Белый носорог — самый крупный из ныне живущих непарнокопытных.
Периссодактилия ( / p ə ˌ r ɪ so oʊ ˈ d æ k t ɪ l ə / , от древнегреческого περισσος , perissós 'нечетный' и δακτιλος , dáktylos 'палец, нога' [3] ) — отряд копытных . Отряд включает около 17 современных видов, разделенных на три семейства : Equidae ( лошади , ослы и зебры ), Rhinocerotidae ( носороги ) и Tapiridae ( тапиры ). Обычно у них уменьшены несущие вес пальцы ног до трех или одного из пяти первоначальных пальцев, хотя у тапиров на передних лапах сохраняются четыре пальца. Пальцы, не несущие нагрузки, либо присутствуют, либо отсутствуют, рудиментарны , либо расположены сзади. Напротив, парнокопытные (парнокопытные) несут большую часть своего веса поровну на четырех или двух (четное количество) из пяти пальцев: третьем и четвертом пальцах. Еще одно различие между ними заключается в том, что непарнокопытные переваривают растительную целлюлозу в своем кишечнике , а не в одной или нескольких камерах желудка , как это делают парнокопытные, за исключением Suina .
В прошлом отряд был значительно более разнообразным: известные вымершие группы включали бронтотериев , палеотериев , халикотериев и парацератеров , причем парацератеры включали крупнейших известных наземных млекопитающих, когда-либо существовавших.
Несмотря на их совершенно разный внешний вид, в 19 веке зоолог Ричард Оуэн признал их родственными семьями , который также придумал название отряда.
Самыми крупными непарнокопытными являются носороги, а вымерший Paraceratherium , безрогий носорог из олигоцена , считается одним из крупнейших наземных млекопитающих всех времен. [4] С другой стороны, один из первых представителей отряда, доисторический конь Эогипп , имел высоту в холке всего от 30 до 60 см (от 12 до 24 дюймов). [5] За исключением карликовых разновидностей домашней лошади и осла, современные непарнокопытные достигают длины тела 180–420 см (71–165 дюймов) и веса от 150 до 4500 кг (от 330 до 9920 фунтов). [6] В то время как у носорогов редкая шерсть и толстый эпидермис , у тапиров и лошадей густая короткая шерсть. Большинство видов серые или коричневые, хотя зебры и молодые тапиры полосатые.
Семь фигур, изображающих кости, кровеносные сосуды, связки и артерии копыта и пясти.
Основные оси как передних, так и задних стоп проходят через третий палец, который всегда самый большой. Остальные пальцы ног уменьшились в разной степени. У тапиров, приспособленных к ходьбе по мягкой земле, четыре пальца на передних ногах и три на задних. У современных носорогов по три пальца как на передних, так и на задних лапах. У современных лошадей только один палец; однако их ступни снабжены копытами, почти полностью закрывающими палец. У носорогов и тапиров, напротив, копыта покрывают только передний край пальцев, а подошва мягкая.
У копытных есть поза, требующая стоять на кончиках пальцев. Лошадиные копытные, имеющие только один палец или копыта, имеют пониженную подвижность конечностей, что позволяет им развивать более высокую скорость бега и ловкость.Различия в строении и физиологии конечностей между копытными и другими млекопитающими можно увидеть в форме плечевой кости . Например, зачастую более короткие и толстые кости принадлежат самым крупным и тяжелым копытным, таким как носорог . [7]
Количество и форма зубов варьируются в зависимости от диеты. Резцы могут быть очень маленькими или вовсе и клыки отсутствовать, как у двух африканских видов носорогов. У лошадей клыки обычно есть только у самцов. Форма поверхности и высота коренных зубов во многом зависят от того, составляют ли основной компонент их рациона мягкие листья или жесткая трава. На каждой половине челюсти имеется по три-четыре щечных зуба, поэтому зубная формула непарнокопытных: 0-3. 0-1. 2-4. 3 1-3 . 1 . 2-4. 3 ×2 = 30-44
