Протеиногенная аминокислота

Протеиногенные аминокислоты — это аминокислоты , которые биосинтетически включаются в белки во время трансляции . Слово «протеиногенный» означает «создающий белок». На протяжении всей известной жизни существует 22 генетически кодируемые (протеиногенные) аминокислоты, 20 из которых входят в стандартный генетический код и еще 2 ( селеноцистеин и пирролизин ), которые могут быть включены с помощью специальных механизмов трансляции. [1]
Напротив, непротеиногенные аминокислоты — это аминокислоты, которые либо не включены в белки (например, ГАМК , L -ДОФА или трийодтиронин ), неправильно включены вместо генетически кодируемой аминокислоты, либо не производятся напрямую и изолированно стандартными клеточными белками. машины (например, гидроксипролин ). Последнее часто является результатом посттрансляционной модификации белков. Некоторые непротеиногенные аминокислоты включены в нерибосомальные пептиды , которые синтезируются нерибосомальными пептидсинтетазами.
И эукариоты , и прокариоты могут включать селеноцистеин в свои белки посредством нуклеотидной последовательности, известной как элемент SECIS , которая заставляет клетку транслировать ближайший кодон UGA в селеноцистеин (UGA обычно является стоп-кодоном ). У некоторых метаногенных прокариот кодон UAG (обычно стоп-кодон) также может транслироваться в пирролизин . [2]
У эукариот имеется только 21 протеиногенная аминокислота, 20 стандартного генетического кода, плюс селеноцистеин . Люди могут синтезировать 12 из них друг из друга или из других молекул промежуточного метаболизма. Остальные девять необходимо потреблять (обычно в виде их белковых производных), поэтому они называются незаменимыми аминокислотами . Незаменимыми аминокислотами являются гистидин , изолейцин , лейцин , лизин , метионин , фенилаланин , треонин , триптофан и валин (т.е. H, I, L, K, M, F, T, W, V). [3]
Было обнаружено, что протеиногенные аминокислоты относятся к набору аминокислот , которые могут распознаваться системами аутоаминоацилирования рибозимов . [4] Таким образом, непротеиногенные аминокислоты были бы исключены из-за случайного эволюционного успеха форм жизни, основанных на нуклеотидах. Были предложены и другие причины, объясняющие, почему определенные специфические непротеиногенные аминокислоты обычно не включаются в белки; например, орнитин и гомосерин циклизуются против основной цепи пептида и фрагментируют белок с относительно коротким периодом полураспада , в то время как другие токсичны, поскольку могут быть ошибочно включены в белки, такие как аналог аргинина канаванин .
отбор Предполагается , что эволюционный определенных протеиногенных аминокислот из первичного супа произошел из-за их лучшего включения в полипептидную цепь, в отличие от непротеиногенных аминокислот. [5]
Структуры
[ редактировать ]Следующее иллюстрирует структуры и сокращения 21 аминокислоты, которые непосредственно кодируются для синтеза белка генетическим кодом эукариот. Приведенные ниже структуры представляют собой стандартные химические структуры, а не типичные формы цвиттер-ионов , существующие в водных растворах.

 L -Аланин
L -Аланин
(В гостях / А) L -аргинин
L -аргинин
(Злой / Р) L -аспарагин
L -аспарагин
(Асн/Н) L -аспарагиновая кислота
L -аспарагиновая кислота
(Асп/Д) L -цистеин
L -цистеин
(Цис/С) L -глутаминовая кислота
L -глутаминовая кислота
(Клей) L -глютамин
L -глютамин
(Глн/Q) Глицин
Глицин
(Глай/Г) L -гистидин
L -гистидин
(Его/Ч) L -изолейцин
L -изолейцин
(Сколько/я) L -лейцин
L -лейцин
(лей / л) L -лизин
L -лизин
(Свет / К) L -метионин
L -метионин
(Мет/М)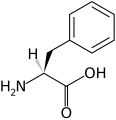 L -фенилаланин
L -фенилаланин
(Пхе/Ф) L -Пролин
L -Пролин
(Про/П) L -серин
L -серин
(Бе/С) L -треонин
L -треонин
(Тр/Т) L -триптофан
L -триптофан
(Трп/Вт) L -тирозин
L -тирозин
(Башня / Y) L -Валин
L -Валин
(Val / V)




















IUPAC / IUBMB теперь также рекомендует стандартные сокращения для следующих двух аминокислот:
 L -селеноцистеин
L -селеноцистеин
(сек/у) L -пирролизин
L -пирролизин
(Pyl / O)

Химические свойства
[ редактировать ]Ниже приводится таблица, в которой перечислены однобуквенные символы, трехбуквенные символы и химические свойства боковых цепей стандартных аминокислот. элементов Перечисленные массы основаны на средневзвешенных значениях изотопов в их естественном содержании . Образование пептидной связи приводит к отщеплению молекулы воды . Следовательно, масса белка равна массе аминокислот, из которых он состоит, минус 18,01524 Да на пептидную связь.
