Гистон H1
| линкерные гистоны семейства H1 и H5 | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | Linker_histone | ||
| Пфам | PF00538 | ||
| ИнтерПро | ИПР005818 | ||
| УМНЫЙ | SM00526 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1ч / СКОПе / СУПФАМ | ||
| |||
Гистон H1 — одно из пяти основных семейств гистоновых белков которые являются компонентами хроматина эукариотических , клеток. Хотя он высококонсервативен , он, тем не менее, является наиболее изменчивым по последовательности гистоном у разных видов.
Структура
[ редактировать ]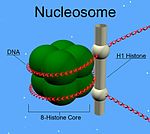
Белки H1 многоклеточных животных имеют центральный глобулярный домен «крылатой спирали» и длинные С- и короткие N-концевые хвосты. H1 занимается упаковкой подструктур «бусинок на веревке» в структуру высокого порядка, детали которой еще не решены. [ 1 ] H1, обнаруженный у протистов и бактерий, иначе известный как нуклеопротеины HC1 и HC2 ( Pfam PF07432 , PF07382 ), лишен центрального домена и N-концевого хвоста. [ 2 ]
H1 менее консервативен, чем коровые гистоны. Глобулярный домен является наиболее консервативной частью H1. [ 3 ]
Функция
[ редактировать ]В отличие от других гистонов, H1 не образует нуклеосомную «бусину». Вместо этого он располагается поверх структуры, удерживая на месте ДНК, обернутую вокруг нуклеосомы. H1 присутствует в половине количества других четырех гистонов, которые вносят две молекулы в каждую нуклеосомную бусину. Помимо связывания с нуклеосомой, белок H1 связывается с областью «линкерной ДНК» (длиной примерно 20–80 нуклеотидов) между нуклеосомами, помогая стабилизировать зигзагообразные волокна хроматина длиной 30 нм. [ 4 ] Многое было изучено о гистоне H1 в результате исследований очищенных волокон хроматина . Ионная экстракция линкерных гистонов из нативного или восстановленного хроматина способствует его разворачиванию в гипотонических условиях от волокон шириной 30 нм до массивов нуклеосом типа «бусинки на нити». [ 5 ] [ 6 ] [ 7 ]
Неясно, способствует ли H1 соленоидоподобному волокну хроматина, в котором открытая линкерная ДНК укорачивается, или он просто способствует изменению угла соседних нуклеосом, не влияя на длину линкера. [ 8 ] Однако было продемонстрировано, что линкерные гистоны способствуют уплотнению волокон хроматина, которые были восстановлены in vitro с использованием синтетических массивов ДНК сильного позиционирующего элемента нуклеосомы «601». [ 9 ] Эксперименты по расщеплению нуклеаз и следам ДНК позволяют предположить, что глобулярный домен гистона H1 локализуется вблизи нуклеосомной диады, где он защищает примерно 15-30 пар оснований дополнительной ДНК. [ 10 ] [ 11 ] [ 12 ] [ 13 ] Кроме того, эксперименты с восстановленным хроматином выявили характерный стеблевой мотив диады в присутствии H1. [ 14 ] Несмотря на пробелы в нашем понимании, появилась общая модель, в которой глобулярный домен H1 закрывает нуклеосому путем сшивания входящей и исходящей ДНК, в то время как хвост связывается с линкерной ДНК и нейтрализует ее отрицательный заряд. [ 8 ] [ 12 ]
Многие эксперименты, посвященные функции H1, были проведены на очищенном, обработанном хроматине в условиях с низким содержанием соли, но роль H1 in vivo менее определена. Клеточные исследования показали, что сверхэкспрессия H1 может вызывать аберрантную морфологию ядра и структуру хроматина и что H1 может служить как положительным, так и отрицательным регулятором транскрипции, в зависимости от гена. [ 15 ] [ 16 ] [ 17 ] В экстрактах яиц Xenopus истощение линкерных гистонов вызывает ~2-кратное удлинение митотических хромосом в длину, тогда как сверхэкспрессия приводит к гиперкомпактированию хромосом в неразделимую массу. [ 18 ] [ 19 ] Полного нокаута H1 in vivo не удалось достичь у многоклеточных организмов из-за существования множества изоформ, которые могут присутствовать в нескольких кластерах генов, но различные изоформы линкерных гистонов в разной степени истощены у Tetrahymena , C. elegans, Arabidopsis, фруктов. мух и мышей, что приводит к различным организмоспецифичным дефектам морфологии ядра, структуры хроматина, метилирования ДНК и/или специфической экспрессии генов. [ 20 ] [ 21 ] [ 22 ]
