислых гумических почвах. PH почвы также может влиять на количество червей, которые входят в диапаузу . Чем более кислая почва, тем раньше черви входят в диапаузу и остаются в диапаузе самое длительное время при рН 6,4.
Земля дождевых червей охотятся многими видами птиц (например, Робинс, Скворцы , Молотки , Чайки , Вороны ), змеи, деревянные черепахи, млекопитающие (например , медведи , кабаны, лисы , ежигог , свиньи, мосы (например, медведи, кабаны , лиси [ 51 ] ) и беспозвоночные (например, муравьи , [ 52 ] плоские черви , грунтовые жуки и другие жуки , улитки , пауки и слизняки ). Землю из дождевых червей имеют много внутренних паразитов , в том числе простейшие , Platyhelminthes, клещи и нематоды ; черви Их можно найти в крови , семинальных везикулах , колом или кишечнике , или в их коконах (например, «Гистиостома Мурчи» в сочетания [ 53 ] ).
Активность дождевого червя анирует и смешивает почву и способствует минерализации питательных веществ и их поглощению растительностью. Некоторые виды дождевого червя выходят на поверхность и паститесь на более высоких концентрациях органического вещества, смешивая его с минеральной почвой. Поскольку высокий уровень смешивания органических веществ связан с плодородием почвы , обилие дождевых червей, как правило, считается полезным для фермеров и садовников. [ 54 ] [ 55 ] Давным -давно, когда 1881 год Чарльз Дарвин писал: «Можно сомневаться, есть ли много других животных, которые сыграли настолько важную роль в истории мира, как и эти скромные организованные существа». [ 56 ]

Кроме того, хотя, как следует из названия, основная среда обитания дождевых червей находится в почве, они не ограничены этой средой обитания. Брандлинговый червь Эйзения Фетида живет в разлагающейся растительной материи и навозе. Arctiosstrotus vancouverensis с острова Ванкувер и Олимпийского полуострова обычно встречается в распадных бревнах хвойных. Aprocrectodea limicola , sparganophilus spp. И несколько других находятся в грязи в ручьях. Некоторые виды являются древесными, [ 57 ] Некоторые водные и некоторые эвригалины (устойчивые к соленой воде) и прибрежный (живущий на морском берегу, например, Pontodrilus litoralis ). [ 58 ] Даже у почвенных видов у специальных мест обитания, таких как почвы, полученные из змеиного , имеют собственную фауну дождевого червя.
Вермикомпостирование органических «отходов» и добавление этого органического вещества к почве, предпочтительно в качестве поверхностной мульчи , обеспечит несколько видов дождевых червей с требованиями пищи и питательных веществ и создаст оптимальные условия температуры и влаги, которые будут стимулировать их активность.
Земные черви являются экологическими показателями здоровья почвы . Продяковые черви питаются распадающимся веществом в почве и анализируя содержимое их пищеварительных трактов, дает представление об общем состоянии почвы. Китовый кишечник накапливает химические вещества, в том числе тяжелые металлы, такие как кадмий , ртуть , цинк и медь . Размер популяции дождевых червей указывает на качество почвы, поскольку здоровая почва будет содержать большее количество дождевых червей. [ 59 ]
Воздействие на окружающую среду
Основные преимущества деятельности дождевого червя для фертильности почвы для сельского хозяйства могут быть обобщены как:
- Биологический : во многих почвах дождевые черви играют важную роль в превращении больших кусочков органического вещества в богатый гумус , тем самым улучшая плодородие почвы. Это достигается за счет действий червя по тяге под поверхностью, нанесенным органическим веществом, таким как падение листьев или навоз, либо для пищи, либо для подключения его норки. Оказавшись в норе, червь измельчит лист, частично переваривает его и смешивает с землей. Червь (см. Внизу справа внизу) может содержать на 40 процентов больше гумуса, чем в верхних 9 дюймах (230 мм) почвы, в которой живет червь. [ 60 ]

