Мутация сдвига рамки

Мутация сдвига рамки считывания (также называемая ошибкой кадрирования или сдвигом рамки считывания ) — это генетическая мутация, вызванная вставками ( инсерциями или делециями ) ряда нуклеотидов в последовательности ДНК, которая не делится на три. Из-за триплетного характера экспрессии генов кодонами от вставка или удаление может изменить рамку считывания (группировку кодонов), что приведет к совершенно отличной трансляции оригинала . Чем раньше в последовательности происходит делеция или вставка, тем более изменен белок. [ 1 ] Мутация сдвига рамки считывания — это не то же самое, что однонуклеотидный полиморфизм , при котором нуклеотид заменяется, а не вставляется или удаляется. Мутация сдвига рамки считывания обычно приводит к считыванию кодонов после мутации, кодирующих разные аминокислоты. Мутация сдвига рамки также изменит первый стоп-кодон («UAA», «UGA» или «UAG»), встречающийся в последовательности. Создаваемый полипептид может быть аномально коротким или аномально длинным и, скорее всего, не будет функциональным. [ 2 ]
Мутации сдвига рамки считывания очевидны при тяжелых генетических заболеваниях, таких как болезнь Тея-Сакса ; они повышают восприимчивость к некоторым видам рака и классам семейной гиперхолестеринемии ; в 1997 году, [ 3 ] мутация сдвига рамки считывания была связана с устойчивостью к инфекции ретровирусом ВИЧ. Мутации сдвига рамки считывания были предложены в качестве источника биологической новизны, как и в случае с предполагаемым созданием нейлоназы , однако эта интерпретация противоречива. Исследование Негоро и др. (2006) [ 4 ] обнаружили, что мутация сдвига рамки считывания вряд ли была причиной и что скорее замена двух аминокислот в активном сайте предковой эстеразы привела к образованию нейлоназы.
Фон
[ редактировать ]Информация, содержащаяся в ДНК, определяет функцию белка в клетках всех организмов. Транскрипция и трансляция позволяют передавать эту информацию для создания белков. Однако ошибка в чтении этого сообщения может привести к неправильной функции белка и в конечном итоге вызвать заболевание, даже если клетка принимает различные корректирующие меры. Генетическая информация передается через ДНК для синтеза белка внутри клеток. Неправильная интерпретация может привести к нарушению функции и заболеванию, несмотря на механизмы клеточной коррекции.
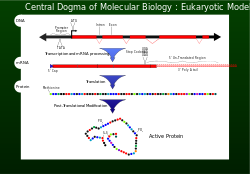
Центральная догма
[ редактировать ]В 1956 году Фрэнсис Крик описал поток генетической информации от ДНК к определенному расположению аминокислот для создания белка как центральную догму. [ 1 ] Для правильного функционирования клетки необходимо, чтобы белки вырабатывались точно для структурной и каталитической активности. Неправильно созданный белок может оказать пагубное воздействие на жизнеспособность клеток и в большинстве случаев привести к тому, что высший организм станет нездоровым из-за аномальных клеточных функций. Чтобы гарантировать, что геном успешно передает информацию, механизмы корректуры , такие как экзонуклеазы и системы репарации несоответствий включаются в репликацию ДНК . [ 1 ]
Транскрипция и перевод
[ редактировать ]После репликации ДНК считывание выбранного участка генетической информации осуществляется путем транскрипции . [ 1 ] Нуклеотиды, содержащие генетическую информацию, теперь находятся на одноцепочечной информационной матрице, называемой мРНК . мРНК включается в субъединицу рибосомы и взаимодействует с рРНК . Генетическая информация, несущаяся в кодонах мРНК, теперь считывается (декодируется) антикодонами тРНК. По мере считывания каждого кодона (триплета) аминокислоты соединяются до тех пор, пока не будет достигнут стоп-кодон (UAG, UGA или UAA). В этот момент полипептид (белок) синтезируется и высвобождается. [ 1 ] На каждые 1000 аминокислот, включенных в белок, не более одной неправильной. Такая точность распознавания кодонов, сохраняющая важность правильной рамки считывания, достигается за счет правильного спаривания оснований в сайте рибосомы А, GTP- гидролизной активности EF-Tu (форма кинетической стабильности) и механизма корректуры при высвобождении EF-Tu. . [ 1 ]