Динамика
[ редактировать ]Хотя большая часть гистона H1 в ядре связана с хроматином, молекулы H1 перемещаются между областями хроматина с довольно высокой скоростью. [ 23 ] [ 24 ]
Трудно понять, как такой динамический белок мог быть структурным компонентом хроматина, но было высказано предположение, что устойчивое равновесие внутри ядра по-прежнему сильно благоприятствует ассоциации между H1 и хроматином, а это означает, что, несмотря на его динамику, подавляющее большинство H1 в любой данный момент времени связан с хроматином. [ 25 ] H1 уплотняет и стабилизирует ДНК под действием силы и во время сборки хроматина, что позволяет предположить, что динамическое связывание H1 может обеспечить защиту ДНК в ситуациях, когда нуклеосомы необходимо удалить. [ 26 ]
Цитоплазматические факторы, по-видимому, необходимы для динамического обмена гистона H1 на хроматине, но они еще не идентифицированы конкретно. [ 27 ] Динамика H1 может в некоторой степени опосредоваться O-гликозилированием и фосфорилированием. О-гликозилирование H1 может способствовать конденсации и уплотнению хроматина. Было показано, что фосфорилирование во время интерфазы снижает сродство H1 к хроматину и может способствовать деконденсации хроматина и активной транскрипции. Однако было показано, что во время митоза фосфорилирование увеличивает сродство H1 к хромосомам и, следовательно, способствует конденсации митотических хромосом. [ 19 ]
Изоформы
[ редактировать ]Семейство H1 у животных включает несколько изоформ H1, которые могут экспрессироваться в разных или перекрывающихся тканях и стадиях развития в одном организме. Причина появления этих множественных изоформ остается неясной, но как их эволюционная консервативность от морского ежа до человека, так и значительные различия в их аминокислотных последовательностях позволяют предположить, что они функционально не эквивалентны. [ 28 ] [ 29 ] [ 3 ] Одной из изоформ является гистон H5 , который встречается только в эритроцитах птиц , которые отличаются от эритроцитов млекопитающих тем, что у них есть ядра . Другой изоформой является ооцит/ зиготическая изоформа H1M (также известная как B4 или H1foo), обнаруженная у морских ежей, лягушек, мышей и людей, которая заменяется в эмбрионе соматическими изоформами H1A-E и H10, напоминающими H5. [ 3 ] [ 30 ] [ 31 ] [ 32 ] Несмотря на то, что H1M имеет больше отрицательных зарядов, чем соматические изоформы, он с более высоким сродством связывается с митотическими хромосомами в Xenopus . экстрактах яиц [ 19 ]
Посттрансляционные модификации
[ редактировать ]Как и другие гистоны, семейство гистонов H1 подвергается значительной посттрансляционной модификации (PTM). Это включает фосфорилирование серина и треонина, ацетилирование лизина, метилирование лизина и убиквитинирование. [ 33 ] Эти ПТМ выполняют множество функций, но менее изучены, чем ПТМ других гистонов.
См. также
[ редактировать ]- нуклеосома
- Гистон
- хроматин
- Варианты линкерного гистона H1
- Другие гистоны белки- , участвующие в хроматине:
Ссылки
[ редактировать ]- ^ Рамакришнан В., Финч Дж.Т., Грациано В., Ли П.Л., Свит Р.М. (март 1993 г.). «Кристаллическая структура глобулярного домена гистона H5 и ее значение для связывания нуклеосом». Природа . 362 (6417): 219–23. Бибкод : 1993Natur.362..219R . дои : 10.1038/362219a0 . ПМИД 8384699 . S2CID 4301198 .