- Химическое вещество : в дополнение к мертвому органическому веществу , дождевой червя также принимает любые другие частицы почвы, которые достаточно маленькие, включая песчаные зерна вплоть до 1 ⁄ 20 дюйма (1,3 мм) - в его желудочке, где эти минутные фрагменты зернистого измельчения все размолот в тонкую пасту, которая затем расщепляется в кишечнике. Когда червь выделяет это в виде слепков, нанесенных на поверхность или глубже в почве, минералы и питательные вещества растений изменяются на доступную форму для использования растений. Исследования в Соединенных Штатах показывают, что свежие дождевые червя в пять раз богаче в доступном азоте , в семи раз богаче в доступных фосфатах и в 11 раз больше в доступном калие , чем окружающие верхние 6 дюймов (150 мм) почвы. В условиях, где гумус в изобилии, вес произведенных слепков может быть более 4,5 килограмма (9,9 фунта) на червя в год. [ 60 ]
- Физический : заново измельчание дождевого червя создает множество каналов через почву и имеет большое значение для поддержания структуры почвы , обеспечивая процессы аэрации и дренажа. [ 61 ] пермакультуры Соучредитель Билл Моллисон отмечает, что, скользя в их туннелях, дождевые черви «действуют как бесчисленная армия поршней, качающих воздух в почвы и из почв в 24-часовом цикле (быстрее ночью)». [ 62 ] Таким образом, дождевой червя не только создает проходы для воздуха и воды для прохождения почвы, но и модифицирует жизненно важный органический компонент, который делает почву здоровой (см. Биотурбацию ). Земля дождевых червей способствуют образованию богатых питательными веществами слепков (глобулы почвы, стабильные в слизи почвы), которые имеют высокую агрегацию почвы и плодородие почвы и качество. [ 60 ] В почвах подзола дождевые черви могут уничтожить характерный полосатый вид профиля почвы, смешивая органические (LFH), элювиальные (E) и верхние иллюальные (B) горизонты, чтобы создать один темный горизонт AP. [ 63 ] [ 64 ]
Земля дождевых червей ускоряют цикл питательных веществ в системе почвы за счет фрагментации и смешивания мусора растений-физическое шлифование и химическое пищеварение. [ 60 ] Существование дождевого червя не может быть воспринято как должное. Доктор We Shewell-Cooper наблюдала «огромные численные различия между соседними садами», а на популяции червя влияют множество факторов окружающей среды, на многие из которых могут влиять правильные методы управления со стороны садовника или фермера. [ 65 ]
Дарвин подсчитал, что пахотная земля содержит до 53 000 на акр (130 000/га) червей, но более недавние исследования дали показатели, свидетельствующие о том, что даже плохая почва может поддерживать 250 000 на акр (620 000/га), в то время как богатые плодородные фермерские угодья могут иметь до 1 750 000 на акр (4300 000/ха), что весом в земле. скот на его поверхности. Богородно органические популяции дождевых червей намного выше - в среднем 500 на квадратный метр (46/кв. Фут) и до 400 G2 [ сомнительно - обсудить ] - Таковы, что для нас 7 миллиардов из нас каждый человек, живой сегодня, имеет поддержку 7 миллионов дождевых червей. [ 66 ]
Способность разрушать органические материалы и выделять концентрированные питательные вещества делает дождевой червя функциональным участником в проектах восстановления. В ответ на нарушения экосистемы, некоторые участки использовали дождевые черви для приготовления почвы для возвращения местной флоры. Исследования со станции D'Ecologie Tropicale de Lamto утверждают, что дождевые черви положительно влияют на скорость макроагрегатного образования, важную особенность для структуры почвы. [ 67 ] Было также обнаружено, что стабильность агрегатов в ответ на воду улучшается при построении дождевыми червями. [ 67 ]
