Обработка речи в мозгу
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
В психолингвистике идей и чувств, а также к тому , языковая обработка относится к тому, как люди используют слова для передачи как такие сообщения обрабатываются и понимаются. Обработка речи считается уникальной человеческой способностью, которая не проявляется с таким же грамматическим человека- пониманием или систематичностью даже у ближайших родственников приматов . [1]
На протяжении всего ХХ века доминирующей моделью [2] Для обработки речи в мозге использовалась модель Гешвинда-Лихтейма-Вернике , которая основана, прежде всего, на анализе пациентов с повреждением головного мозга . Однако благодаря улучшениям внутрикорковых электрофизиологических записей мозга обезьян и человека , а также неинвазивным методам, таким как фМРТ , ПЭТ , МЭГ и ЭЭГ , двойной слуховой путь [3] [4] Выявлена и двухпотоковая модель разработана . В соответствии с этой моделью существует два пути, соединяющих слуховую кору с лобной долей , каждый из которых отвечает за разные лингвистические роли. Слуховой вентральный путь потока отвечает за распознавание звуков и поэтому известен как слуховой путь «что». Слуховой дорсальный поток как у людей, так и у приматов отвечает за локализацию звука и, соответственно, известен как слуховой путь «где». У людей этот путь (особенно в левом полушарии ) также отвечает за производство речи , повторение речи, чтение по губам , а также фонологическую рабочую и долговременную память . В соответствии с моделью эволюции языка «откуда к чему», [5] [6] Причина, по которой ADS характеризуется таким широким набором функций, заключается в том, что каждая из них указывает на разные стадии эволюции языка.
Разделение двух потоков сначала происходит в слуховом нерве , где передняя ветвь входит в переднее ядро улитки в стволе мозга, что дает начало слуховому вентральному потоку. Задняя ветвь входит в дорсальное и задне-вентральное ядро улитки, давая начало слуховому дорсальному потоку. [7] : 8
Языковая обработка может также происходить в отношении жестовых языков или письменного контента .
Ранние модели нейролингвистики
[ редактировать ]
На протяжении всего ХХ века в наших знаниях о языковой обработке мозга доминировала модель Вернике-Лихтхайма-Гешвинда. [8] [2] [9] Модель Вернике-Лихтхайма-Гешвинда в первую очередь основана на исследованиях, проведенных на людях с повреждением головного мозга, у которых, как сообщалось, были различные нарушения, связанные с речью. В соответствии с этой моделью слова воспринимаются через специализированный центр рецепции слов ( зону Вернике ), расположенный в левом височно-теменном переходе . Затем эта область проецируется на центр производства слов ( зону Брока ), который расположен в левой нижней лобной извилине . Поскольку считалось, что почти весь языковой ввод проходит через область Вернике, а весь языковой вывод — через область Брока, стало чрезвычайно сложно определить основные свойства каждой области. Отсутствие четкого определения вклада областей Вернике и Брока в человеческий язык чрезвычайно затрудняло идентификацию их гомологов у других приматов. [10] Однако с появлением фМРТ и ее применением для картирования поражений было показано, что эта модель основана на неправильных корреляциях между симптомами и поражениями. [11] [12] [13] [14] [15] [16] [17] Опровержение столь влиятельной и доминирующей модели открыло дверь новым моделям обработки речи в мозгу.