- ^ Касинский Х.Э., Льюис Дж.Д., Дакс Дж.Б., Аусио Дж. (январь 2001 г.). «Происхождение гистонов линкера H1» . Журнал ФАСЭБ . 15 (1): 34–42. дои : 10.1096/fj.00-0237рев . ПМИД 11149891 . S2CID 10089116 .
- ^ Jump up to: а б с Иззо А., Камениарц К., Шнайдер Р. (апрель 2008 г.). «Семейство гистонов H1: конкретные члены, конкретные функции?» . Биологическая химия . 389 (4): 333–43. дои : 10.1515/BC.2008.037 . ПМИД 18208346 . S2CID 1516241 .
- ^ Чон, Кван В.; Березней, Рональд (1995). Структурная и функциональная организация ядерного матрикса . Бостон: Академическая пресса. стр. 214–7 . ISBN 978-0-12-364565-4 .
- ^ Финч Дж. Т., Клуг А. (июнь 1976 г.). «Соленоидальная модель суперструктуры хроматина» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (6): 1897–901. Бибкод : 1976PNAS...73.1897F . дои : 10.1073/pnas.73.6.1897 . ПМК 430414 . ПМИД 1064861 .
- ^ Тома Ф., Коллер Т. (сентябрь 1977 г.). «Влияние гистона H1 на структуру хроматина» (PDF) . Клетка . 12 (1): 101–7. дои : 10.1016/0092-8674(77)90188-X . ПМИД 561660 . S2CID 11155120 .
- ^ Тома Ф., Коллер Т., Клуг А. (ноябрь 1979 г.). «Участие гистона H1 в организации нуклеосомы и солевых суперструктур хроматина» . Журнал клеточной биологии . 83 (2 Части 1): 403–27. дои : 10.1083/jcb.83.2.403 . ПМК 2111545 . ПМИД 387806 .
- ^ Jump up to: а б ван Холде К., Златанова Дж (октябрь 1996 г.). «Что определяет складку хроматинового волокна?» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (20): 10548–55. Бибкод : 1996PNAS...9310548V . дои : 10.1073/pnas.93.20.10548 . ПМК 38190 . ПМИД 8855215 .
- ^ Рут А., Сандин С., Роудс Д. (июль 2008 г.). «Длина нуклеосомного повтора и стехиометрия линкерных гистонов определяют структуру волокон хроматина» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (26): 8872–7. Бибкод : 2008PNAS..105.8872R . дои : 10.1073/pnas.0802336105 . ПМЦ 2440727 . ПМИД 18583476 .
- ^ Варшавский А.Я., Бакаев В.В., Георгиев Г.П. (февраль 1976 г.). «Гетерогенность субъединиц хроматина in vitro и расположение гистона H1» . Исследования нуклеиновых кислот . 3 (2): 477–92. дои : 10.1093/нар/3.2.477 . ПМК 342917 . ПМИД 1257057 .
- ^ Уитлок Дж. П., Симпсон RT (июль 1976 г.). «Удаление гистона H1 обнажает сегмент ДНК из пятидесяти пар оснований между нуклеосомами» . Биохимия . 15 (15): 3307–14. дои : 10.1021/bi00660a022 . ПМИД 952859 .
- ^ Jump up to: а б Аллан Дж., Хартман П.Г., Крейн-Робинсон С., Авилес FX (декабрь 1980 г.). «Строение гистона Н1 и его расположение в хроматине» . Природа . 288 (5792): 675–9. Бибкод : 1980Natur.288..675A . дои : 10.1038/288675a0 . ПМИД 7453800 . S2CID 4262304 .