Общие химические свойства
[ редактировать ]| Аминокислота | Короткий | Сокращение. | Среднее масса ( Да ) | пи | ПК 1 (α-COO - ) | ПК 2 (α-NH 3 + ) |
|---|---|---|---|---|---|---|
| Аланин | А | Земля | 89.09404 | 6.01 | 2.35 | 9.87 |
| Цистеин | С | Цис | 121.15404 | 5.05 | 1.92 | 10.70 |
| Аспарагиновая кислота | Д | Асп | 133.10384 | 2.85 | 1.99 | 9.90 |
| Глутаминовая кислота | И | Глу | 147.13074 | 3.15 | 2.10 | 9.47 |
| Фенилаланин | Ф | Пхе | 165.19184 | 5.49 | 2.20 | 9.31 |
| Глицин | Г | Гли | 75.06714 | 6.06 | 2.35 | 9.78 |
| Гистидин | ЧАС | Его | 155.15634 | 7.60 | 1.80 | 9.33 |
| изолейцин | я | С | 131.17464 | 6.05 | 2.32 | 9.76 |
| Лизин | К | Свет | 146.18934 | 9.60 | 2.16 | 9.06 |
| Лейцин | л | Лео | 131.17464 | 6.01 | 2.33 | 9.74 |
| Метионин | М | Из | 149.20784 | 5.74 | 2.13 | 9.28 |
| Аспарагин | Н | Асн | 132.11904 | 5.41 | 2.14 | 8.72 |
| Пирролизин | ТО | Pyl | 255.31 | ? | ? | ? |
| Пролин | П | Про | 115.13194 | 6.30 | 1.95 | 10.64 |
| Глютамин | вопрос | Глн | 146.14594 | 5.65 | 2.17 | 9.13 |
| Аргинин | Р | Арг | 174.20274 | 10.76 | 1.82 | 8.99 |
| Серин | С | Быть | 105.09344 | 5.68 | 2.19 | 9.21 |
| Треонин | Т | чр | 119.12034 | 5.60 | 2.09 | 9.10 |
| Селеноцистеин | В | сек. | 168.053 | 5.47 | 1.91 | 10 |
| Валин | V | Вал | 117.14784 | 6.00 | 2.39 | 9.74 |
| Триптофан | В | Трп | 204.22844 | 5.89 | 2.46 | 9.41 |
| Тирозин | И | Тир | 181.19124 | 5.64 | 2.20 | 9.21 |
Свойства боковой цепи
[ редактировать ]| Аминокислота | Короткий | Сокращение. | Боковая цепь | Гидро- фобический | пКа § | Полярный | рН | Маленький | Крошечный | Ароматный или алифатический | Ван дер Ваальс объем (Å 3 ) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Аланин | А | Земля | -CH-CH3 | - | - | Алифатический | 67 | ||||
| Цистеин | С | Цис | -СН 2 Ш | 8.55 | кислый | - | 86 | ||||
| Аспарагиновая кислота | Д | Асп | -СН 2 СООН | 3.67 | кислый | - | 91 | ||||
| Глутаминовая кислота | И | Глу | -СН 2 СН 2 СООН | 4.25 | кислый | - | 109 | ||||
| Фенилаланин | Ф | Пхе | -СН 2 С 6 Н 5 | - | - | Ароматный | 135 | ||||
| Глицин | Г | Гли | -ЧАС | - | - | - | 48 | ||||
| Гистидин | ЧАС | Его | -СН 2 - С 3 Н 3 Н 2 | 6.54 | слабый базовый | Ароматный | 118 | ||||
| изолейцин | я | С | -СН(СН 3 )СН 2 СН 3 | - | - | Алифатический | 124 | ||||
| Лизин | К | Свет | -(СН 2 ) 4 NH 2 | 10.40 | базовый | - | 135 | ||||
| Лейцин | л | Лео | -СН 2 СН(СН 3 ) 2 | - | - | Алифатический | 124 | ||||