Сдвиг рамки считывания может также происходить во время профазной белки gag-pol-env трансляции, приводя к образованию различных белков из перекрывающихся открытых рамок считывания, таких как ретровирусные . Это довольно часто встречается у вирусов , а также у бактерий и дрожжей (Farabaugh, 1996). Обратная транскриптаза , в отличие от РНК-полимеразы II , считается более сильной причиной возникновения мутаций сдвига рамки считывания. В экспериментах только 3–13% всех мутаций сдвига рамки возникали из-за РНК-полимеразы II. У прокариот частота ошибок, вызывающих мутации сдвига рамки считывания, находится лишь где-то в диапазоне 0,0001 и 0,00001. [ 5 ]
Существует несколько биологических процессов, которые помогают предотвратить мутации сдвига рамки считывания. Возникают обратные мутации, которые изменяют мутированную последовательность обратно на исходную последовательность дикого типа . Другой возможностью коррекции мутации является использование мутации-супрессора . Это компенсирует эффект исходной мутации, создавая вторичную мутацию, сдвигая последовательность, чтобы можно было прочитать правильные аминокислоты. Гид-РНК также можно использовать для вставки или удаления уридина в мРНК после транскрипции, это позволяет получить правильную рамку считывания. [ 1 ]
Важность кодон-триплета
[ редактировать ]
Кодон — это набор из трех нуклеотидов , триплет, кодирующий определенную аминокислоту . Первый кодон устанавливает рамку считывания, благодаря чему начинается новый кодон. основной цепи белка Аминокислотная последовательность определяется смежными тройками. [ 6 ] Кодоны играют ключевую роль в трансляции генетической информации для синтеза белков. Рамка считывания устанавливается в момент начала трансляции мРНК и поддерживается при чтении одного триплета в другой. Чтение генетического кода подчиняется трем правилам контроля кодонов в мРНК. Во-первых, кодоны читаются в направлении от 5’ к 3’. Во-вторых, кодоны не перекрываются и в сообщении нет пробелов. Последнее правило, как сказано выше, заключается в том, что сообщение транслируется в фиксированной рамке чтения. [ 1 ]
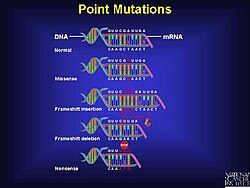
Механизм
[ редактировать ]Мутации сдвига рамки считывания могут возникать случайно или быть вызваны внешним стимулом. Обнаружение мутаций сдвига рамки считывания может происходить несколькими различными методами. Сдвиг рамки считывания — это лишь один из типов мутаций, которые могут привести к образованию неполных или неправильных белков, но на их долю приходится значительный процент ошибок в ДНК. В неизмененном гене кодоны (тройки нуклеотидов) интерпретируются последовательно, причем каждый кодон кодирует определенную аминокислоту. кислота. Это известно как стандартная рамка считывания. Однако в случае мутаций сдвига рамки считывания в последовательность ДНК вставляется дополнительный нуклеотид (или более), нарушая типичную рамку считывания и вызывая сдвиг последовательности.
Эта вставка вызывает сдвиг рамки считывания из-за триплетной природы генетического кода. Например, добавление дополнительной буквы «А» приводит к сдвигу последовательности, вызывая считывание совершенно другого набора кодонов. Это отклонение в генетической информации приводит к тому, что рибосома, которая считывает мРНК для синтеза белка, неправильно интерпретирует генетические данные. Следовательно, генерируется совершенно другой ряд аминокислот, что приводит к образованию измененной последовательности белка. В большинстве случаев новая рамка считывания приводит к ранней встрече со стоп-кодоном, что приводит к образованию укороченного и обычно неактивного белка. Эту форму мутации называют ранним стоп-кодоном или нонсенс-мутацией.
Генетический или экологический
[ редактировать ]Это генетическая мутация на уровне нуклеотидных оснований. Почему и как происходят мутации сдвига рамки считывания, постоянно ведется поиск. Было проведено экологическое исследование, в частности, производство мутаций сдвига рамки, индуцированных УФ-излучением , ДНК-полимеразами, дефицитными по экзонуклеазной активности 3' → 5'. Нормальная последовательность 5' GTC GTT TTA CAA 3' была заменена на GTC GTT T TTA CAA (MIDT) или GTC GTT C TTA CAA (MIDC) для изучения сдвигов рамки. ДНК-полимеразы E. coli pol I Kf и T7, Мутантные ферменты лишенные экзонуклеазной активности 3' → 5', производят УФ-индуцированные ревертанты с более высокой частотой, чем их опытные аналоги по экзонуклеазе . Данные показывают, что потеря корректурной активности увеличивает частоту сдвигов кадров, вызванных УФ-излучением. [ 7 ]