- ^ Стайнов Д.З., Крейн-Робинсон С. (декабрь 1988 г.). «Отпечаток линкерных гистонов H5 и H1 на нуклеосоме» . Журнал ЭМБО . 7 (12): 3685–91. дои : 10.1002/j.1460-2075.1988.tb03250.x . ПМК 454941 . ПМИД 3208745 .
- ^ Беднар Дж., Горовиц Р.А., Григорьев С.А., Каррутерс Л.М., Хансен Дж.К., Костер А.Дж., Вудкок К.Л. (ноябрь 1998 г.). «Нуклеосомы, линкерная ДНК и линкерный гистон образуют уникальный структурный мотив, который управляет сворачиванием и уплотнением хроматина высшего порядка» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (24): 14173–8. Бибкод : 1998PNAS...9514173B . дои : 10.1073/pnas.95.24.14173 . ПМК 24346 . ПМИД 9826673 .
- ^ Дворкин-Растл Э., Кандольф Х., Смит Р.К. (февраль 1994 г.). «Материнский вариант гистона H1, H1M (белок B4), является преобладающим гистоном H1 у эмбрионов Xenopus pregastrula». Биология развития . 161 (2): 425–39. дои : 10.1006/dbio.1994.1042 . ПМИД 8313993 .
- ^ Браун Д.Т., Александр Б.Т., Ситтман Д.Б. (февраль 1996 г.). «Дифференциальный эффект сверхэкспрессии варианта H1 на развитие клеточного цикла и экспрессию генов» . Исследования нуклеиновых кислот . 24 (3): 486–93. дои : 10.1093/нар/24.3.486 . ПМК 145659 . ПМИД 8602362 .
- ^ Гунджан А., Александр Б.Т., Ситтман Д.Б., Браун Д.Т. (декабрь 1999 г.). «Влияние сверхэкспрессии варианта гистона H1 на структуру хроматина» . Журнал биологической химии . 274 (53): 37950–6. дои : 10.1074/jbc.274.53.37950 . ПМИД 10608862 .
- ^ Мареска Т.Дж., Фридман Б.С., Хилд Р. (июнь 2005 г.). «Гистон H1 необходим для архитектуры и сегрегации митотических хромосом в экстрактах яиц Xenopus laevis» . Журнал клеточной биологии . 169 (6): 859–69. дои : 10.1083/jcb.200503031 . ПМК 2171634 . ПМИД 15967810 .
- ^ Jump up to: а б с Фридман Б.С., Хилд Р. (июнь 2010 г.). «Функциональное сравнение гистонов H1 у Xenopus выявило специфическую для изоформ регуляцию с помощью Cdk1 и RanGTP» . Современная биология . 20 (11): 1048–52. Бибкод : 2010CBio...20.1048F . дои : 10.1016/j.cub.2010.04.025 . ПМЦ 2902237 . ПМИД 20471264 .
- ^ Шен X, Ю Л, Вейр Дж. В., Горовский М. А. (июль 1995 г.). «Линкерные гистоны не являются существенными и влияют на конденсацию хроматина in vivo» . Клетка . 82 (1): 47–56. дои : 10.1016/0092-8674(95)90051-9 . ПМИД 7606784 . S2CID 14427681 .
- ^ Едрусик М.А., Шульце Э. (апрель 2001 г.). «Одна изоформа гистона H1 (H1.1) необходима для подавления хроматина и развития зародышевой линии у Caenorhabditis elegans» . Разработка . 128 (7): 1069–80. дои : 10.1242/dev.128.7.1069 . ПМИД 11245572 .
- ^ Лу Х, Вонтакал С.Н., Емельянов А.В., Морсильо П., Конев А.Ю., Федоров Д.В., Скулчи А.И. (февраль 2009 г.). «Линкерный гистон H1 необходим для развития дрозофилы, формирования перицентрического гетерохроматина и нормальной структуры политенной хромосомы» . Гены и развитие . 23 (4): 452–65. дои : 10.1101/gad.1749309 . ПМЦ 2648648 . ПМИД 19196654 .