| Метионин | М | Из | -СН 2 СН 2 С СН 3 | - | - | Алифатический | 124 | ||||
| Аспарагин | Н | Асн | -СН 2 КОНН 2 | - | - | - | 96 | ||||
| Пирролизин | ТО | Pyl | -(CH 2 ) 4 NHCO C 4 H 5 N CH 3 | без даты | слабый базовый | - | ? | ||||
| Пролин | П | Про | -СН 2 СН 2 СН 2 - | - | - | - | 90 | ||||
| Глютамин | вопрос | Глн | -СН 2 СН 2 КОНН 2 | - | - | - | 114 | ||||
| Аргинин | Р | Арг | -(СН 2 ) 3 NH-C(NH)NH 2 | 12.3 | строго базовый | - | 148 | ||||
| Серин | С | Быть | -СН 2 ОН | - | - | - | 73 | ||||
| Треонин | Т | чр | -СН(ОН) СН3 | - | - | - | 93 | ||||
| Селеноцистеин | В | сек. | -CH 2 SeH | 5.43 | кислый | - | ? | ||||
| Валин | V | Вал | -СН( СН3 ) 2 | - | - | Алифатический | 105 | ||||
| Триптофан | В | Трп | -СН 2 С 8 Н 6 Н | - | - | Ароматный | 163 | ||||
| Тирозин | И | Тир | -СН 2 -С 6 Н 4 ОН | 9.84 | слабокислый | Ароматный | 141 |
§: Значения Asp, Cys, Glu, His, Lys и Tyr были определены с использованием аминокислотного остатка, расположенного в центре пентапептида аланина. [6] Значение Arg взято из Pace et al. (2009). [7] Значение секунд взято из Byun & Kang (2011). [8]
Н.Д.: Значение pKa пирролизина не сообщалось.
Примечание. Значение pKa аминокислотного остатка в небольшом пептиде обычно немного отличается, когда он находится внутри белка. Расчеты рКа белка иногда используются для расчета изменения значения рКа аминокислотного остатка в этой ситуации.
Экспрессия генов и биохимия
[ редактировать ]| Аминокислота | Короткий | Сокращение. | Кодон (ы) | возникновение | Незаменим ‡ у человека | |||
|---|---|---|---|---|---|---|---|---|
| в архейских белках (%) & | в белках бактерий (%) & | в белках эукариот (%) & | в белках человека (%) & | |||||
| Аланин | А | Земля | GCU, GCC, GCA, GCG | 8.2 | 10.06 | 7.63 | 7.01 | Нет |
| Цистеин | С | Цис | УГУ, УГК | 0.98 | 0.94 | 1.76 | 2.3 | Условно |
| Аспарагиновая кислота | Д | Асп | ГАУ, ГАЦ | 6.21 | 5.59 | 5.4 | 4.73 | Нет |
| Глутаминовая кислота | И | Глу | ГАА, ГАГ | 7.69 | 6.15 | 6.42 | 7.09 | Условно |
| Фенилаланин | Ф | Пхе | УУУ, УУК | 3.86 | 3.89 | 3.87 | 3.65 | Да |
| Глицин | Г | Гли | ГГУ, ГГК, ГГА, ГГГ | 7.58 | 7.76 | 6.33 | 6.58 | Условно |
| Гистидин | ЧАС | Его | КАУ, САС | 1.77 | 2.06 | 2.44 | 2.63 | Да |
| изолейцин | я | С | ОМГ, ОМГ, ОМГ | 7.03 | 5.89 | 5.1 | 4.33 | Да |
| Лизин | К | Свет | ААА, ААГ | 5.27 | 4.68 | 5.64 | 5.72 | Да |
| Лейцин | л | Лео | УУА, УУГ, КУУ, КУК, КУА, КУГ | 9.31 | 10.09 | 9.29 | 9.97 | Да |