Хотя выбросы парниковых газов еще не полностью оцениваются, выбросы дождевых червей, вероятно, способствуют глобальному потеплению, особенно после того, как дождевые черви на высшем уровне увеличивают скорость углеродных циклов и распространяются людьми во многих новых географиях. [ 68 ]
Угрозы
Азотистые удобрения имеют тенденцию создавать кислые условия , которые являются смертельными для червей, а на поверхности часто обнаруживаются мертвые образцы после применения таких веществ, как ДДТ , сера извести и арсенат свинца . В Австралии изменения в методах сельского хозяйства, такие как применение суперфосфатов на пастбищах и переход от пастырского земледелия на пахотное земледелие, оказали разрушительное влияние на популяции гигантского дождевого червя Гиппсленда , что приводило к их классификации как охраняемого вида . Во всем мире некоторые популяции дождевых червей были опустошены отклонением от органической продукции и распылением синтетических удобрений и биоцидов, причем по меньшей мере три вида теперь перечислены как вымершие, но гораздо больше находятся под угрозой исчезновения. [ 69 ]
Экономическое воздействие

Различные виды червей используются в вермикультуре , практика кормления органических отходов до дождевых червей для разложения пищевых отходов. Обычно это Eisenia fetida (или ее близкая относительная Eisenia Andrei ) или червь с брендами, обычно известный как тигровый червь или красный вигглер. Они отличаются от почвенных дождевых червей. В тропиках африканский ночной тире eudrilus eugeniae [ 70 ] индийские синие периониксные экскаватус и используются .
Дождевые черви продаются по всему миру; Рынок значительный. По словам Дуга Колликутта, «в 1980 году 370 миллионов червей были экспортированы из Канады, с канадской экспортной стоимостью 13 миллионов долларов США и американской розничной стоимостью 54 миллиона долларов». [ 71 ]
Земные черви обеспечивают отличный источник белка для рыб, птицы и свиней, но также традиционно используются для потребления человеком. Noke - это кулинарный термин, используемый маори Новой Зеландии для обозначения дождевых червей, которые они рассматривают деликатесы для своих вождей.
Смотрите также
- Дрилосфера , часть почвы под влиянием выделений и отливок дождевого червя
- Образование овощной формы через действия червей , книга 1881 года Чарльза Дарвина
- Почвенная жизнь
- Вермикомпост
- Вермифильтер
- Вермифильтер туалет
- Червь очаровательно
Ссылки
- ^ Андерсон, Фрэнк; Джеймс, Самуил (июнь 2017 г.). «Эволюция дождевых червей» . BMC . Получено 3 февраля 2024 года .
- ^ Omodeo, Pietro (2000). «Эволюция и биогеография мегадрилов (Annelida, Clitellata)» . Итальянский журнал зоологии . 67–2 (2): 179–201. doi : 10.1080/1125000000009356313 . S2CID 86293273 .
- ^ Бонковски, Майкл; Гриффитс, Брайан С.; Ритц, Карл (ноябрь 2000 г.). «Предпочтения пищевых дождевых червей для почвенных грибов». Pedobiologia . 44–6 (6): 667. Bibcode : 20000pedob..44..666b . doi : 10.1078/s0031-4056 (04) 70080-3 .
- ^ Высокий, Клайв А.; Lofty, JR (1977). Биология дождевых червей . Лондон: Чепмен и Холл. п. 80. ISBN 0-412-14950-8 .
- ^ Эдвардс, Клайв А.; Lofty, JR (1977). Биология дождевых червей . Лондон: Чепмен и Холл. п. 19. ISBN 0-412-14950-8 .
- ^ Эдвардс, Клайв А.; Lofty, JR (1977). Биология дождевых червей . Лондон: Чепмен и Холл. с. Предисловие. ISBN 0-412-14950-8 .
- ^ Коулман, Дэвид С.; Кроссли, да; Хендрикс, Пол Ф. (2004). Основы экологии почвы . Амстердам; Бостон: Elsevier Academic Press. п. 170. ISBN 0-12-179726-0 .