Гортанный мешочек , небольшой выступ слуховой трубы , который дренирует среднее ухо , является характерной особенностью Perissodactyla. [10] Гортанный мешок вызывает особую озабоченность в лошадей ветеринарной практике из-за его частого поражения при некоторых серьезных инфекциях. Аспергиллез (заражение плесенью Aspergillus ) гортанного мешка (также называемый микозом гортанного мешка ) может вызвать серьезное повреждение тканей мешка, а также окружающих структур, включая важные черепные нервы (нервы IX-XII: языкоглоточный , блуждающий , добавочный и Подъязычные нервы [10] ) и внутренняя сонная артерия . [10] Удушение ( инфекция Streptococcus equi equi ) — высокозаразная респираторная инфекция лошадей. [11] [12] это может привести к скоплению гноя в гортанном мешке; [13] лошади с S. equi equi, колонизирующим их гортанный мешочек, могут продолжать периодически выделять бактерии в течение нескольких месяцев. [14] и на это время их следует изолировать от других лошадей, чтобы предотвратить передачу инфекции. Из-за периодического характера выделения S. equi equi , преждевременное повторное введение инфицированной лошади может привести к риску заражения других лошадей, даже если лошадь, выделяющая S. equi, выглядит здоровой и ранее могла сдать отрицательные образцы. Функцию гортанного мешка определить сложно, но сейчас считается, что он играет роль в охлаждении крови во внутренней сонной артерии перед ее попаданием в мозг. [10]
Все непарнокопытные являются ферментаторами задней кишки. В отличие от жвачных животных , ферментеры задней кишки сохраняют переваренную пищу, покинувшую желудок , в расширенной слепой кишке , где пища переваривается микробами. отсутствует Желчный пузырь . Желудок непарнокопытных устроен просто, а у лошадей слепая кишка вмещает до 90 л (24 галлона США). очень Тонкая кишка длинная, у лошадей достигает 26 м (85 футов). Извлечение питательных веществ из пищи относительно неэффективно, что, вероятно, объясняет, почему непарнокопытные не имеют маленьких размеров; Потребность в питательных веществах на единицу массы тела у крупных животных ниже, поскольку у них соотношение площади поверхности к объему меньше .
В отличие от парнокопытных, у непарнокопытных отсутствует сонная сеть . [15] теплообмен, снижающий зависимость температуры мозга от температуры тела. В результате непарнокопытные обладают ограниченной гибкостью терморегуляции по сравнению с парнокопытными, что ограничивает их местообитаниями с низкой сезонностью и богатыми пищей и водой, такими как тропические леса. Напротив, парнокопытные обитают в широком диапазоне образов жизни — от Полярного круга до пустынь и тропических саванн. [15]
Ограничение среды обитания и браконьерство угрожают выживанию большинства видов носорогов, включая индийского носорога. показанного здесь
Большинство современных видов непарнокопытных занимают небольшую часть своего первоначального ареала. Члены этой группы сейчас встречаются только в Центральной и Южной Америке, восточной и южной Африке, а также в центральной, южной и юго-восточной Азии. [16] В период пика существования непарнокопытных, от эоцена до олигоцена , непарнокопытные были распространены на большей части земного шара, за исключением Австралии и Антарктиды. Лошади и тапиры прибыли в Южную Америку после образования Панамского перешейка около 3 миллионов лет назад в плиоцене . Их североамериканские собратья вымерли около 10 000 лет назад, и остался только тапир Бэрда , ареал которого простирается до территории нынешней южной Мексики. Тарпаны были уничтожены в Европе XIX века. Охота и разрушение среды обитания превратили выжившие виды непарнокопытных в фрагментированные популяции. Напротив, одомашненные лошади и ослы получили распространение по всему миру, а дикие животные обоих видов теперь также встречаются в регионах за пределами их первоначального ареала, например, в Австралии.
Непарнокопытные обитают в разных средах обитания, что приводит к разному образу жизни. Тапиры одиночки и населяют преимущественно влажные тропические леса. Носороги, как правило, живут поодиночке в довольно сухих саваннах, а в Азии — на влажных болотах или в лесных районах. Лошади населяют открытые территории, такие как луга, степи или полупустыни, и живут группами. Непарнокопытные — исключительно травоядные животные, питающиеся в той или иной степени травой, листьями и другими частями растений. Часто проводят различие между питающимися преимущественно травой ( белые носороги , лошади ) и питающимися листьями (тапиры, другие носороги).