- ^ Мистели Т., Гунджан А., Хок Р., Бастин М., Браун Д.Т. (декабрь 2000 г.). «Динамическое связывание гистона H1 с хроматином в живых клетках». Природа . 408 (6814): 877–81. Бибкод : 2000Natur.408..877M . дои : 10.1038/35048610 . ПМИД 11130729 . S2CID 4428934 .
- ^ Чен Д., Дундр М., Ван С., Люнг А., Ламонд А., Мистели Т., Хуанг С. (январь 2005 г.). «Конденсированный митотический хроматин доступен факторам транскрипции и структурным белкам хроматина» . Журнал клеточной биологии . 168 (1): 41–54. дои : 10.1083/jcb.200407182 . ПМК 2171683 . ПМИД 15623580 .
- ^ Бастин М., Катез Ф., Лим Дж. Х. (март 2005 г.). «Динамика функции гистона H1 в хроматине» . Молекулярная клетка . 17 (5): 617–20. doi : 10.1016/j.molcel.2005.02.019 . ПМИД 15749012 .
- ^ Сяо Б., Фридман Б.С., Миллер К.Е., Хилд Р., Марко Дж.Ф. (декабрь 2012 г.). «Гистон H1 уплотняет ДНК под действием силы и во время сборки хроматина» . Молекулярная биология клетки . 23 (24): 4864–71. doi : 10.1091/mbc.E12-07-0518 . ПМЦ 3521692 . ПМИД 23097493 .
- ^ Фридман Б.С., Миллер К.Е., Хилд Р. (сентябрь 2010 г.). Чимини Д. (ред.). «Экстракты яиц Xenopus повышают динамику гистона H1 на хроматине сперматозоидов» . ПЛОС ОДИН . 5 (9): e13111. Бибкод : 2010PLoSO...513111F . дои : 10.1371/journal.pone.0013111 . ПМЦ 2947519 . ПМИД 20927327 .
- ^ Штайнбах О.К., Вольф А.П., Рупп Р.А. (сентябрь 1997 г.). «Соматические линкерные гистоны вызывают потерю мезодермальной компетентности у Xenopus». Природа . 389 (6649): 395–9. Бибкод : 1997Natur.389..395S . дои : 10.1038/38755 . ПМИД 9311783 . S2CID 4390287 .
- ^ Де С., Браун Д.Т., Лу Ж.Х., Лено Г.Х., Веллман С.Е., Ситтман Д.Б. (июнь 2002 г.). «Варианты гистона H1 дифференциально ингибируют репликацию ДНК за счет сродства к хроматину, опосредованного их карбоксильно-концевыми доменами». Джин . 292 (1–2): 173–81. дои : 10.1016/S0378-1119(02)00675-3 . ПМИД 12119111 .
- ^ Хочбин С (июнь 2001 г.). «Разнообразие гистонов H1: соединение регуляторных сигналов с функцией линкерных гистонов» . Джин . 271 (1): 1–12. дои : 10.1016/S0378-1119(01)00495-4 . ПМИД 11410360 .
- ^ Годде Дж.С., Ура К. (март 2008 г.). «Взлом загадочного гистонового кода линкера» . Журнал биохимии . 143 (3): 287–93. дои : 10.1093/jb/mvn013 . ПМИД 18234717 .
- ^ Хаппель Н., Денеке Д. (февраль 2009 г.). «Гистон H1 и его изоформы: вклад в структуру и функцию хроматина» . Джин . 431 (1–2): 1–12. дои : 10.1016/j.gene.2008.11.003 . ПМИД 19059319 .
- ^ Харшман С.В., Янг Н.Л., Партун М.Р., Фрейтас М.А. (ноябрь 2013 г.). «Гистоны H1: современные перспективы и проблемы» . Исследования нуклеиновых кислот . 41 (21): 9593–609. дои : 10.1093/нар/gkt700 . ПМЦ 3834806 . ПМИД 23945933 .