- ^ Подпрыгнуть до: а беременный Кливленд П. Хикман -младший; Ларри С. Робертс; Фрэнсис М Хикман (1984). Интегрированные принципы зоологии (7 -е изд.). Times Mirror/Mosby College Publishing. п. 344 . ISBN 978-0-8016-2173-4 .
- ^ Blakemore 2012 , p. XL
- ^ Блейкмор, RJ; и др. (2007). «Megascolex (promegascocolex) Mekongianus Cognetti, 1922 - его степень, экология и ассигнование по Amynthas (Clitellata / Oligochaeta: Megascolecidae)» (PDF) . Брошюры Zoologica.
- ^ Edwards & Bohlen 1996 , p. 11
- ^ Подпрыгнуть до: а беременный в Sims & Gerard 1985 , с. 3–6.
- ^ Edwards & Bohlen 1996 , p. 3
- ^ Feldkamp, J. (1924). J. , » « Feldkamp
- ^ Edwards & Bohlen 1996 , p. 8-9.
- ^ Edwards & Bohlen 1996 , p. 1
- ^ Sims & Gerard 1985 , p. 8
- ^ Подпрыгнуть до: а беременный Edwards & Bohlen 1996 , p. 12
- ^ Edwards & Bohlen 1996 , p. 6
- ^ Myrmecofourmis (2018). «Познакомьтесь с брызганием дождевого червя» . YouTube. Архивировано из оригинала ( видео ) 2021-10-30.
- ^ «Эксперимент: сравнение скоростей двух размеров нервных волокон» . Backyardbrains . Получено 4 апреля 2015 года .
- ^ Дрюс, CD; Ланда, КБ; McFall, JL (1978). «Гигантское нервное волокно активность в неповрежденных, свободно движущихся дождевых червях» . Журнал экспериментальной биологии . 72 : 217–227. doi : 10.1242/jeb.72.1.217 . PMID 624897 .
- ^ Quillin, KJ (1999). «Кинематическое масштабирование локомоции гидростатическими животными: онтогенство перистальтического ползания по дождевым червям Lumbricus terrestris». Журнал экспериментальной биологии . 202 (6): 661–674. doi : 10.1242/jeb.202.6.661 . PMID 10021320 .
- ^ Подпрыгнуть до: а беременный «Земля-скрипутная система» . Кронодон . Получено 3 апреля 2015 года .
- ^ Elwood, RW (2011). "Боль и страдания в беспозвоночных?" Полем Ilar Journal . 52 (2): 175–84. doi : 10.1093/ilar.52.2.175 . PMID 21709310 .
- ^ Смит, JA (1991). «Вопрос о боли у беспозвоночных» . Ilar Journal . 33 (1–2): 25–31. doi : 10.1093/ilar.33.1-2.25 .
- ^ Röhlich, P.; Aros, B.; Virágh, Sz. «Прекрасная структура фоторецепторных клеток в дождевой червях, Lumbricus terrestris ». Журнал клеточных исследований и микроскопической анатомии . 104 (3): 345–357. Doi : 10.1007/bf00335687 . PMID 4193489 . S2CID 22771585 .
- ^ Edwards & Bohlen 1996 , p. 13
- ^ Либеке, Мануэль; Strittmatter, Николь; Ферн, Сара; Морган, А. Джон; Килл, Питер; Фухсер, Йенс; Уоллис, Дэвид; Palchykov, Vitalii; Робертсон, Джереми (2015-08-04). «Уникальные метаболиты защищают дождевые черви от растений полифенолов» . Природная связь . 6 : 7869. Bibcode : 2015natco ... 6.7869L . doi : 10.1038/ncomms8869 . PMC 4532835 . PMID 26241769 .
- ^ Edwards & Bohlen 1996 , с. 13–15.
- ^ Sims & Gerard 1985 , p. 10
- ^ Кливленд П. Хикман -младший; Ларри С. Робертс; Фрэнсис М Хикман (1984). Интегрированные принципы зоологии (7 -е изд.). Times Mirror/Mosby College Publishing. С. 344–345 . ISBN 978-0-8016-2173-4 .