Непарнокопытные характеризуются длительным периодом беременности и небольшим размером помета, обычно принося одного детеныша. Период беременности составляет 330–500 дней, самый длинный у носорогов. Новорожденные непарнокопытные являются скороспелыми , то есть потомство рождается уже вполне самостоятельным: например, [17] молодые лошади могут начать следовать за матерью уже через несколько часов. [18] Молодняк выкармливают в течение относительно долгого времени, часто до второго года жизни, при этом носороги достигают половой зрелости примерно в восемь или десять лет, а лошади и тапиры созревают примерно в возрасте от двух до четырех лет. [19] [20] Непарнокопытные долгожители: некоторые виды, например носороги, в неволе достигают возраста почти 50 лет. [21]
Традиционно непарнокопытных относили к другим млекопитающим, таким как парнокопытные , даманы , слоны и другие «копытные». Тесное семейное родство с даманами подозревалось на основании сходства в строении уха и ходе сонной артерии .
Однако молекулярно-генетические исследования показали, что копытные полифилетичны , а это означает, что в некоторых случаях сходство является результатом конвергентной эволюции, а не общего происхождения . Слоны и даманы теперь считаются принадлежащими к Afrotheria , поэтому не являются близкими родственниками непарнокопытных. Они, в свою очередь, входят в Laurasiatheria , суперотряд, возникший на бывшем суперконтиненте Лавразия . Молекулярно-генетические данные позволяют предположить, что раздвоенные парнокопытные (содержащие китообразных в виде глубоко расположенного субклада) являются сестринским таксоном Perissodactyla; вместе эти две группы образуют Euungulata . [22] Еще дальше находятся летучие мыши (Chiroptera) и Ferae (общий таксон хищных животных Carnivora и ящеров Pholidota ). [23] В дискредитированном альтернативном сценарии существует тесная связь между непарнокопытными, хищниками и летучими мышами, причем это собрание включает Pegasoferae . [24]
Внутренняя систематика Euungulata по Welker et al. 2015 год [25]
Согласно исследованиям, опубликованным в марте 2015 года, непарнокопытные находятся в тесном семейном родстве, по крайней мере, с некоторыми из так называемых Meridiungulata — весьма разнообразной группы млекопитающих, обитающих от палеоцена до плейстоцена в Южной Америке, систематическое единство которых во многом необъяснимо. Некоторые из них были классифицированы на основе их палеогеографического распространения. Однако тесная связь с непарнокопытными может быть установлена путем секвенирования белков и сравнения с ископаемым коллагеном из остатков филогенетически молодых представителей Meridiungulata (в частности, Macrauchenia из Litopterna и Toxodon из Notoungulata ). Обе группы родства, непарнокопытные и Litopterna-Notoungulata, теперь входят в таксон более высокого уровня Panperissodactyla . Эта группа родства входит в состав Euungulata, к которому относятся также парнокопытные и киты (Artiodactyla). Отделение группы Litopterna-Notoungulata от непарнокопытных, вероятно, произошло еще до Мел-палеогеновое вымирание . «Кондилартов», вероятно, можно считать отправной точкой развития двух групп, так как они представляют собой разнородную группу примитивных копытных, населявших в палеогене преимущественно северное полушарие . [25] [26]
Непарнокопытные (Perissodactyla) включают три современных семейства, насчитывающих около 17 видов; однако у лошадей точное количество до сих пор остается спорным. Носороги и тапиры более тесно связаны друг с другом, чем с лошадьми. Отделение лошадей от других непарнокопытных произошло по данным молекулярно-генетического анализа в палеоцене около 56 миллионов лет назад, тогда как носороги и тапиры отделились в нижнем-среднем эоцене , около 47 миллионов лет назад.
Внутренние взаимоотношения современных Perissodactyla [27] [28] [29]
Существует множество окаменелостей непарнокопытных многовариантной формы. К основным направлениям развития относятся следующие группы:
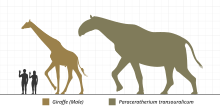
Масштабная диаграмма парацератериида Paraceratherium , одного из крупнейших когда-либо существовавших наземных млекопитающих. Brontotherioidea были одними из самых ранних известных крупных млекопитающих, состоящих из семейств Brontotheriidae (синоним Titanotheriidae), наиболее известным представителем которых был Megacerops и более базальное семейство Lambdotheriidae . На поздней стадии их развития обычно характеризовали костный рог на переходе от носа к лобной кости и плоские коренные зубы, пригодные для пережевывания мягкой растительной пищи. Brontotheroidea, обитавшие почти исключительно в Северной Америке и Азии, вымерли в начале верхнего эоцена .