Обнаружение
[ редактировать ]флуоресценция
[ редактировать ]Влияние соседних оснований и вторичной структуры на обнаружение частоты мутаций сдвига рамки считывания было тщательно исследовано с использованием флуоресценции . Флуоресцентно-меченная ДНК с помощью аналогов оснований позволяет изучать локальные изменения последовательности ДНК. [ 8 ] Исследования влияния длины цепи праймера показывают, что равновесная смесь четырех конформаций гибридизации наблюдалась, когда основания матрицы образовывали петлю в виде выпуклости, т.е. структура, фланкированная с обеих сторон дуплексной ДНК. Напротив, двухпетлевая структура с необычной несложенной конформацией ДНК на ее нижнем крае наблюдалась, когда экструдированные основания располагались на стыке праймер-матрица, показывая, что несовпадения могут быть модифицированы соседней вторичной структурой ДНК. [ 9 ]
Секвенирование
[ редактировать ]
Секвенирование по Сэнгеру и пиросеквенирование — это два метода, которые использовались для обнаружения мутаций сдвига рамки считывания, однако вполне вероятно, что полученные данные будут не самого высокого качества. Тем не менее, с помощью секвенирования Сэнгера было идентифицировано 1,96 миллиона инделей , которые не пересекаются с другими базами данных. Когда наблюдается мутация сдвига рамки считывания, ее сравнивают с базой данных мутаций генома человека (HGMD), чтобы определить, оказывает ли мутация повреждающий эффект. Это делается путем рассмотрения четырех функций. Во-первых, соотношение между затронутой и консервативной ДНК, во-вторых, расположение мутации относительно транскрипта, в-третьих, соотношение консервативных и затронутых аминокислот и, наконец, расстояние от индели до конца экзона . [ 10 ]
Массивно-параллельное секвенирование — это новый метод, который можно использовать для обнаружения мутаций. Используя этот метод, можно секвенировать до 17 гигабаз одновременно, в отличие от ограниченного диапазона секвенирования по Сэнгеру, составляющего всего около 1 килобаз. Для проведения этого теста доступно несколько технологий, и рассматривается возможность его использования в клинических приложениях. [ 11 ] При тестировании на различные виды рака современные методы позволяют рассматривать только один ген за раз. Массивно-параллельное секвенирование позволяет одновременно выявить множество мутаций, вызывающих рак, в отличие от нескольких конкретных тестов. [ 12 ] Эксперимент по определению точности этого нового метода секвенирования был протестирован на 21 гене и не выявил ложноположительных сигналов о мутациях сдвига рамки считывания. [ 13 ]
Диагностика
[ редактировать ]США В патенте (5958684), выданном Леувеном в 1999 году, подробно описаны методы и реагенты для диагностики заболеваний, вызванных или связанных с геном, имеющим соматическую мутацию, вызывающую мутацию сдвига рамки считывания. Эти методы включают предоставление образца ткани или жидкости и проведение анализа гена на предмет мутации сдвига рамки считывания или белка, возникшего в результате мутации этого типа. Нуклеотидная последовательность подозреваемого гена получена из опубликованных последовательностей генов или в результате клонирования и секвенирования подозреваемого гена. Затем прогнозируется аминокислотная последовательность, кодируемая геном. [ 14 ] Секвенирование NA: секвенирование по Сэнгеру или секвенирование следующего поколения (NGS) можно использовать для прямого секвенирования ДНК и идентификации вставок или делеций. Полимеразная цепная реакция (ПЦР): ПЦР можно использовать для амплификации конкретной области, содержащей мутацию, для последующего анализа. Мультиплексная амплификация зонда, зависимая от лигирования (MLPA): MLPA — это метод, используемый для обнаружения изменений количества копий и небольших вставок или делеций. Сравнительная геномная гибридизация (CGH): CGH используется для обнаружения хромосомного дисбаланса, который может включать крупные вставки или делеции.