| Метионин | М | Из | АВГ | 2.35 | 2.38 | 2.25 | 2.13 | Да |
| Аспарагин | Н | Асн | ААУ, ААК | 3.68 | 3.58 | 4.28 | 3.58 | Нет |
| Пирролизин | ТО | Pyl | УАГ * | 0 | 0 | 0 | 0 | Нет |
| Пролин | П | Про | CCU, CCC, CCA, CCG | 4.26 | 4.61 | 5.41 | 6.31 | Нет |
| Глютамин | вопрос | Глн | САА, ЦАГ | 2.38 | 3.58 | 4.21 | 4.77 | Нет |
| Аргинин | Р | Арг | CGU, CGC, CGA, CGG, AGA, AGG | 5.51 | 5.88 | 5.71 | 5.64 | Условно |
| Серин | С | Быть | УКУ, УКК, УЦА, UCG, AGU, AGC | 6.17 | 5.85 | 8.34 | 8.33 | Нет |
| Треонин | Т | чр | АКУ, АКК, АСА, АЧГ | 5.44 | 5.52 | 5.56 | 5.36 | Да |
| Селеноцистеин | В | сек. | УГА ** | 0 | 0 | 0 | >0 | Нет |
| Валин | V | Вал | ГУУ, ГУК, ГУА, ГУГ | 7.8 | 7.27 | 6.2 | 5.96 | Да |
| Триптофан | В | Трп | Угги | 1.03 | 1.27 | 1.24 | 1.22 | Да |
| Тирозин | И | Тир | УАУ, ОАК | 3.35 | 2.94 | 2.87 | 2.66 | Условно |
| Стоп-кодон † | - | Срок | УАА, УАГ, УГА †† | ? | ? | ? | — | — |
* UAG обычно представляет собой янтарный стоп-кодон , но в организмах, содержащих биологический механизм, кодируемый кластером генов pylTSBCD, будет включена аминокислота пирролизин. [9]
** UGA обычно представляет собой стоп-кодон опала (или умбры), но кодирует селеноцистеин, если элемент SECIS . присутствует
† Стоп -кодон не является аминокислотой, но включен для полноты описания.
†† UAG и UGA не всегда действуют как стоп-кодоны (см. выше).
‡ Незаменимая аминокислота не может синтезироваться в организме человека и поэтому должна поступать с пищей. Условно незаменимые аминокислоты обычно не требуются в рационе, но должны поставляться экзогенно определенным группам населения, которые не синтезируют их в адекватных количествах.
& Встречаемость аминокислот основана на протеомах 135 архей, 3775 бактерий, 614 протеомов эукариот и протеоме человека (21 006 белков) соответственно. [10]
Масс-спектрометрия
[ редактировать ]В масс-спектрометрии пептидов и белков полезно знание масс остатков. Масса пептида или белка представляет собой сумму масс остатков плюс массу воды ( моноизотопная масса = 18,01056 Да; средняя масса = 18,0153 Да). Массы остатков рассчитывают по табличным химическим формулам и атомным весам. [11] В масс-спектрометрии ионы также могут включать один или несколько протонов ( моноизотопная масса = 1,00728 Да; средняя масса* = 1,0074 Да). *Протоны не могут иметь среднюю массу, это сбивает с толку вывод о том, что дейтроны являются действительным изотопом, но они должны принадлежать к другому виду (см. Гидрон (химия) ).