Equoidea ( лошади ) также развилась в эоцене. Palaeotheriidae известны главным образом из Европы; их самый известный представитель — Eohippus , вымерший в олигоцене . Напротив, семейство лошадей (Equidae) процветало и распространялось. Со временем в этой группе наблюдалось уменьшение количества пальцев на ногах, удлинение конечностей и постепенное приспособление зубов к поеданию жесткой травы.
Rhinocerotoidea (родственники носорогов) включали большое разнообразие форм от эоцена до олигоцена, в том числе листофагов размером с собаку, полуводных животных, а также огромных животных с длинной шеей. Лишь у некоторых были рога на носу. Amynodontidae были водными животными, похожими на гиппопотамов. У Hyracodontidae развились длинные конечности и длинные шеи, которые были наиболее выражены у Paraceratherium (ранее известного как Baluchitherium или Indricotherium ), второго по величине известного наземного млекопитающего, когда-либо жившего на земле (после Palaeoloxodon namadicus) . [30] ). Носороги (Rhinocerotidae) появились в среднем эоцене; пять видов сохранились до наших дней.
Наибольшего разнообразия Tapiroidea достигла в эоцене, когда несколько семейств обитали в Евразии и Северной Америке. Они сохранили примитивное телосложение и отличались развитым туловищем. К вымершим семействам этой группы относятся Helaletidae . [ нужна ссылка ]
Несколько групп млекопитающих, традиционно классифицируемых как кондиларты , которые долгое время считались таксоном мусорной корзины , такие как гиопсодонтиды и фенакодонтиды , теперь считаются частью совокупности непарнокопытных. Фенакодонтиды, по-видимому, являются стеблево-периссодактилями, тогда как гиопсодонтиды тесно связаны с лошадьми и бронтотериями, несмотря на их более примитивный общий вид.
Демостилия [31] и Anthracobunidae традиционно относили к афротериям , но на самом деле они могут представлять собой стеблевые непарнокопытные. Это ранняя линия млекопитающих, которые пришли в воду и распространились по полуводным и полностью морским нишам в океане Тетис и северной части Тихого океана . Однако более поздние исследования показали, что, хотя антракобуниды являются явными непарнокопытными, десмостилии имеют достаточно смешанные признаки, чтобы предположить, что их положение среди афротерий не исключено. [32]
Взаимоотношения внутри большой группы непарнокопытных до конца не изучены. Первоначально, после создания Ричардом Оуэном в 1848 году «Непарнокопытных», нынешние представители считались равными по рангу. В первой половине 20 века началась более систематическая дифференциация непарнокопытных, основанная на рассмотрении ископаемых форм, и их объединили в два крупных подотряда: Hippomorpha и Ceratomorpha. Гиппоморфы включают сегодняшних лошадей и их вымерших представителей ( Equoidea ); Ceratomorpha состоит из тапиров и носорогов, а также их вымерших представителей ( Tapiroidea и Rhinocerotoidea ). [41] Названия Hippomorpha и Ceratomorpha были введены в 1937 году Хорасом Элмером Вудом в ответ на критику названия «Solidungula», предложенного им тремя годами ранее. Он был основан на группе лошадей и тридактилов, а также на комплексе носорог/тапир. [42] [43] Вымершие бронтотерииды также были отнесены к группе Hippomorpha и, следовательно, имеют близкое родство с лошадьми. Некоторые исследователи принимают это отнесение из-за схожих особенностей зубов, но существует также мнение, что очень базальное положение у непарнокопытных помещает их скорее в группу Titanotheriomorpha . [40] [44]