Частота
[ редактировать ]Несмотря на правила, которые управляют генетическим кодом и различными механизмами, присутствующими в клетке и обеспечивающими правильную передачу генетической информации в процессе репликации ДНК, а также во время трансляции, мутации все же происходят; Мутация сдвига рамки считывания — не единственный тип. Есть как минимум два других типа признанных точковых мутаций, а именно миссенс-мутация и нонсенс-мутация . [ 1 ] Мутация сдвига рамки может радикально изменить способность кодирования (генетическую информацию) сообщения. [ 1 ] Небольшие вставки или делеции (менее 20 пар оснований) составляют 24% мутаций, которые проявляются при признанных в настоящее время генетических заболеваниях. [ 10 ]
Обнаружено, что мутации сдвига рамки считывания чаще встречаются в повторяющихся областях ДНК. Причиной этого является проскальзывание фермента полимеразы в повторяющихся областях, что позволяет мутациям проникнуть в последовательность . [ 15 ] Можно провести эксперименты , чтобы определить частоту мутации сдвига рамки считывания путем добавления или удаления заранее установленного количества нуклеотидов. Были проведены эксперименты по добавлению четырех пар оснований, называемые экспериментами +4, но команда из Университета Эмори изучила разницу в частоте мутаций, добавляя и удаляя пару оснований. Было показано, что не было разницы в частоте добавления и удаления пары оснований. Однако есть разница в результате использования белка. [ 15 ]
Болезнь Хантингтона является одним из девяти нарушений повторения кодонов, вызванных мутациями расширения полиглутамина, которые включают спинно-мозжечковую атаксию (SCA) 1, 2, 6, 7 и 3, спинобульбарную мышечную атрофию и дентаторубал-паллидолуизианотрофию. Может существовать связь между заболеваниями, вызванными полиглутамином и мутациями расширения полиаланина, например сдвигом рамки исходного продукта гена SCA3, кодирующего CAG/полиглутамины, на GCA/полиаланин. Рибосомальное проскальзывание во время трансляции белка SCA3 было предложено как механизм, приводящий к сдвигу от полиглутамина к рамке, кодирующей полиаланин. Делеция динуклеотида или вставка одного нуклеотида в полиглутаминовый тракт экзона 1 хантингтина сместят CAG, рамку, кодирующую полиглутаминин, на +1 (сдвиг рамки на +1) к GCA, рамке, кодирующей полиаланин, и введут новый эпитоп на С-конец Htt экзон 1 (APAAAAPAATRPCG). [ 16 ]
Болезни
[ редактировать ]Некоторые заболевания имеют мутации сдвига рамки считывания как, по крайней мере, часть причины. Знание распространенных мутаций также может помочь в диагностике заболевания. В настоящее время предпринимаются попытки с пользой использовать мутации сдвига рамки считывания при лечении заболеваний, изменяя рамку считывания аминокислот.
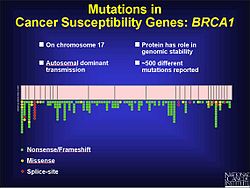

Рак
[ редактировать ]Известно, что мутации сдвига рамки считывания являются фактором колоректального рака, а также других видов рака с микросателлитной нестабильностью . Как указывалось ранее, мутации сдвига рамки считывания с большей вероятностью возникают в области повторяющейся последовательности. Когда восстановление несоответствия ДНК не устраняет добавление или удаление оснований, эти мутации с большей вероятностью будут патогенными. Частично это может быть связано с тем, что опухоли не приказано прекратить рост. Эксперименты на дрожжах и бактериях помогают выявить характеристики микросателлитов, которые могут способствовать восстановлению дефектных несоответствий ДНК. К ним относятся длина микросателлита , состав генетического материала и чистота повторов. Согласно экспериментальным результатам, более длинные микросателлиты имеют более высокую частоту мутаций сдвига рамки считывания. Фланкирующая ДНК также может способствовать мутациям сдвига рамки считывания. [ 17 ] При раке простаты мутация сдвига рамки изменяет открытую рамку считывания (ORF) и предотвращает апоптоза возникновение . Это приводит к нерегулируемому росту опухоли . Хотя существуют факторы окружающей среды, которые способствуют прогрессированию рака простаты , существует также генетический компонент. В ходе тестирования кодирующих регионов на выявление мутаций было обнаружено 116 генетических вариантов, в том числе 61 мутация сдвига рамки считывания. [ 18 ] В 17-й хромосоме имеется более 500 мутаций, которые, по-видимому, играют роль в развитии рака молочной железы и яичников в гене BRCA1, многие из которых являются сдвигом рамки считывания. [ 19 ]
болезнь Крона
[ редактировать ]Болезнь Крона связана с геном NOD2. Мутация представляет собой вставку цитозина в положение 3020. Это приводит к преждевременному образованию стоп-кодона, укорачивая белок, который должен транскрибироваться. Когда белок способен нормально формироваться, он реагирует на бактериальные липосахариды, при этом мутация 3020insC предотвращает реакцию белка. [ 20 ]