- ^ Подпрыгнуть до: а беременный Farabee, HJ "Электронная система" . Архивировано из оригинала 30 июля 2012 года . Получено 29 июля 2012 года .
- ^ Кливленд П. Хикман -младший; Ларри С. Робертс; Фрэнсис М Хикман (1984). Интегрированные принципы зоологии (7 -е изд.). Times Mirror/Mosby College Publishing. С. 345–346 . ISBN 978-0-8016-2173-4 .
- ^ Домингес Дж., Айра М., Брейнхолт Дж.В., Стоянович М., Джеймс С.В., Перес-Лосада М. (2015). «Подземная эволюция: новые корни для старого дерева ламбрицидных дождевых червей» . Мол Филогенет. Эвол . 83 : 7–19. Bibcode : 2015molpe..83 .... 7d . doi : 10.1016/j.ympev.2014.10.024 . PMC 4766815 . PMID 25463017 .
- ^ Fernández R, Almodódódódóvar A, Novo M, Simancas B, Díaz Cosín DJ (2012). «Добавление сложности в комплекс: новое понимание филогении, диверсификации и происхождения партеногенеза в видах Aprocrectodea Caliginosa (Oligocidae, Lubricidae) Мол Филогенет. Эвол 64 (2): 368–7 Bibcode : 2012molpe..64..368f Doi : 10.1016/ j.ympev.2012.04.0 PMID 22542691
- ^ Cosín DJD, Novo M., Fernández R. (2011) Репродукция дождевых червей: половой отбор и партеногенез. В: Карака А. (ред.) Биология дождевых червей. Почвенная биология, том 24. Springer, Berlin, Heidelberg, с. 76ff. [1]
- ^ Ново М., Фернандес-Герра А., Гибабет Г. (2013). Полем Мол Биил Эвол . 30 (7): 1614–2 doi : 10.1093/ mst HDL : 10261/94159 . PMID 2356327 .
- ^ Бернштейн Х, Хопф Ф.А., Мичод Р.Е. (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения в области генетики. Тол. 24. С. 323–70. doi : 10.1016/s0065-2660 (08) 60012-7 . ISBN 978-0-12-017624-3 Полем PMID 3324702 .
- ^ Подпрыгнуть до: а беременный Hertel-Aas T, Ougton DH, Jaworska A, Brunburg G (ноябрь 2011 г.). «Индукция и восстановление ДНК -слома и окисленные на основе соматических и сперматогенных клеток из Eisenia ftida после воздействия ивингера» Мутагенез 26 (6): 783–9 Doi : 10.1093/ mutage/ ger0 21825113PMID
- ^ Chang WS, Tsai CW, Lin CC, Lin CH, Shen WC, Lin SS, Bau DT (2011). «Земные черви восстанавливают H2O2-индуцированные окислительные аддукты ДНК без удаления, индуцированных УФ-индуцированным димером пиримидина». In vivo . 25 (6): 977–81. PMID 22021692 .
- ^ Quillan, KJ (2000). «Онтогенетическое масштабирование закусочных сил в дождевом червях Lumbricus terrestris» . Журнал экспериментальной биологии . 203 (Pt 18): 2757–2770. doi : 10.1242/jeb.203.18.2757 . PMID 10952876 . Получено 4 апреля 2015 года .
- ^ Гейтс, GE (апрель 1949 г.). «Регенерация в дождевой червях, Эйзения Фотида (Savigny) 1826. I. Передняя регенерация» (PDF) . Биологический бюллетень . 96 (2): 129–139. doi : 10.2307/1538195 . JSTOR 1538195 . PMID 18120625 . Архивировано из оригинала (PDF) на 2007-04-01.
- ^ Гейтс, GE (1 января 1953 г.). «О регенеративной способности дождевых червей семейства Lumbricidae». Американский натуралист Мидленда . 50 (2): 414–419. doi : 10.2307/2422100 . JSTOR 2422100 .