Первоначально Chalicotheriidae рассматривались как представители Hippomorpha и были представлены как таковые в 1941 году. Уильям Берриман Скотт считал, что, как непарнокопытные с когтями, они принадлежат к новому подотряду Ancylopoda (где Ceratomorpha и Hippomorpha как непарнокопытные были объединены в группа Chelopoda). [45] Термин Ancylopoda, придуманный Эдвардом Дринкером Коупом в 1889 году, был установлен для халикотериев. Однако дальнейшие морфологические исследования 1960-х годов показали среднее положение Ancylopoda между Hippomorpha и Ceratomorpha. Леонард Бертон Радинский считал все три основные группы непарнокопытных равными, основываясь на чрезвычайно длительном и независимом филогенетическом развитии трех линий. [46] В 1980-х годах Джереми Дж. Хукер увидел общее сходство между Ancylopoda и Ceratomorpha, основанное на зубном ряде, особенно у самых ранних представителей, что привело к объединению в 1984 году двух представлений во временный отряд Tapiromorpha . В то же время он расширил Ancylopoda, включив в него Lophiodontidae . Название «Tapiromorpha» восходит к Эрнсту Геккелю, который придумал его в 1873 году, но долгое время оно считалось синонимом Ceratomorpha, поскольку Вуд не рассматривал его в 1937 году, когда были названы Ceratomorpha, поскольку в прошлом этот термин использовался совершенно по-другому. [47] Также в 1984 году Роберт М. Шох использовал концептуально схожий термин Moropomorpha, который сегодня применяется как синоним Tapiromorpha. [48] В состав Tapiromorpha входят ныне вымершие Isectolophidae, сестринская группа группы Ancylopoda-Ceratomorpha и, таким образом, самые примитивные члены этого комплекса родства. [44] [37]
Эволюционное развитие Perissodactyla хорошо задокументировано в летописи окаменелостей. Многочисленные находки свидетельствуют об адаптивной радиации этой группы, которая когда-то была гораздо более разнообразной и широко рассредоточенной. Радинскую из позднего палеоцена Восточной Азии часто считают одним из древнейших близких родственников копытных. [49] Его череп размером 8 см, вероятно, принадлежал очень маленькому и примитивному животному с π-образным рисунком коронки на эмали задних коренных зубов, похожим на таковой у непарнокопытных и их родственников, особенно носорогов. [50] Находки камбайтерия и калитерия в камбейских сланцах на западе Индии указывают на их происхождение из Азии, датируемое нижним эоценом примерно 54,5 миллиона лет назад. [51] [52] Их зубы также имеют сходство с Радинской , а также с кладой Тетитерия . [53] [54] На близкое родство с Tethytheria указывает также седловидная конфигурация ладьевидных суставов и меаксонное строение передних и задних лап. [ нужна ссылка ] Однако эта конструкция отличается от конструкции Cambaytherium , что указывает на то, что на самом деле он является членом сестринской группы. Предки Perissodactyla, возможно, прибыли через островной мост с афро-арабской суши на Индийский субконтинент, когда он дрейфовал на север, в сторону Азии. [55] Исследование камбайтерия предполагает его происхождение из Индии до или недалеко от ее столкновения с Азией. [56]
Сопоставление гиопсодонтид и фенакодонтид с Perissodactyla в целом предполагает более древнее лавразийское происхождение и распространение клады, рассредоточенной по северным континентам уже в раннем палеоцене. Эти формы уже имеют достаточно развитую молярную морфологию, без промежуточных форм, свидетельствующих о ходе ее развития. [57] Тесная связь между меридионгулятыми млекопитающими и непарнокопытными, в частности, представляет интерес, поскольку последние появились в Южной Америке вскоре после события K – T , что подразумевает быструю экологическую радиацию и расселение после массового вымирания. [58]
Эогиппус , ранний родственник лошади, является одним из старейших известных непарнокопытных.