Муковисцидоз
[ редактировать ]Муковисцидоз (МВ) – заболевание, в основе которого лежит мутация гена регулятора трансмембранной проводимости МВ (CFTR). Выявлено более 1500 мутаций, но не все вызывают заболевание. [ 21 ] Большинство случаев муковисцидоза являются результатом мутации ∆F508, которая удаляет всю аминокислоту. Для диагностики CF представляют интерес две мутации сдвига рамки считывания: CF1213delT и CF1154-insTC. Обе эти мутации обычно происходят в тандеме по крайней мере с еще одной мутацией. Оба они приводят к небольшому снижению функции легких и встречаются примерно у 1% обследованных пациентов. Эти мутации были идентифицированы с помощью секвенирования по Сэнгеру. [ 22 ]
ВИЧ
[ редактировать ]CCR5 является одним из кофакторов проникновения в клетку, связанных с ВИЧ, чаще всего связан со штаммами, не индуцирующими синцитий, и наиболее выражен у пациентов с ВИЧ, в отличие от пациентов со СПИДом. Делеция в 32 пары оснований в CCR5 была идентифицирована как мутация, которая сводит на нет вероятность ВИЧ-инфекции. Эта область открытой рамки считывания ORF содержит мутацию сдвига рамки считывания, приводящую к преждевременному стоп-кодону. Это приводит к потере функции ВИЧ-корецептора in vitro. CCR5-1 считается диким типом, а CCR5-2 считается мутантным аллелем. Те, у кого была гетерозиготная мутация CCR5, были менее восприимчивы к развитию ВИЧ. В исследовании, несмотря на высокий уровень воздействия вируса ВИЧ, не было ни одного человека, гомозиготного по мутации CCR5, который дал бы положительный результат на ВИЧ. [ 3 ]
Болезнь Тея-Сакса
[ редактировать ]Болезнь Тея-Сакса — смертельное заболевание, поражающее центральную нервную систему. Чаще всего встречается у младенцев и маленьких детей. Прогрессирование заболевания начинается еще в утробе матери , но симптомы не появляются примерно до 6-месячного возраста. Лекарства от этой болезни не существует. [ 23 ] Известно, что мутации в гене β-гексозаминидазы А (Hex A) влияют на возникновение синдрома Тея-Сакса: описано 78 мутаций различных типов, 67 из которых, как известно, вызывают заболевание. Большинство наблюдаемых мутаций (65/78) представляют собой одноосновные замены или SNP, 11 делеций, 1 большую и 10 малых, а также 2 инсерции. 8 из наблюдаемых мутаций представляют собой сдвиг рамки считывания, 6 делеций и 2 инсерции. Вставка из 4 пар оснований в экзоне 11 наблюдается в 80% случаев болезни Тея-Сакса у еврейского населения ашкенази . Мутации сдвига рамки считывания приводят к появлению раннего стоп-кодона, который, как известно, играет роль в заболевании у младенцев. Заболевание с отсроченным началом, по-видимому, вызвано четырьмя различными мутациями, одна из которых представляет собой делецию трех пар оснований. [ 24 ]
Синдром Смита-Магениса
[ редактировать ]Синдром Смита-Магениса (СМС) представляет собой сложный синдром, включающий интеллектуальные нарушения, нарушения сна, поведенческие проблемы и различные черепно-лицевые, скелетные и висцеральные аномалии. В большинстве случаев СМС имеется общая делеция размером ~3,5 Мб, которая включает ген индуцированного ретиноевой кислотой-1 ( RAI1 ). Другие случаи иллюстрируют вариабельность фенотипа SMS , ранее не выявленную для мутации RAI1, включая потерю слуха, отсутствие самооскорбительного поведения и умеренные глобальные задержки. Секвенирование RAI1 выявило мутацию гептамерного C-тракта (CCCCCCC) в экзоне 3, приводящую к мутациям сдвига рамки считывания. Из семи зарегистрированных мутаций сдвига рамки считывания, происходящих в поли-С-трактах в RAI1, четыре случая (~ 57%) происходят в этом гептамерном С-тракте. Результаты показывают, что этот гептамерный C-тракт является предпочтительной горячей точкой рекомбинации вставки/делеции (SNindels) и, следовательно, основной мишенью для анализа у пациентов с подозрением на мутации в RAI1. [ 25 ]
Гипертрофическая кардиомиопатия
[ редактировать ]Гипертрофическая кардиомиопатия является наиболее частой причиной внезапной смерти у молодых людей, в том числе у тренированных спортсменов, и обусловлена мутациями генов, кодирующих белки сердечного саркомера. Мутации в гене тропонина С ( TNNC1 ) являются редкой генетической причиной гипертрофической кардиомиопатии. Недавнее исследование показало, что мутация сдвига рамки считывания (c.363dupG или p.Gln122AlafsX30) в тропонине С была причиной гипертрофической кардиомиопатии (и внезапной сердечной смерти) у 19-летнего мужчины. [ 26 ]
Лекарства