- ^ "Invertebrata 20a предметы" . Архивировано из оригинала 2006-06-22 . Получено 2006-11-20 .
- ^ Fender & McKey-Fender (1990). Руководство по биологии почвы . Wiley-Interscience. ISBN 978-0-471-04551-9 .
- ^ Подпрыгнуть до: а беременный в Blakemore, RJ (2006) (март 2006 г.). «Пересмотренный ключ к семействам дождевых червей из Blakemore (2000) плюс обзоры Criodrilidae (включая Biwadrilidae) и Octochaetidae» (PDF) . Серия доступных для поиска текстов о биоразнообразии, экологии и систематике дождевого червя из различных регионов мира . Annelida.net . Получено 15 мая 2012 года .
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ «Землям» . Земля для дождевых червей.elte.hu . Получено 19 марта 2022 года .
- ^ [Ref Blakemore (2016) Космополитические дождевые черви]
- ^ ПРИМОННЫЕ ЗАМЕРКИ: Возобновление агропосистем (SA Fall, 1990 (V3N1)) Архивировали 2007-07-13 на машине Wayback
- ^ Гулд, Эдвин; Макшиа, Уильям; Гранд, Теодор (1993). «Функция звезды в носовой моле, condylura cristata». Журнал млекопитающих . 74 (1): 108–116. doi : 10.2307/1381909 . ISSN 0022-2372 . JSTOR 1381909 .
- ^ Dejean, A.; Schatx, B. (1999). «Захват добычи Psalidomyrmex Procerus (Formicidae; Ponerinae), специализированный хищник дождевых червей (Annelida)». Социобиология : 545–554. ISSN 0361-6525 .
- ^ Оливер, Джеймс Х. (1962). «Клещ паразита в коконах дождевых червей» . Журнал паразитологии . 48 (1): 120–123. doi : 10.2307/3275424 . ISSN 0022-3395 . JSTOR 3275424 . PMID 14481811 .
- ^ Департамент первичных отраслей NSW, как дождевые черви могут помочь вашей почве архивировано 2017-08-07 на машине Wayback
- ^ Галвестон округа Ассоциация мастера -садовника, выгодные в саду: #38 Переверны
- ^ Дарвин, Чарльз (1881). Образование овощной формы посредством действия червей, с наблюдениями за их привычками . Джон Мюррей . Найден в проекте Гутенберг Эткс Формирование растительной формы, Дарвином
- ^ Haum, Лоуренс; Шеной, Мегха; Захария, веселый; Борхес, Рене М. (май 2006 г.). «Сосуществование муравьев и древесного дождевого червя в мирмекофите индийских западных Гат: анти-пресс-эффект слизи дождевого червя» . Журнал тропической экологии . 22 (3): 341–344. doi : 10.1017/s0266467405003111 . ISSN 1469-7831 .
- ^ Blakemore, RJ (2007). «Происхождение и средства рассеивания космополитического Pontodrilus litoralis (Oligochaeta: Megascolecidae)» . Европейский журнал биологии почвы.
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Fründ, Heinz-Christian; Грэф, Ульферт; Tischer, Sabine (2011), Karaca, Ayten (ed.), «Продяковые черви как биоиндикаторы качества почвы» , Биология дождевых червей , почвенная биология, вып. 24, Берлин, Гейдельберг: Springer Berlin Heidelberg, с. 261–278, doi : 10.1007/978-3-642-14636-7_16 , ISBN 978-3-642-14635-0 Получено 2021-02-18
- ^ Подпрыгнуть до: а беременный в дюймовый Найл С. Брэди; Рэй Р. Вейл (2009). Элементы природы и свойств почв (3 -е изд.). Прентис Холл. ISBN 978-0-13-501433-2 .
- ^ Lines-Kelly, Ребекка (2021). «Как дождевые черви могут помочь вашей почве» . www.dpi.nsw.gov.au. Архивировано из оригинала 2021-10-08 . Получено 2021-11-30 .
