Центральная догма молекулярной биологии
Центральная догма молекулярной биологии касается потока генетической информации внутри биологической системы. Часто говорят: «ДНК производит РНК, а РНК производит белок». [1] хотя это не его первоначальный смысл. Впервые это было сформулировано Фрэнсисом Криком в 1957 году. [2] [3] затем опубликовано в 1958 году: [4] [5]
Центральная догма. Это означает, что однажды «информация» попала в белок, и она не может выйти обратно. Более подробно, передача информации от нуклеиновой кислоты к нуклеиновой кислоте или от нуклеиновой кислоты к белку может быть возможна, но передача от белка к белку или от белка к нуклеиновой кислоте невозможна. Информация здесь означает точное определение последовательности либо оснований нуклеиновой кислоты, либо аминокислотных остатков в белке.
Он повторил это в статье Nature , опубликованной в 1970 году: «Центральная догма молекулярной биологии касается детальной по остаткам передачи последовательной информации . Она утверждает, что такая информация не может быть передана обратно от белка к белку или нуклеиновой кислота." [6]
Вторая версия центральной догмы популярна, но неверна. Это упрощенный путь ДНК → РНК → белок, опубликованный Джеймсом Уотсоном в первом издании «Молекулярной биологии гена» (1965). Версия Уотсона отличается от версии Крика, поскольку Уотсон описывает двухэтапный процесс (ДНК → РНК и РНК → белок) как центральную догму. [7] Хотя догма, первоначально сформулированная Криком, остается в силе и сегодня, [6] [8] В версии Ватсона нет. [2]
Информация о биологической последовательности
[ редактировать ]Биополимеры , которые содержат ДНК, РНК и (поли) пептиды, представляют собой линейные полимеры (т.е. каждый мономер связан не более чем с двумя другими мономерами). Последовательность . их мономеров эффективно кодирует информацию Передача информации от одной молекулы к другой является точной, детерминированной передачей, при которой последовательность одного биополимера используется в качестве матрицы для создания другого биополимера с последовательностью, которая полностью зависит от последовательности исходного биополимера. Когда ДНК транскрибируется в РНК, к ней присоединяется ее комплемент. Коды ДНК A, G, T и C передаются в коды РНК U, C, A и G соответственно. Кодирование белков осуществляется группами по три штуки, известными как кодоны . Стандартная таблица кодонов применима к людям и млекопитающим, но некоторые другие формы жизни (включая митохондрии человека) [9] ) используйте разные переводы . [10]
Общие передачи биологической последовательной информации
[ редактировать ]Таблица трех классов передачи информации. Общий Особенный Неизвестный ДНК → ДНК РНК → ДНК Белок → ДНК ДНК → РНК РНК → РНК Белок → РНК РНК → белок Белок → белок ДНК → белок
Репликация ДНК
[ редактировать ]В том смысле, что репликация ДНК должна происходить, если генетический материал должен быть предоставлен для потомства любой клетки, как соматической , так и репродуктивной , копирование ДНК в ДНК, возможно, является фундаментальным шагом в передаче информации. Сложная группа белков, называемая реплисомой, осуществляет репликацию информации от родительской цепи к комплементарной дочерней цепи.
Транскрипция
[ редактировать ]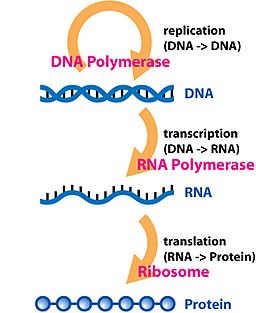
Транскрипция — это процесс, при котором информация, содержащаяся в участке ДНК, реплицируется в форме вновь собранной части информационной РНК (мРНК). Ферменты, облегчающие этот процесс, включают РНК-полимеразу и факторы транскрипции . В эукариотических клетках первичным транскриптом является пре-мРНК . Пре-мРНК должна быть обработана для продолжения трансляции. Процессинг включает добавление 5'-кэпа и поли-А-хвоста к цепи пре-мРНК с последующим сплайсингом . При необходимости происходит альтернативный сплайсинг , увеличивающий разнообразие белков, которые может производить любая отдельная мРНК. Продуктом всего процесса транскрипции (который начался с образования цепи пре-мРНК) является зрелая цепь мРНК.