[ редактировать ]Найти лекарство от болезней, вызванных мутациями сдвига рамки считывания, удается редко. Исследования по этому вопросу продолжаются. Одним из примеров является первичный иммунодефицит (ПИД), наследственное заболевание, которое может привести к увеличению числа инфекций. Существует 120 генов и 150 мутаций, которые играют роль в развитии первичных иммунодефицитов. Стандартным лечением в настоящее время является генная терапия , но это очень рискованное лечение и часто может привести к другим заболеваниям, таким как лейкемия. Процедуры генной терапии включают модификацию слитого белка нуклеазы цинкового фингера, расщепление обоих концов мутации, что, в свою очередь, удаляет ее из последовательности. , опосредованный антисмысловыми олигонуклеотидами, Пропуск экзонов является еще одной возможностью мышечной дистрофии Дюшенна . Этот процесс позволяет пройти мутацию, так что остальная часть последовательности остается в рамке, а функция белка остается неизменной. Однако это не излечивает болезнь, а лишь лечит симптомы и применимо только к структурным белкам или другим повторяющимся генам. Третий вид ремонта. ревертантный мозаицизм , который возникает естественным путем путем создания обратной мутации или мутации во втором сайте, которая корректирует рамку считывания. Эта реверсия может произойти в результате внутригенной рекомбинации , митотической конверсии гена, проскальзывания ДНК второго сайта или сайт-специфической реверсии. Это возможно при некоторых заболеваниях, таких как Х-сцепленный тяжелый комбинированный иммунодефицит (ТКИД), синдром Вискотта-Олдрича и синдром Блума . Не существует лекарств или других фармакогеномных методов, помогающих при ПИД. [ 27 ]
В европейском патенте (EP1369126A1), выданном Борком в 2003 году, описан метод, используемый для профилактики рака и лечения рака и предраковых заболеваний, таких как спорадические опухоли с дефицитом репарации несоответствия ДНК (MMR) и опухоли, связанные с HNPCC. Идея состоит в том, чтобы использовать иммунотерапию с использованием комбинаторных смесей опухолеспецифичных пептидов, полученных из мутаций сдвига рамки считывания, чтобы вызвать цитотоксический Т-клеточный ответ, специально направленный против опухолевых клеток. [ 28 ]
См. также
[ редактировать ]- Трансляционный сдвиг рамки
- Мутация
- Транскрипция (генетика)
- Перевод (биология)
- кодон
- белок
- рамка для чтения
- точечная мутация
- болезнь Крона
- Болезнь Тея-Сакса
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж Лосик, Ричард; Уотсон, Джеймс Д.; Бейкер, Таня А.; Белл, Стивен; Ганн, Александр; Левин, Майкл В. (2008). Молекулярная биология гена (6-е изд.). Сан-Франциско: Пирсон/Бенджамин Каммингс. ISBN 978-0-8053-9592-1 .
- ^ «ДНК постоянно меняется в процессе мутации» . Природа . Проверено 17 мая 2019 г.
- ^ Перейти обратно: а б Циммерман П.А., Баклер-Уайт А., Алхатиб Г., Сполдинг Т., Кубофчик Дж., Комбадьер С., Вайсман Д., Коэн О., Рауберт А., Лам Г., Ваккарецца М., Кеннеди П.Е., Кумарасвами В., Георгий Дж.В., Детельс Р., Хантер Дж., Чопек М., Бергер Э.А., Фаучи А.С., Натман Т.Б., Мерфи П.М. (январь 1997 г.). «Наследственная устойчивость к ВИЧ-1, обусловленная инактивирующей мутацией хемокинового рецептора CC 5: исследования в популяциях с контрастными клиническими фенотипами, определенным расовым происхождением и количественным риском» . Молекулярная медицина . 3 (1): 23–36. ПМК 2230106 . ПМИД 9132277 .
- ^ Негоро С., Оки Т., Сибата Н., Мизуно Н., Вакитани Ю., Цурукаме Дж., Мацумото К., Кавамото И., Такео М., Хигути Ю. (ноябрь 2005 г.). «Рентгеноструктурный анализ 6-аминогексаноат-димергидролазы: молекулярная основа рождения фермента, разлагающего олигомер нейлона» . J Биол Хим . 280 (47): 39644–52. дои : 10.1074/jbc.m505946200 . ПМИД 16162506 .
- ^ Чжан, Дж (август 2004 г.). «РНК-полимераза II хозяина вносит минимальный вклад в ретровирусные мутации сдвига рамки считывания» . Журнал общей вирусологии . 85 (Часть 8): 2389–95. дои : 10.1099/vir.0.80081-0 . ПМИД 15269381 .
- ^ Кокс, Майкл; Нельсон, Дэвид Р.; Ленинджер, Альберт Л. (2008). Ленингерские принципы биохимии . Сан-Франциско: WH Freeman. ISBN 978-0-7167-7108-1 .
- ^ Сагер, Дафна; Теркингтон, Эдит; Ачарья, Соня; Штраус, Бернард (июль 1994 г.). «Продуцирование мутаций сдвига рамки считывания, индуцированных УФ-излучением, in vitro с помощью ДНК-полимераз с дефицитом экзонуклеазной активности 3' → 5'». Журнал молекулярной биологии . 240 (3): 226–242. дои : 10.1006/jmbi.1994.1437 . ПМИД 8028006 .
- ^ Джонсон, Нил П.; Уолтер А. Баасе; Питер Х. фон Хиппель (март 2004 г.). «Низкоэнергетический круговой дихроизм 2-аминопуриндинуклеотида как зонд локальной конформации ДНК и РНК» . Proc Natl Acad Sci США . 101 (10): 3426–31. Бибкод : 2004PNAS..101.3426J . дои : 10.1073/pnas.0400591101 . ПМЦ 373478 . ПМИД 14993592 .