- ^ Моллон, Билл, Пермакультура- Руководство по дизайнеру , Tagari Press, 1988
- ^ RT Fisher (январь 1928 г.). «Изменения почвы и сильвикультура в Гарвардском лесу» (PDF) . Экология . 9 (1): 6–11. Bibcode : 1928ecol .... 9 .... 6f . doi : 10.2307/1929537 . JSTOR 1929537 . Получено 2022-03-19 .
- ^ Лангмай, К.К. (1 февраля 1964 г.). Девственные подзолы Суо 44 (1): 34–3 doi : 4141/ cjss6-0 2022Марш
- ^ Купер, Шеуэлл; Почва, гумус и здоровье ISBN 978-0-583-12796-7
- ^ Blakemore, RJ (2017) (2017-02-12). «Природная статья, чтобы ознаменовать день рождения Чарльза Дарвина 12 февраля» . Вермекология.
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Подпрыгнуть до: а беременный Бланшарт, Эрик (1992-12-01). «Восстановление дождевых червей (мегасколециды) макроагрегатной структуры деструктурированной почвы саванны в полевых условиях». Почвенная биология и биохимия . 24 (12): 1587–1594. Bibcode : 1992sbibi..24.1587b . doi : 10.1016/0038-0717 (92) 90155-q .
- ^ Берк, Дэвид (26 декабря 2019 г.). «Сила корма дождевого червя и как это может повлиять на изменение климата» . CBC .
- ^ Blakemore, RJ (2018) (2018). «Критическое снижение дождевых червей от органического происхождения под интенсивным, гуминовым сельским хозяйством в чьей-ом» . Почвенные системы . 2 (2). Почвенные системы 2 (2): 33: 33. doi : 10.3390/почвы2020033 .
{{cite journal}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Blakemore, RJ (2015). «Экогасономический профиль культового вермикопостера -« африканский ночной », Eudrilus eugeniae (Kinberg, 1867)» . Африканские беспозвоночные 56: 527-548. Архивировано с оригинала 2016-10-22.
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Колликутт, Дуг. «Биология ночного гусеница (Lumbricus terrestris)» . Натуральный . Получено 5 июня 2022 года .
Работы цитируются
- Блакмор, Роберт Дж. (2012). Космополитические дождевые черви-экогаксономическое руководство по видам мирового переноса. (5 -е изд) . Йокогама, Япония: вермекология.
- Эдвардс, Клайв А.; Bohlen, PJ (1996). Биология и экология дождевых червей . Springer Science & Business Media. ISBN 978-0-412-56160-3 .
- Симс, Реджинальд Уильям; Джерард, Б. (1985). Земля дождевых червей: ключи и примечания для идентификации и изучения видов . Лондон: опубликовано для Лондонского общества Линни и ассоциации наук о солоноватых водах EJ Brill/DR. В. Бэкхайс.
Дальнейшее чтение
- Эдвардс, Клайв А. (ред.) Экология дождевого червя . Boca Raton: CRC Press, 2004. Второе пересмотренное издание. ISBN 0-8493-1819-X
- Ли, Кенет Э. Перелезные черви: их экология и отношения с почвами и землепользой . Академическая пресса. Сидней, 1985. ISBN 0-12-440860-5
- Стюарт, Эми. Земля двигалась: на замечательных достижениях дождевых червей . Чапел -Хилл, Северная Каролина: Алгонкинские книги, 2004. ISBN 1-56512-337-9
Внешние ссылки
 СМИ, связанные с дождевыми червями в Wikimedia Commons
СМИ, связанные с дождевыми червями в Wikimedia Commons  Данные, связанные с Lumbricina в Wikispecies
Данные, связанные с Lumbricina в Wikispecies - Чисхолм, Хью , изд. (1911). . Encyclopædia Britannica (11 -е изд.). Издательство Кембриджского университета.