Перевод
[ редактировать ]Зрелая мРНК попадает в рибосому , где транслируется . В прокариотических клетках, не имеющих ядерного компартмента, процессы транскрипции и трансляции могут быть связаны друг с другом без четкого разделения. В эукариотических клетках место транскрипции ( ядро клетки ) обычно отделено от места трансляции ( цитоплазмы ), поэтому мРНК должна транспортироваться из ядра в цитоплазму, где она может связываться рибосомами. Рибосома считывает триплетные кодоны мРНК , обычно начиная с AUG ( аденин - урацил - гуанин ) или инициаторного кодона метионина ниже сайта связывания рибосомы . Комплексы факторов инициации и факторов элонгации переводят аминоацилированные транспортные РНК (тРНК) в комплекс рибосома-мРНК, сопоставляя кодон мРНК с антикодоном тРНК. Каждая тРНК несет соответствующий аминокислотный остаток, который можно добавить к полипептидной синтезируемой цепи. Когда аминокислоты соединяются в растущую пептидную цепь, она начинает сворачиваться в правильную конформацию. Перевод заканчивается стоп-кодон , который может представлять собой триплет UAA, UGA или UAG.
мРНК не содержит всей информации для уточнения природы зрелого белка. Зарождающаяся полипептидная цепь, высвобождаемая из рибосомы, обычно требует дополнительной обработки, прежде чем появится конечный продукт. Во-первых, правильный процесс складывания сложен и жизненно важен. Для большинства белков требуются другие белки-шапероны, чтобы контролировать форму продукта. Некоторые белки затем вырезают внутренние сегменты из своих собственных пептидных цепей, сращивая свободные концы, ограничивающие разрыв; в таких процессах внутренние «отброшенные» секции называются интеинами . Другие белки необходимо разделить на несколько секций без сплайсинга. Некоторые полипептидные цепи должны быть сшиты, а другие должны быть присоединены к кофакторам, таким как гем (гем), прежде чем они станут функциональными.
Специальная передача биологической последовательной информации
[ редактировать ]Обратная транскрипция
[ редактировать ]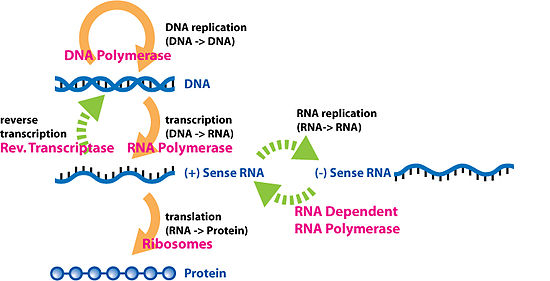
Обратная транскрипция — это перенос информации с РНК на ДНК (обратный нормальной транскрипции). Известно, что это происходит в случае ретровирусов , таких как ВИЧ , а также у эукариот в случае ретротранспозонов и синтеза теломер .Это процесс, посредством которого генетическая информация из РНК транскрибируется в новую ДНК. Семейство ферментов, участвующих в этом процессе, называется обратной транскриптазой .
Репликация РНК
[ редактировать ]Репликация РНК – это копирование одной РНК на другую. Многие вирусы размножаются таким образом. Ферменты, которые копируют РНК в новую РНК, называемые РНК-зависимыми РНК-полимеразами , также обнаружены у многих эукариот, где они участвуют в подавлении РНК . [11]
Редактирование РНК , при котором последовательность РНК изменяется комплексом белков и «направляющей РНК», также можно рассматривать как перенос РНК на РНК.