Perissodactyla появились относительно внезапно в начале нижнего палеоцена около 63 миллионов лет назад как в Северной Америке, так и в Азии, в виде фенакодонтид и гиопсодонтид . Самые старые находки из дошедшей до нас группы происходят, среди других источников, от Сифригиппуса , предка лошадей из семейства Уиллсвуд в северо-западном Вайоминге . [59] [60] Вскоре после этого в составе семейства Газиев в Белуджистане появились далекие предки тапиров , такие как Ganderalophus , а также Litolophus из линии Chalicotheriidae или Eotitanops из группы brontotheriidae. [61] [62] Первоначально представители разных линий выглядели очень похоже: с выгнутой спиной и, как правило, с четырьмя пальцами на передних и тремя на задних лапах. Эогиппус , считающийся представителем семейства лошадиных, внешне напоминал Гирачия , первого представителя линии носорогов и тапиров. [63] Все они были небольшими по сравнению с более поздними формами и жили в лесах, питаясь фруктами и листвой. Первыми представителями мегафауны , появившимися были бронтотерии в среднем и верхнем эоцене, . Мегацеропс , известный из Северной Америки, достигал высоты в холке 2,5 м (8 футов 2 дюйма) и мог весить чуть более 3 метрических тонн (3,3 коротких тонны). Упадок бронтотериев в конце эоцена связан с конкуренцией, возникшей из-за появления более успешных травоядных. [33] [64]
Более успешные линии непарнокопытных появились в конце эоцена, когда густые джунгли уступили место степи, например носороги-халикотерииды и их ближайшие родственники; их развитие также началось с очень мелких форм. Парацератерий , одно из крупнейших млекопитающих, когда-либо живших на Земле, эволюционировал в эту эпоху. [65] [66] Они весили до 20 метрических тонн (22 коротких тонны) и жили на протяжении всего олигоцена в Евразии. Около 20 миллионов лет назад, в начале миоцена, непарнокопытные впервые достигли Африки, когда она соединилась с Евразией из-за закрытия океана Тетис . новые животные, такие как мамонты , Однако по той же причине на территории древних поселений непарнокопытных проникли и создав конкуренцию, которая привела к исчезновению некоторых их линий. Возникновение жвачных животных , занимавших схожие экологические ниши и имевших гораздо более эффективную пищеварительную систему, также связано со снижением разнообразия непарнокопытных. Существенной причиной сокращения численности непарнокопытных стало изменение климата в миоцене, которое привело к более прохладному и сухому климату, сопровождавшемуся распространением открытых ландшафтов. Однако некоторые линии процветали, например, лошади и носороги; анатомические приспособления позволили им потреблять более жесткую травяную пищу. Это привело к появлению открытых форм суши, которые доминировали во вновь созданных ландшафтах. С появлением Панамский перешеек в плиоцене непарнокопытные и другие представители мегафауны получили доступ к одному из последних обитаемых континентов: Южной Америке. [67] [68] Однако многие непарнокопытные вымерли в конце ледникового периода, в том числе американские лошади и эласмотерии . Вопрос о том, стала ли причиной исчезновения мегафауны ледникового периода чрезмерная охота со стороны людей (гипотеза чрезмерного истребления), изменение климата или сочетание обоих факторов, остается спорным. [33]
В 1758 году в своей основополагающей работе Systema Naturae Линней ( 1707–1778) классифицировал лошадей ( Equus ) вместе с гиппопотамами ( Hippopotamus ). В то время в эту категорию входили и тапиры ( Tapirus ), точнее равнинный или южноамериканский тапир ( Tapirus terrestus ), единственный известный тогда в Европе тапир. Линней отнес этого тапира к Hippopotamus terrestris и отнес оба рода к группе Belluae («звери»). Он объединил носорогов с Glires , группой, состоящей теперь из зайцеобразных и грызунов . Матюрен Жак Бриссон (1723–1806) впервые разделил тапиров и бегемотов в 1762 году, введя понятие le Tapir . Он также отделил носорогов от грызунов, но не объединил три семейства, ныне известные как непарнокопытные. При переходе к XIX веку отдельные роды непарнокопытных были связаны с различными другими группами, например с хоботными и парнокопытными копытными . В 1795 году Этьен Жоффруа Сен-Илер (1772–1844) и Жорж Кювье. (1769–1832) ввёл термин «толстокожие» ( Pachydermata ), включив в него не только носорогов и слонов, но также бегемотов, свиней, пекариев , тапиров и даманов. [41] [44] [69] [70] Лошадей по-прежнему считали отдельной от других млекопитающих группой, и их часто называли Solidungula или Solipèdes , что означает «однокопытное животное». [71] [72]
В 1861 году Анри Мари Дюкроте де Бленвиль (1777–1850) классифицировал копытных по строению их ног, отличая животных с четным числом пальцев от животных с нечетным числом. Он отнес лошадей как сплошных копытных к тапирам и носорогам как к многоногим животным и назвал их всех вместе onguligrades à doigtsimpairs , приближаясь к понятию о непарнокопытных как о систематической единице. Ричард Оуэн (1804–1892) процитировал Бленвилля в своем исследовании ископаемых млекопитающих острова Уайт и ввел название Perissodactyla . [41] [44]
В 1884 году Отниэль Чарльз Марш (1831–1899) предложил концепцию Mesaxonia , которую он использовал для обозначения так называемых непарнопалых копытных, включая их вымерших родственников, но явно исключая дамана. Mesaxonia теперь считается синонимом Perissodactyla , но иногда его также использовали для настоящих непарнокопытных как подкатегории (носорогов, лошадей, тапиров), тогда как Perissodactyla обозначала весь отряд, включая дамана. Предположение о том, что даманы были Perissodactyla, сохранялось вплоть до 20 века. [73] Только с появлением молекулярно-генетических методов исследования было признано, что даман был близким родственником не непарнокопытных, а скорее слонов и ламантинов. [22] [74]
Домашняя лошадь и осел играют важную роль в истории человечества, особенно как транспортные, рабочие и вьючные животные. Одомашнивание . обоих видов началось за несколько тысячелетий до нашей эры Из-за автомобилизации сельского хозяйства и распространения автомобильного движения такое использование резко сократилось в промышленных странах Запада; езда обычно рассматривается скорее как хобби или спорт. Однако в менее развитых регионах мира традиционное использование этих животных все еще широко распространено. В меньшей степени лошадей и ослов разводят ради мяса и молока.