- ^ Баасе, Уолтер А.; Дэвис Хосе; Бенджамин К. Понедел; Питер Х. фон Хиппель; Нил П. Джонсон (2009). «ДНК-модели делеций сдвига рамки тринуклеотида: образование петель и выпуклостей на стыке праймер-матрица» . Исследования нуклеиновых кислот . 37 (5): 1682–9. дои : 10.1093/нар/gkn1042 . ПМЦ 2655659 . ПМИД 19155277 .
- ^ Перейти обратно: а б Ху, Дж; Нг, ПК (9 февраля 2012 г.). «Прогнозирование эффектов смещения кадра» . Геномная биология . 13 (2): С9. дои : 10.1186/gb-2012-13-2-r9 . ПМЦ 3334572 . ПМИД 22322200 .
- ^ Такер, Трейси; Марра, Марко; Фридман, Ян М. (2009). «Массовое параллельное секвенирование: следующий большой шаг в генетической медицине» . Американский журнал генетики человека . 85 (2): 142–154. дои : 10.1016/j.ajhg.2009.06.022 . ПМЦ 2725244 . ПМИД 19679224 .
- ^ Уолш, Т.; Касадей, С.; Ли, МК; Пеннил, CC; Норд, АС; Торнтон, AM; Роб, В.; Агнью, К.Дж.; Стрэй, С.М.; Викраманаяке, А.; Норквист, Б.; Пеннингтон, КП; Гарсия, РЛ; Кинг, М.-К.; Свишер, Э.М. (2011). «С обложки: мутации в 12 генах наследственного рака яичников, фаллопиевых труб и брюшины, выявленные с помощью массового параллельного секвенирования» . Proc Natl Acad Sci США . 108 (44): 18032–7. Бибкод : 2011PNAS..10818032W . дои : 10.1073/pnas.1115052108 . ПМК 3207658 . ПМИД 22006311 .
- ^ Уолш, Т.; Ли, МК; Касадей, С.; Торнтон, AM; Стрэй, С.М.; Пеннил, К.; Норд, АС; Манделл, Дж.Б.; Свишер, Э.М.; Кинг, М.-К. (2010). «Обнаружение наследственных мутаций рака молочной железы и яичников с использованием геномного захвата и массового параллельного секвенирования» . Proc Natl Acad Sci США . 107 (28): 12629–33. Бибкод : 2010PNAS..10712629W . дои : 10.1073/pnas.1007983107 . ПМК 2906584 . ПМИД 20616022 .
- ^ Патент США 5 958 684 (28 сентября 1999 г.) «Диагностика нейродегенеративных заболеваний», автор Leeuwen et al.
- ^ Перейти обратно: а б Харф, Б.Д.; Джинкс-Робертсон, С. (июль 1999 г.). «Удаление промежуточных продуктов сдвига рамки считывания с помощью белков репарации несоответствия в Saccharomyces cerevisiae» . Молекулярная и клеточная биология . 19 (7): 4766–73. дои : 10.1128/MCB.19.7.4766 . ПМЦ 84275 . ПМИД 10373526 .
- ^ Дэвис, Дж. Э.; Рубинштейн, округ Колумбия (2006). «Продукты сдвига рамки полиаланин и полисерин при болезни Гентингтона» . Журнал медицинской генетики . 43 (11): 893–896. дои : 10.1136/jmg.2006.044222 . ПМК 2563184 . ПМИД 16801344 .
- ^ Шмольдт, А; Бенте, ХФ; Хаберланд, Г. (1 сентября 1975 г.). «Метаболизм дигитоксина микросомами печени крысы» . Биохимическая фармакология . 24 (17): 1639–41. дои : 10.1016/0006-2952(75)90094-5 . hdl : 10033/333424 . ПМИД 10 .
- ^ Сюй, СяоЛин; Лю, Фэн; Шэнь, ЦзяньГо, Цзичжун; Ван, Чен, Линь; Сюй, Мин (май 2013 г.). «Рак простаты человека с помощью RNA-Seq». Gene . 519 (2): 343–7. doi : 10.1016/j.gene.2013.01.046 . PMID 23434521 .
- ^ «Геномика рака» . Национальный институт рака при Национальном институте здравоохранения. Архивировано из оригинала 18 марта 2013 года . Проверено 24 марта 2013 г.