| Аминокислота | Короткий | Сокращение. | Формула | Мой. масса § ( Да ) | Среднее масса ( Да ) |
|---|---|---|---|---|---|
| Аланин | А | Земля | С 3 Ч 5 НЕТ | 71.03711 | 71.0779 |
| Цистеин | С | Цис | С 3 Н 5 НОС | 103.00919 | 103.1429 |
| Аспарагиновая кислота | Д | Асп | С 4 Ч 5 НЕТ 3 | 115.02694 | 115.0874 |
| Глутаминовая кислота | И | Глу | С 5 Ч 7 НЕТ 3 | 129.04259 | 129.1140 |
| Фенилаланин | Ф | Пхе | C9HC9H9NO NO | 147.06841 | 147.1739 |
| Глицин | Г | Гли | С 2 Ч 3 НЕТ | 57.02146 | 57.0513 |
| Гистидин | ЧАС | Его | C6H7NC6H7N3O O | 137.05891 | 137.1393 |
| изолейцин | я | С | С 6 Ч 11 НЕТ | 113.08406 | 113.1576 |
| Лизин | К | Свет | С 6 Н 12 Н 2 О | 128.09496 | 128.1723 |
| Лейцин | л | Лео | С 6 Ч 11 НЕТ | 113.08406 | 113.1576 |
| Метионин | М | Из | С 5 Ч 9 США | 131.04049 | 131.1961 |
| Аспарагин | Н | Асн | С 4 Н 6 Н 2 О 2 | 114.04293 | 114.1026 |
| Пирролизин | ТО | Pyl | С 12 Н 19 Н 3 О 2 | 237.14773 | 237.2982 |
| Пролин | П | Про | С 5 Ч 7 НЕТ | 97.05276 | 97.1152 |
| Глютамин | вопрос | Глн | C5H8N2OC5H8N2O2 | 128.05858 | 128.1292 |
| Аргинин | Р | Арг | С 6 Н 12 Н 4 О | 156.10111 | 156.1857 |
| Серин | С | Быть | С 3 Ч 5 НЕТ 2 | 87.03203 | 87.0773 |
| Треонин | Т | чр | C4H7NOC4H7NO2 | 101.04768 | 101.1039 |
| Селеноцистеин | В | сек. | С 3 Н 5 НОС | 150.95364 | 150.0489 |
| Валин | V | Вал | С 5 Ч 9 НЕТ | 99.06841 | 99.1311 |
| Триптофан | В | Трп | С 11 Н 10 Н 2 О | 186.07931 | 186.2099 |
| Тирозин | И | Тир | C9H9NOC9H9NO2 | 163.06333 | 163.1733 |
Стехиометрия и метаболические затраты в клетке
[ редактировать ]В таблице ниже указано содержание аминокислот в клетках E.coli и метаболические затраты (АТФ) на синтез аминокислот. Отрицательные числа указывают на то, что метаболические процессы являются энергетически выгодными и не требуют затрат чистого АТФ клетки. [12] В состав аминокислот входят аминокислоты в свободной форме и в полимеризационной форме (белки).
| Аминокислота | Короткий | Сокращение. | Избыток (количество молекул (×10 8 ) для клеток E. coli ) | Стоимость АТФ в синтезе | |
|---|---|---|---|---|---|
| Аэробный условия | Анаэробный условия | ||||
| Аланин | А | Земля | 2.9 | -1 | 1 |
| Цистеин | С | Цис | 0.52 | 11 | 15 |
| Аспарагиновая кислота | Д | Асп | 1.4 | 0 | 2 |
| Глутаминовая кислота | И | Глу | 1.5 | -7 | -1 |
| Фенилаланин | Ф | Пхе | 1.1 | -6 | 2 |
| Глицин | Г | Гли | 3.5 | -2 | 2 |
| Гистидин | ЧАС | Его | 0.54 | 1 | 7 |
| изолейцин | я | С | 1.7 | 7 | 11 |
| Лизин | К | Свет | 2.0 | 5 | 9 |
| Лейцин | л | Лео | 2.6 | -9 | 1 |
| Метионин | М | Из | 0.88 | 21 | 23 |
| Аспарагин | Н | Асн | 1.4 | 3 | 5 |
| Пирролизин | ТО | Pyl | - | - | - |
| Пролин | П | Про | 1.3 | -2 | 4 |
| Глютамин | вопрос | Глн | 1.5 | -6 | 0 |
| Аргинин | Р | Арг | 1.7 | 5 | 13 |
| Серин | С | Быть | 1.2 | -2 | 2 |
| Треонин | Т | чр | 1.5 | 6 | 8 |
| Селеноцистеин | В | сек. | - | - | - |
| Валин | V | Вал | 2.4 | -2 | 2 |
| Триптофан | В | Трп | 0.33 | -7 | 7 |
| Тирозин | И | Тир | 0.79 | -8 | 2 |
Примечания
[ редактировать ]| Аминокислота | Сокращение. | Примечания | |
|---|---|---|---|
| Аланин | А | Земля | Он очень распространен и очень универсален, он более жесткий, чем глицин, но достаточно мал, чтобы устанавливать лишь небольшие стерические ограничения для конформации белка. Он ведет себя достаточно нейтрально и может располагаться как в гидрофильных участках белка снаружи, так и в гидрофобных участках внутри. |
| Аспарагин или аспарагиновая кислота | Б | Аскс | Заполнитель, когда любая аминокислота может занимать позицию |