Прямая трансляция с ДНК в белок
[ редактировать ]Прямая трансляция ДНК в белок была продемонстрирована в бесклеточной системе (т.е. в пробирке) с использованием экстрактов E. coli , содержащих рибосомы, но не интактные клетки. Эти фрагменты клеток могли синтезировать белки из матриц одноцепочечной ДНК, выделенных из других организмов (например, мыши или жабы), и было обнаружено, что неомицин усиливает этот эффект. Однако было неясно, соответствует ли этот механизм трансляции именно генетическому коду. [12] [13]
Передача информации, явно не описанная в теории
[ редактировать ]Посттрансляционная модификация
[ редактировать ]После того как аминокислотные последовательности белка были транслированы с цепей нуклеиновых кислот, их можно редактировать с помощью соответствующих ферментов. Хотя это форма белка, влияющая на последовательность белка, которая явно не охвачена центральной догмой, существует не так много ярких примеров, когда связанные концепции двух областей имеют много общего друг с другом.
Синтез нерибосомальных пептидов
[ редактировать ]Некоторые белки синтезируются нерибосомальными пептидсинтетазами , которые могут представлять собой большие белковые комплексы, каждый из которых специализируется на синтезе только одного типа пептидов. Нерибосомальные пептиды часто имеют циклическую и/или разветвленную структуру и могут содержать непротеиногенные аминокислоты — оба этих фактора отличают их от белков, синтезируемых рибосомами. Примером нерибосомальных пептидов являются некоторые антибиотики.
Интеины
[ редактировать ]Интеин — это «паразитный» сегмент белка, который способен вырезать себя из цепи аминокислот при их выходе из рибосомы и воссоединить оставшиеся части пептидной связью таким образом, что основной «остов» белка не не развалиться. Это случай, когда белок меняет свою первичную последовательность по сравнению с последовательностью, первоначально кодируемой ДНК гена. Кроме того, большинство интеинов содержат хоминг-эндонуклеазу или домен HEG, который способен находить копию родительского гена, не включающую нуклеотидную последовательность интеина. При контакте с копией, не содержащей интеина, домен HEG инициирует механизм восстановления двухцепочечного разрыва ДНК . Этот процесс приводит к копированию последовательности интеина из исходного исходного гена в ген, не содержащий интеина. Это пример того, как белок напрямую редактирует последовательность ДНК, а также увеличивает наследственное распространение последовательности.
Метилирование
[ редактировать ]Вариации в состояниях метилирования ДНК могут экспрессии генов значительно изменить уровни . Вариации метилирования обычно происходят под действием ДНК- метилаз . Когда изменение является наследственным, оно считается эпигенетическим . Если изменение информационного статуса не передается по наследству, это будет соматический эпитип . Эффективное информационное содержание было изменено посредством воздействия белка или белков на ДНК, но первичная последовательность ДНК не изменилась.
Прионы
[ редактировать ]Прионы – это белки с определенными аминокислотными последовательностями в определенных конформациях. Они размножаются в клетках-хозяевах путем внесения конформационных изменений в другие молекулы белка с той же аминокислотной последовательностью, но с другой конформацией, функционально важной или вредной для организма. Как только белок трансформируется в прионную форму, он меняет функцию. В свою очередь, он может передавать информацию в новые клетки и переконфигурировать более функциональные молекулы этой последовательности в альтернативную форму приона. У некоторых типов прионов грибов это изменение непрерывное и прямое; информационный поток — Белок → Белок.