Напротив, существование в дикой природе почти всех других видов непарнокопытных резко сократилось из-за охоты и разрушения среды обитания. Квагга лошадь вымерла, а Пржевальского когда-то была истреблена в дикой природе.
^ Балленджер, Лиз; Майерс, Фил. «Rhinocerotidae (носороги)». Сеть разнообразия животных. Получено 27 марта 2021 г.. Балленджер, Лиз; Майерс, Фил. «Rhinocerotidae (носороги)». Сеть разнообразия животных. Получено 27 марта 2021 г.
^ Перейти обратно: а б Дэн Граур; Маноло Гуи; Лоран Дюре (1997). «Эволюционное сходство отряда Perissodactyla и филогенетический статус надпорядковых таксонов Ungulata и Altungulata. Молекулярная филогенетика и эволюция». Молекулярная филогенетика и эволюция . 7 (2): 195–200. дои : 10.1006/mpev.1996.0391 . ПМИД 9126561 .
^ Марио А. Коззуоль; Камила Л. Клозато; Элизет К. Оланда; Флавио Х.Г. Родригес; Сэмюэл Ниноу; Бенуа де Туази; Родриго Редондо; АФ Фабрисио Р. Сантос (2013). «Новый вид тапира Амазонки» . Журнал маммологии . 94 (6): с 1331 по 1345. doi : 10.1644/12-MAMM-A-169.1 .
^ Гонг, Ю.; Ван, Ю.; и др. (декабрь 2019 г.). «Диетические адаптации и палеоэкология Lophialetidae (Mammalia, Tapiroidea) из эоцена бассейна Эрлиан, Китай: объединенные данные мезоодежды и анализа стабильных изотопов». Палеонтология . 63 (4): 547–564. дои : 10.1111/пала.12471 . S2CID 214053761 .
^ Перейти обратно: а б с Джордж Гейлорд (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории : 252–258.
^ Гораций Элмер Вуд (1934). «Ревизия Hyrachyidae». Бюллетень Американского музея естественной истории : 181–295.
^ Перейти обратно: а б с д Роберт М. Шох (1989). «Краткий исторический обзор классификации периссодактилей». Эволюция непарнокопытных . Издательство Оксфордского университета. стр. 13–23.
^ Уильям Берриман Скотт (1941). «Часть V: Периссодактилия». Фауна млекопитающих олигоцена Уайт-Ривер. Труды Американского философского общества . Новая серия. 28 (5): 747–964. дои : 10.2307/1005518 . JSTOR 1005518 .
^ Леонард Б. Радинский (1964). « Paleomoropus , новый раннеэоценовый халикотерий (Mammalia, Perissodactyla) и ревизия эоценовых халикотериев». Новитаты Американского музея : 1–28.
^ Джей Джей Хукер (1984). «Примитивный цератоморф (Perissodactyla, Mammalia) из раннего третичного периода Европы». Зоологический журнал Линнеевского общества . 82 (1–2): 229–244. дои : 10.1111/j.1096-3642.1984.tb00545.x .