| Цистеин | С | Цис | Атом серы легко связывается с ионами тяжелых металлов . В окислительных условиях два цистеина могут образовать дисульфидную связь с образованием аминокислоты цистин . Когда цистины являются частью белка, , инсулина например , третичная структура стабилизируется, что делает белок более устойчивым к денатурации ; следовательно, дисульфидные связи часто встречаются в белках, которым приходится функционировать в суровых условиях окружающей среды, включая пищеварительные ферменты (например, пепсин и химотрипсин ) и структурные белки (например, кератин ). Дисульфиды также содержатся в пептидах, слишком маленьких, чтобы самостоятельно сохранять стабильную форму (например, в инсулине ). |
| Аспарагиновая кислота | Д | Асп | Asp ведет себя аналогично глутаминовой кислоте и несет гидрофильную кислотную группу с сильным отрицательным зарядом. Обычно он расположен на внешней поверхности белка, что делает его водорастворимым. Он связывается с положительно заряженными молекулами и ионами и часто используется в ферментах для фиксации ионов металлов. Находясь внутри белка, аспартат и глутамат обычно сочетаются с аргинином и лизином. |
| Глутаминовая кислота | И | Глу | Glu ведет себя аналогично аспарагиновой кислоте и имеет более длинную и немного более гибкую боковую цепь. |
| Фенилаланин | Ф | Пхе | Важные для человека фенилаланин, тирозин и триптофан содержат большую жесткую ароматическую группу на боковой цепи. Это самые большие аминокислоты. Подобно изолейцину, лейцину и валину, они гидрофобны и имеют тенденцию ориентироваться внутрь свернутой белковой молекулы. Фенилаланин может превращаться в тирозин. |
| Глицин | Г | Гли | Из-за двух атомов водорода у α-углерода глицин не является оптически активным . Это самая маленькая аминокислота, она легко вращается и придает гибкость белковой цепи. Он способен проникать в самые узкие места, например, в тройную спираль коллагена . Поскольку слишком большая гибкость обычно нежелательна, в качестве структурного компонента он встречается реже, чем аланин. |
| Гистидин | ЧАС | Его | Он необходим для человека. Даже в слабокислых условиях происходит протонирование азота, изменяющее свойства гистидина и полипептида в целом. Он используется многими белками в качестве регуляторного механизма, изменяя конформацию и поведение полипептида в кислых областях, таких как поздняя эндосома или лизосома , вызывая изменение конформации ферментов. Однако для этого необходимо лишь несколько гистидинов, поэтому их сравнительно мало. |
| изолейцин | я | С | Иль необходим человеку. Изолейцин, лейцин и валин имеют большие алифатические гидрофобные боковые цепи. Их молекулы жесткие, а их взаимные гидрофобные взаимодействия важны для правильного сворачивания белков, поскольку эти цепи имеют тенденцию располагаться внутри белковой молекулы. |
| Лейцин или изолейцин | Дж | Xle | Заполнитель, когда любая аминокислота может занимать позицию |
| Лизин | К | Свет | Лиз необходим человеку и ведет себя аналогично аргинину. Он содержит длинную гибкую боковую цепь с положительно заряженным концом. Гибкость цепи делает лизин и аргинин пригодными для связывания с молекулами с множеством отрицательных зарядов на поверхности. Например, активные области ДНК -связывающих белков богаты аргинином и лизином. Благодаря сильному заряду эти две аминокислоты склонны располагаться на внешних гидрофильных поверхностях белков; когда они обнаруживаются внутри, они обычно соединяются с соответствующей отрицательно заряженной аминокислотой, например, аспартатом или глутаматом. |
| Лейцин | л | Лео | Лей необходим человеку и ведет себя аналогично изолейцину и валину. |