Некоторые ученые, такие как Ален Э. Бюссар и Юджин Кунин, утверждают, что наследование, опосредованное прионами, нарушает центральную догму молекулярной биологии. [14] [15] Однако Розалинд Ридли в книге «Молекулярная патология прионов» (2001) написала, что «прионная гипотеза не противоречит центральной догме молекулярной биологии — что информация, необходимая для производства белков, закодирована в нуклеотидной последовательности нуклеиновой кислоты — потому что она не утверждает, что белки реплицируются. Скорее, он утверждает, что внутри белковых молекул существует источник информации, который способствует их биологической функции, и что эта информация может передаваться другим молекулам». [16]
Естественная генная инженерия
[ редактировать ]Джеймс А. Шапиро утверждает, что расширенную совокупность этих примеров следует классифицировать как естественную генную инженерию , и их достаточно для фальсификации центральной догмы. Хотя Шапиро получил уважительное отношение к своей точке зрения, его критики не были убеждены, что его прочтение центральной догмы соответствует тому, что намеревался Крик. [17] [18]
Использование термина догма
[ редактировать ]В своей What автобиографии Mad Pursuit Крик написал о выборе слова «догма» и о некоторых проблемах, которые оно ему причинило:
«Я назвал эту идею центральной догмой, я подозреваю, по двум причинам. Я уже использовал очевидное слово «гипотеза» в гипотезе последовательности , и вдобавок я хотел предположить, что это новое предположение было более центральным и более мощным. ... Как оказалось, использование слова «догма» причинило больше проблем, чем пользы. Много лет спустя Жак Моно заметил мне, что я, похоже, не понимаю правильного использования слова «догма», которое представляет собой убеждение , которое невозможно. Можно сомневаться , я смутно это понимал, но, поскольку я считал, что все религиозные убеждения не имеют под собой основы, я использовал это слово так, как я сам о нем думал, а не так, как делает большая часть мира, и просто применил его. великая гипотеза, которая, какой бы правдоподобной она ни была, имела мало прямого экспериментального подтверждения».
Подобным образом Хорас Фриланд Джадсон пишет в «Восьмом дне творения» : [19]
«Я считал, что догма — это идея, для которой не было разумных доказательств . Видите?!» И Крик взревел от восторга. «Я просто не знал догма , что означает . И с тем же успехом я мог бы назвать это «Центральной гипотезой» или… вы знаете. Именно это я и хотел сказать. Догма была просто крылатой фразой».
Сравнение с барьером Вейсмана
[ редактировать ]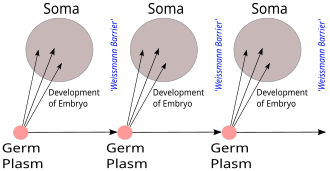
Барьер Вейсмана , предложенный Августом Вейсманом в 1892 году, различает «бессмертные» линии зародышевых клеток ( зародышевую плазму ), которые производят гаметы, и «одноразовые» соматические клетки. Наследственная информация передается только от клеток зародышевой линии к соматическим клеткам (т. е. соматические мутации не наследуются). Это, до открытия роли или структуры ДНК, не предсказывает центральную догму, но предвосхищает ее геноцентричный взгляд на жизнь, хотя и в немолекулярных терминах. [20] [21]
См. также
[ редактировать ]- Жизнь
- Клетка (биология)
- Деление клеток
- Ген
- Экспрессия генов
- Эпигенетика
- Геном
- Альтернативный сплайсинг
- Генетический код
- Рибопереключатель
Ссылки
[ редактировать ]- ^ Ливитт С.А. (июнь 2010 г.). «Расшифровка генетического кода: Маршалл Ниренберг» . Управление истории НИЗ. Архивировано из оригинала 17 марта 2015 г. Проверено 2 марта 2012 г.
- ^ Перейти обратно: а б Кобб М. (сентябрь 2017 г.). «60 лет назад Фрэнсис Крик изменил логику биологии» . ПЛОС Биология . 15 (9): e2003243. дои : 10.1371/journal.pbio.2003243 . ПМК 5602739 . ПМИД 28922352 .
- ^ «Репозиторий архивов CSHL | О синтезе белка» . libgallery.cshl.edu . Проверено 13 ноября 2018 г.
- ^ Крик Ф.Х. (1958). «О синтезе белка». В ФК Сандерс (ред.). Симпозиумы Общества экспериментальной биологии, номер XII: Биологическая репликация макромолекул . Издательство Кембриджского университета. стр. 138–163.