^ Роберт Милтон Шох (1984). «Два необычных экземпляра хелалетов Йельского музея Пибоди в коллекциях и некоторые комментарии о происхождении тапирид (Perissodactyla, Mammalia)». Музей Пибоди, Йельский университет: 1–20. {{cite journal}}: Для цитирования журнала требуется |journal= ( помощь )
^ Малкольм К. Маккенна; Чоу Минна; Суйин Тин; Ло Чжэси (1989). « Radinskya yupingae , непарнокопытное млекопитающее из позднего палеоцена Китая». Эволюция непарнокопытных . Издательство Оксфордского университета. стр. 24–36.
^ Кеннет Д. Роуз (2006). Начало эпохи млекопитающих . Издательство Университета Джонса Хопкинса. стр. 242–267.
^ Кеннет Д. Роуз; Тьерри Смит; Раджендра С. Рана; Ашок Сахни; Х. Сингх; А. Питер Миссиен (2006). «Раннеэоценовый (ипрский) комплекс континентальных позвоночных из Индии с описанием нового антракобунида (Mammalia, tethytheria)». Журнал палеонтологии позвоночных . 26 : 219–225. doi : 10.1671/0272-4634(2006)26[219:eeycva]2.0.co;2 . S2CID 86206151 .
^ Питер Миссиен; Грегг Ф. Ганнелл; Филип Д. Джинджерич (2011). «Новые бронтотерииды (Mammalia, Perissodactyla) из раннего и среднего эоцена Пакистана, имеющие значение для палеобиогеографии млекопитающих». Журнал палеонтологии . 85 (4): 665–677. дои : 10.1666/10-087.1 . S2CID 129473193 .
^ Главач, Керстин; Эрфурт, Йорг (2007). «Морфология зубов и стратиграфическое распространение Hyrachyus minimus » ( Perissodactyla, Mammalia) в эоценовых слоях Гейзельталя . Ежегодник Галле по геологическим наукам . Приложение 23: 161–173.
^ Кристин Дженис (2008). «Эволюционная история поиска и выпаса копытных». У Иэна Дж. Гордона; Герберт Х. Т. Принс (ред.). Экология просмотра и выпаса . Экологические исследования. Том. 195. Спрингер. стр. 21–45. дои : 10.1007/978-3-540-72422-3_2 . ISBN 978-3-540-72421-6 .
^ Бентон, Майкл Дж. (1997). Палеонтология позвоночных . Лондон: Чепмен и Холл. п. 343. ИСБН 0-412-73810-4 .
^ Этьен Жоффруа Сен-Илер; Жорж Кювье (1795). «Память о новом разделении млекопитающих и о принципах, которые должны послужить основой в этом виде работы». Энциклопедический магазин : 164–190.
^ Жорж Кювье (1817). «Царство животных распределено в соответствии с его организацией и служит основой естественной истории животных». Введение в сравнительную анатомию . 1 :1–540.
^ Иоганн Фридрих Блюменбах (1779). Справочник по естественной истории . стр. 168–448.
^ Дональд Р. Протеро; Роберт М. Шох (1989). «Классификация непарнокопытных». Эволюция непарнокопытных . Издательство Оксфордского университета. стр. 530–537.
Мартин С. Фишер: Месаксония (Perissodactyla) Perissodactyla. В: Вильфрид Вестхайде, Рейнхард Ригер (ред.): Систематическая зоология. Часть 2: Вихрь или краниоты . Spektrum Akademischer Verlag, Гейдельберг и Берлин, 2004 г., стр. 646–655, ISBN 3-8274-0307-3 .
Рональд М. Новак: Млекопитающие мира Уокера . 6-е издание. Издательство Университета Джонса Хопкинса, Балтимор, 1999 г., ISBN 0-8018-5789-9 .
Томас С. Кемп: Происхождение и эволюция млекопитающих Издательство Оксфордского университета, Оксфорд, 2005. ISBN 0-19-850761-5 .
А. Х. Мюллер: Учебник палеозоологии, Том III: Позвоночные животные , Часть 3: Млекопитающие. 2-е издание. Густав Фишер Верлаг, Йена и Штутгарт, 1989 год. ISBN 3-334-00223-3 .
Arc.Ask3.Ru Номер скриншота №: e276089fe942270adef56f7b8df824b2__1722285360 URL1:https://arc.ask3.ru/arc/aa/e2/b2/e276089fe942270adef56f7b8df824b2.html Заголовок, (Title) документа по адресу, URL1: Perissodactyla - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)