| Метионин | М | Из | Мет необходим для человека. Всегда первая аминокислота, которая включается в белок, иногда она удаляется после трансляции. Как и цистеин, он содержит серу, но с метильной группой вместо водорода. Эту метильную группу можно активировать, и она используется во многих реакциях, когда новый атом углерода добавляется к другой молекуле. |
| Аспарагин | Н | Асн | Подобно аспарагиновой кислоте, Asn содержит амидную группу, в которой Asp имеет карбоксильную группу . |
| Пирролизин | ТО | Pyl | Похож на лизин , но имеет пирролиновое кольцо. |
| Пролин | П | Про | Pro содержит необычное кольцо у N-концевой аминогруппы, которое приводит амидную последовательность CO-NH в фиксированную конформацию. Он может разрушать структуры сворачивания белка, такие как α-спираль или β-лист , вызывая желаемый излом в белковой цепи. Обычный для коллагена , он часто подвергается посттрансляционной модификации до гидроксипролина . |
| Глютамин | вопрос | Глн | Подобно глутаминовой кислоте, Gln содержит амидную группу, где Glu имеет карбоксильную группу . Используемая в белках и в качестве хранилища аммиака , это самая распространенная аминокислота в организме. |
| Аргинин | Р | Арг | Функционально аналогичен лизину. |
| Серин | С | Быть | Серин и треонин имеют короткую группу, оканчивающуюся гидроксильной группой. Его водород легко удалить, поэтому серин и треонин часто выступают донорами водорода в ферментах. Оба они очень гидрофильны, поэтому внешние области растворимых белков, как правило, богаты ими. |
| Треонин | Т | чр | Важный для человека Thr ведет себя аналогично серину. |
| Селеноцистеин | В | сек. | Селеновый селен аналог цистеина, в котором заменяет атом серы . |
| Валин | V | Вал | Важный для человека, Вал ведет себя аналогично изолейцину и лейцину. |
| Триптофан | В | Трп | Важный для человека Trp ведет себя аналогично фенилаланину и тирозину. Это предшественник серотонина и естественно флуоресцентный . |
| Неизвестный | Х | Да | Заполнитель, когда аминокислота неизвестна или не важна. |
| Тирозин | И | Тир | Тир ведет себя аналогично фенилаланину (предшественнику тирозина) и триптофану и является предшественником меланина , адреналина и гормонов щитовидной железы . Естественно флуоресцентный , его флуоресценция обычно гасится за счет передачи энергии триптофанам. |
| Глутаминовая кислота или глутамин | С | Глкс | Заполнитель, когда любая аминокислота может занимать позицию |

Катаболизм
[ редактировать ]Аминокислоты можно классифицировать по свойствам их основных продуктов: [13]
- Глюкогенный, продукты которого способны образовывать глюкозу путем глюконеогенеза.
- Кетогенные продукты, не обладающие способностью образовывать глюкозу: эти продукты все еще можно использовать для кетогенеза или синтеза липидов .
- Аминокислоты катаболизируются как в глюкогенные, так и в кетогенные продукты.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Амброгелли А., Палиура С., Зёлль Д. (январь 2007 г.). «Естественное расширение генетического кода». Химическая биология природы . 3 (1): 29–35. дои : 10.1038/nchembio847 . ПМИД 17173027 .
- ^ Лобанов А.В., Туранов А.А., Хэтфилд Д.Л., Гладышев В.Н. (август 2010 г.). «Двойные функции кодонов в генетическом коде» . Критические обзоры по биохимии и молекулярной биологии . 45 (4): 257–65. дои : 10.3109/10409231003786094 . ПМК 3311535 . ПМИД 20446809 .
- ^ Молодой VR (август 1994 г.). «Потребности взрослых в аминокислотах: необходимость серьезного пересмотра текущих рекомендаций» (PDF) . Журнал питания . 124 (8 дополнений): 1517S–1523S. дои : 10.1093/jn/124.suppl_8.1517S . ПМИД 8064412 .