- ^ Огура Ю, Бонен Д.К., Инохара Н., Николае Д.Л., Чен Ф.Ф., Рамос Р., Бриттон Х., Моран Т., Каралиускас Р., Дюрр Р.Х., Ачкар Дж.П., Брант С.Р., Бэйлесс Т.М., Киршнер Б.С., Ханауэр С.Б., Нуньес Г., Чо Дж.Х. (31 мая 2001 г.). «Мутация сдвига рамки считывания в NOD2, связанная с предрасположенностью к болезни Крона» (PDF) . Природа . 411 (6837): 603–6. Бибкод : 2001Natur.411..603O . дои : 10.1038/35079114 . hdl : 2027.42/62856 . ПМИД 11385577 . S2CID 205017657 .
- ^ Фаррелл П.М., Розенштейн Б.Дж., Уайт ТБ, Аккурсо Ф.Дж., Кастеллани С., Каттинг Г.Р., Дьюри П.Р., Легрис В.А., Мэсси Дж., Парад Р.Б., Рок М.Дж., Кэмпбелл П.В. (2008). «Руководство по диагностике муковисцидоза у новорожденных и пожилых людей: консенсусный отчет Фонда муковисцидоза» . Журнал педиатрии . 153 (2): С4–С14. дои : 10.1016/j.jpeds.2008.05.005 . ПМК 2810958 . ПМИД 18639722 .
- ^ Яннуцци, MC; Стерн, Р.К.; Коллинз, Ф.С.; Достопочтенный, Коннектикут; Хидака, Н.; Стронг, Т; Беккер, Л; Драмм, МЛ; Уайт, МБ; Джеррард, Б. (февраль 1991 г.). «Две мутации сдвига рамки считывания в гене муковисцидоза» . Американский журнал генетики человека . 48 (2): 227–31. ПМК 1683026 . ПМИД 1990834 .
- ^ «Знакомство с болезнью Тея-Сакса» . Национальный институт исследования генома человека . Проверено 24 марта 2013 г.
- ^ Меровиц, Р. (1997). «Мутации, вызывающие болезнь Тея-Сакса, и нейтральный полиморфизм в гене Hex A» . Человеческая мутация . 9 (3): 195–208. doi : 10.1002/(SICI)1098-1004(1997)9:3<195::AID-HUMU1>3.0.CO;2-7 . ПМИД 9090523 .
- ^ Труонг, Хоа Т; Даддинг, Трейси; Бланшар, Кристофер Л.; Элси, Сара Х (2010). «Горячая точка мутации сдвига рамки, выявленная при синдроме Смита-Магениса: отчет о случае и обзор литературы» . BMC Медицинская генетика . 11 (1): 142. дои : 10.1186/1471-2350-11-142 . ПМЦ 2964533 . ПМИД 20932317 .
- ^ Чанг В.К., Китнер С., Марон Б.Дж. (июнь 2011 г.). «Новая мутация сдвига рамки считывания в тропонине C (TNNC1), связанная с гипертрофической кардиомиопатией и внезапной смертью». Кардиол Янг . 21 (3): 345–8. дои : 10.1017/S1047951110001927 . ПМИД 21262074 . S2CID 46682245 .
- ^ Ху, Хайлян; Гатти, Ричард А. (2008). «Новые подходы к лечению первичных иммунодефицитов: фиксация мутаций химическими препаратами» . Современное мнение в области аллергии и клинической иммунологии . 8 (6): 540–6. дои : 10.1097/ACI.0b013e328314b63b . ПМК 2686128 . ПМИД 18978469 .
- ^ Европейский патент [1] (10 декабря 2003 г.) «Использование кодирования пептидов, полученных из мутации сдвига рамки считывания микросателлитной области, для лечения рака» Bork et al.
Дальнейшее чтение
[ редактировать ]- Фарабо П.Дж. (1996). «Программированный поступательный сдвиг кадра» . Анну. Преподобный Жене . 30 (1): 507–28. дои : 10.1146/annurev.genet.30.1.507 . ПМК 239420 . ПМИД 8982463 .
- Льюис, Рики (2005). Генетика человека: концепции и приложения (6-е изд.). Бостон, Массачусетс: МакГроу Хилл. стр. 227–8. ISBN 978-0-07-111156-0 .
- «Нилоназные ферменты» . 20 апреля 2004 года . Проверено 2 июня 2009 г.
Внешние ссылки
[ редактировать ]- Сдвиг рамки + мутация Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- База данных NCBI dbSNP — «центральное хранилище как одноосновных нуклеотидных замен, так и коротких делеционных и инсерционных полиморфизмов»
- Wise2 - выравнивает белок по последовательности ДНК, допуская сдвиги рамки и интроны.
- FastY — сравнение последовательности ДНК с базой данных последовательностей белков, с учетом пропусков и сдвигов рамки считывания.
- Путь. Архивировано 19 июля 2011 г. в Wayback Machine — инструмент, который сравнивает два белка сдвига рамки считывания ( обратной трансляции ). принцип
- HGMD - База данных мутаций генома человека