- ^ Крик, Фрэнсис. ХК (1958). «О синтезе белка» . Симпозиумы Общества экспериментальной биологии . 12 . Симпозиумы Общества экспериментальной биологии № XII: Биологическая репликация макромолекул. п. 153. ПМИД 13580867 .
- ^ Перейти обратно: а б Крик Ф. (август 1970 г.). «Центральная догма молекулярной биологии» . Природа . 227 (5258): 561–3. Бибкод : 1970Natur.227..561C . дои : 10.1038/227561a0 . ПМИД 4913914 . S2CID 4164029 .
- ^ Моран Л.А. (15 января 2007 г.). «Песчаная дорожка: основные понятия: центральная догма молекулярной биологии» . sandwalk.blogspot.com . Проверено 17 марта 2018 г.
- ^ Кобб, Мэтью (2015). Величайшая тайна жизни: гонка за расшифровкой генетического кода . Основные книги. ISBN 978-0-465-06267-6 .
Когда Крик сформулировал центральную догму, его целью было не переосмыслить разделение клеток Вейсмана на соматическую линию и зародышевую линию или защитить современное понимание эволюции путем естественного отбора от идеи наследования приобретенных характеристик. Центральная догма основывалась на известных или предполагаемых закономерностях передачи биохимической информации в клетке, а не на какой-либо догматической позиции. По существу, оно было уязвимо для того, чтобы быть признанным недействительным в результате будущих открытий. Тем не менее, в своих основах она оказалась правильной. Реальные или очевидные исключения из этого правила, такие как ретротранскрипционная прионная болезнь или трансгенерационные эпигенетические эффекты, не подорвали его основную истину. (с. 263)
- ^ Баррелл Б.Г., Банкир А.Т., Друэн Дж. (1979). «Другой генетический код в митохондриях человека». Природа . 282 (5735): 189–194. Бибкод : 1979Natur.282..189B . дои : 10.1038/282189a0 . ПМИД 226894 . S2CID 4335828 . ( [1] )
- ^ Эльжановский А, Остелл Дж (07 апреля 2008 г.). «Генетические коды» . Национальный центр биотехнологической информации (NCBI) . Проверено 3 августа 2021 г.
- ^ Алквист П. (май 2002 г.). «РНК-зависимые РНК-полимеразы, вирусы и подавление РНК». Наука . 296 (5571): 1270–3. Бибкод : 2002Sci...296.1270A . дои : 10.1126/science.1069132 . ПМИД 12016304 . S2CID 42526536 .
- ^ Маккарти Би Джей, Холланд Джей Джей (сентябрь 1965 г.). «Денатурированная ДНК как прямая матрица для синтеза белка in vitro» . Труды Национальной академии наук Соединенных Штатов Америки . 54 (3): 880–6. Бибкод : 1965ПНАС...54..880М . дои : 10.1073/pnas.54.3.880 . ПМК 219759 . ПМИД 4955657 .
- ^ . Узава Т., Ямагиши А., Осима Т. (июнь 2002 г.). «Синтез полипептидов, управляемый ДНК как посредником в бесклеточном синтезе полипептидов крайними термофилами, Thermus thermophilus HB27 и штаммом 7 Sulfolobus tokodaii». Журнал биохимии . 131 (6): 849–53. doi : 10.1093/oxfordjournals.jbchem.a003174 . ПМИД 12038981 .
- ^ Буссар А.Е. (август 2005 г.). «Научная революция? Прионная аномалия может бросить вызов центральной догме молекулярной биологии» . Отчеты ЭМБО . 6 (8): 691–4. дои : 10.1038/sj.embor.7400497 . ПМЦ 1369155 . ПМИД 16065057 .
- ^ Кунин Е.В. (август 2012 г.). «Сохраняется ли центральная догма?» . Биология Директ . 7:27 . дои : 10.1186/1745-6150-7-27 . ПМЦ 3472225 . ПМИД 22913395 .