- ^ Эривес А (август 2011 г.). «Модель ферментов протоантикодонов РНК, требующих гомохиральности L-аминокислот» . Журнал молекулярной эволюции . 73 (1–2): 10–22. Бибкод : 2011JMolE..73...10E . дои : 10.1007/s00239-011-9453-4 . ПМЦ 3223571 . ПМИД 21779963 .
- ^ Френкель-Пинтер, Моран; Хейнс, Джей В.; С, Мартин; Петров Антон С.; Буркар, Брэдли Т.; Кришнамурти, Раманарайан; Худ, Николас В.; Леман, Люк Дж.; Уильямс, Лорен Дин (13 августа 2019 г.). «Селективное включение белковых катионных аминокислот по сравнению с небелковыми в модельных реакциях пребиотической олигомеризации» . Труды Национальной академии наук . 116 (33): 16338–16346. Бибкод : 2019PNAS..11616338F . дои : 10.1073/pnas.1904849116 . ISSN 0027-8424 . ПМК 6697887 . ПМИД 31358633 .
- ^ Теркилл Р.Л., Гримсли Г.Р., Шольц Дж.М., Пейс К.Н. (май 2006 г.). «Значения pK ионизируемых групп белков» . Белковая наука . 15 (5): 1214–8. дои : 10.1110/ps.051840806 . ПМЦ 2242523 . ПМИД 16597822 .
- ^ Пейс С.Н., Гримсли Г.Р., Шольц Дж.М. (май 2009 г.). «Ионизируемые группы белков: значения pK и их вклад в стабильность и растворимость белка» . Журнал биологической химии . 284 (20): 13285–9. дои : 10.1074/jbc.R800080200 . ПМЦ 2679426 . ПМИД 19164280 .
- ^ Бён Б.Дж., Кан Ю.К. (май 2011 г.). «Конформационные предпочтения и значение pK (a) остатка селеноцистеина». Биополимеры . 95 (5): 345–53. дои : 10.1002/bip.21581 . ПМИД 21213257 . S2CID 11002236 .
- ^ Ротер М., Кшицкий Дж. А. (август 2010 г.). «Селеноцистеин, пирролизин и уникальный энергетический метаболизм метаногенных архей» . Архея . 2010 : 1–14. дои : 10.1155/2010/453642 . ПМЦ 2933860 . ПМИД 20847933 .
- ^ Козловский Л.П. (январь 2017 г.). «Протеом-pI: база данных изоэлектрических точек протеома» . Исследования нуклеиновых кислот . 45 (Д1): Д1112–Д1116. дои : 10.1093/nar/gkw978 . ПМК 5210655 . ПМИД 27789699 .
- ^ «Атомный вес и изотопный состав всех элементов» . НИСТ . Проверено 12 декабря 2016 г.
- ^ Филлипс Р., Кондев Дж., Териот Дж., Гарсия Х.Г., Орм Н. (2013). Физическая биология клетки (Второе изд.). Гирляндная наука. п. 178. ИСБН 978-0-8153-4450-6 .
- ^ Ферье Д.Р. (2005). «Глава 20: Разложение и синтез аминокислот». В Champe PC, Харви Р.А., Ферье Д.Р. (ред.). Иллюстрированные обзоры Липпинкотта: биохимия (Иллюстрированные обзоры Липпинкотта) . Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-2265-0 .
Общие ссылки
[ редактировать ]- Нельсон, Дэвид Л.; Кокс, Майкл М. (2000). Ленингерские принципы биохимии (3-е изд.). Стоит издательства. ISBN 978-1-57259-153-0 .
- Кайт Дж., Дулитл РФ (май 1982 г.). «Простой метод отображения гидропатического характера белка». Журнал молекулярной биологии . 157 (1): 105–32. CiteSeerX 10.1.1.458.454 . дои : 10.1016/0022-2836(82)90515-0 . ПМИД 7108955 .
- Мейеренрих, Уве Дж. (2008). Аминокислоты и асимметрия жизни (1-е изд.). Спрингер. ISBN 978-3-540-76885-2 .
- Биохимия, Харперс (2015). Harpers Illustrated Biochemistry (30-е изд.). Ланге. ISBN 978-0-07-182534-4 .
Внешние ссылки
[ редактировать ]| Общие темы |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| По свойствам |
| ||||||||||