- ^ Ридли Р. (2001). «Что бы Томас Генри Хаксли сделал с прионными болезнями?». В Бейкер HF (ред.). Молекулярная патология прионов . Методы молекулярной медицины. Хумана Пресс. стр. 1–16 . ISBN 0-89603-924-2 .
- ^ Уилкинс А.С. (январь 2012 г.). «(Обзор) Эволюция: взгляд из XXI века» . Геномная биология и эволюция . 4 (4): 423–426. дои : 10.1093/gbe/evs008 . ПМЦ 3342868 .
- ^ Моран Л.А. (май – июнь 2011 г.). «(Обзор) Эволюция: взгляд из XXI века» . Отчеты Национального центра научного образования . 32,3 (9): 1–4. Архивировано из оригинала 15 сентября 2013 г. Проверено 27 октября 2012 г.
- ^ Джадсон Х.Ф. (1996). «Глава 6: Я считал, что догма — это идея, для которой не было разумных доказательств . Видишь?! ». Восьмой день творения: творцы революции в биологии (изд. к 25-летию). Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-477-7 .
- ^ Де Тьеж А., Танге К., Брэкман Дж., Ван де Пер Ю. (январь 2014 г.). «От ДНК-к NA-центризму и новые условия геноцентризма». Биология и философия . 29 (1): 55–69. дои : 10.1007/s10539-013-9393-z . S2CID 85866639 .
- ^ Тернер Дж.С. (2013). Хеннинг Б.Г., Скарф AC (ред.). Второй закон биологии: гомеостаз, цель и желание . Роуман и Литтлфилд. п. 192. ИСБН 978-0-7391-7436-4 .
Там, где Вейсман говорил, что изменения, приобретенные в течение жизни организма, не могут иметь обратную связь с передаваемыми признаками в зародышевой линии, CDMB теперь добавил, что информация, закодированная в белках, не может иметь обратной связи и влиять на генетическую информацию в какой бы то ни было форме. , что, по сути, представляло собой молекулярную переработку барьера Вейсмана.
{{cite book}}:|work=игнорируется ( помогите )
Дальнейшее чтение
[ редактировать ]- Буссар А.Е. (август 2005 г.). «Научная революция? Прионная аномалия может бросить вызов центральной догме молекулярной биологии» . Отчеты ЭМБО . 6 (8): 691–4. дои : 10.1038/sj.embor.7400497 . ПМЦ 1369155 . ПМИД 16065057 .
- Бейкер, Гарри Ф. (2001). Молекулярная патология прионов (методы молекулярной медицины) . Хумана Пресс. ISBN 0-89603-924-2
- Ли Джей Джей, Биггин, доктор медицинских наук (март 2015 г.). «Экспрессия генов. Статистика переоценивает центральную догму» . Наука . 347 (6226): 1066–7. Бибкод : 2015Sci...347.1066L . дои : 10.1126/science.aaa8332 . ПМИД 25745146 .
- Пирас В., Томита М., Сельвараджо К. (2012). «Является ли центральная догма глобальным свойством клеточного потока информации?» . Границы в физиологии . 3 : 439. doi : 10.3389/fphys.2012.00439 . ПМК 3505008 . ПМИД 23189060 .
- Робинсон В.Л. (2009). «Переосмысление центральной догмы: некодирующие РНК биологически значимы». Урологическая онкология . 27 (3): 304–6. дои : 10.1016/j.urolonc.2008.11.004 . ПМИД 19414118 .
Внешние ссылки
[ редактировать ]- Разработка центральной догмы – Scitable: естественное образование
- Анимация Центральной догмы от RIKEN - NatureDocumentaries.org
- Дискуссия о вызовах «центральной догме молекулярной биологии»
- Объяснение центрального догмата с помощью музыкальной аналогии.
- «Фрэнсис Гарри Комптон Крик (1916–2004)» А. Андрея в Энциклопедии Embryo Project.