Современные модели нейролингвистики
[ редактировать ]Анатомия
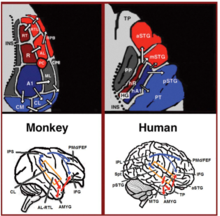
[ редактировать ]За последние два десятилетия произошел значительный прогресс в нашем понимании нейронной обработки звуков у приматов. Первоначально путем регистрации нейронной активности в слуховой коре обезьян. [18] [19] и позже уточнено с помощью гистологического окрашивания [20] [21] [22] и фМРТ , исследования с помощью [23] В первичной слуховой коре идентифицировано 3 слуховых поля, а вокруг них показано 9 ассоциативных слуховых полей (рис. 1 вверху слева). Анатомические исследования и исследования повреждений также показали разделение между передними и задними слуховыми полями, при этом передние первичные слуховые поля (области R-RT) проецируются на передние ассоциативные слуховые поля (области AL-RTL), а задние первичные слуховые поля (область А1), проецирующаяся на задние ассоциативные слуховые поля (области CL-CM). [20] [24] [25] [26] В последнее время накопились данные, указывающие на гомологию между слуховыми полями человека и обезьяны. У людей исследования гистологического окрашивания выявили два отдельных слуховых поля в первичной слуховой области извилины Хешля . [27] [28] и путем картирования тонотопической организации первичных слуховых полей человека с помощью фМРТ высокого разрешения и сравнения ее с тонотопической организацией первичных слуховых полей обезьяны была установлена гомология между передним первичным слуховым полем человека и областью R обезьяны (обозначенной у людей как область hR) и заднее первичное слуховое поле человека и область А1 обезьяны (обозначаемая у человека как область hA1). [29] [30] [31] [32] [33] Внутрикорковые записи слуховой коры человека также продемонстрировали сходные закономерности подключения со слуховой корой обезьяны. Запись с поверхности слуховой коры (надвисочная плоскость) показала, что передняя извилина Хешля (область hR) выступает преимущественно в средне-переднюю верхнюю височную извилину (mSTG-aSTG), а задняя извилина Хешля (область hA1) выступает в первую очередь. к задней верхней височной извилине (pSTG) и височной плоскости (область PT; рисунок 1 вверху справа). [34] [35] Связями между областью hR и aSTG и hA1 с pSTG согласуются данные фМРТ- исследования пациента с нарушением распознавания звуков ( слуховая агнозия ), у которого была выявлена сниженная двусторонняя активация в областях hR и aSTG, но с сохраненной активацией в mSTG. пСТГ. [36] Этот паттерн связи также подтверждается исследованием, в котором была зафиксирована активация с латеральной поверхности слуховой коры и сообщалось об одновременных неперекрывающихся кластерах активации в pSTG и mSTG-aSTG при прослушивании звуков. [37]
Ниже слуховой коры анатомические исследования на обезьянах выявили проекции от передних ассоциативных слуховых полей (областей AL-RTL) к вентральной префронтальной и премоторной коре нижней лобной извилины (IFG). [38] [39] и миндалевидное тело . [40] Исследования кортикальной записи и функциональной визуализации на макаках дополнительно детализировали этот поток обработки, показав, что акустическая информация течет от передней слуховой коры к височному полюсу (TP), а затем к IFG. [41] [42] [43] [44] [45] [46] Этот путь обычно называют слуховым вентральным потоком (AVS; рисунок 1, красные стрелки внизу слева). В отличие от передних слуховых полей, исследования показали, что задние слуховые поля (области CL-CM) проецируются в основном в дорсолатеральную префронтальную и премоторную кору (хотя некоторые проекции заканчиваются в IFG). [47] [39] Кортикальные записи и анатомические исследования на обезьянах также предоставили доказательства того, что этот поток обработки течет из задних слуховых полей в лобную долю через ретрансляционную станцию во внутритеменной борозде (IPS). [48] [49] [50] [51] [52] [53] Этот путь обычно называют слуховым дорсальным потоком (ADS; рисунок 1, синие стрелки внизу слева). Сравнение путей белого вещества, участвующих в коммуникации у людей и обезьян, с методами диффузионно-тензорной визуализации указывает на схожие связи AVS и ADS у двух видов (Monkey, [52] Человек [54] [55] [56] [57] [58] [59] ). Было показано, что у людей pSTG проецируется в теменную долю ( сильвиевское теменно-височное соединение - нижняя теменная долька ; Spt- IPL ), а оттуда в дорсолатеральную префронтальную и премоторную кору (рис. 1, синие стрелки внизу справа) и Было показано, что aSTG проецируется на переднюю височную долю (средняя височная извилина-височный полюс; MTG-TP) и оттуда на IFG (рис. 1, нижняя правая красная стрелка).
Слуховой вентральный поток
[ редактировать ]Слуховой вентральный поток (АВС) соединяет слуховую кору со средней височной извилиной и височным полюсом , который, в свою очередь, соединяется с нижней лобной извилиной . Этот путь отвечает за распознавание звуков и поэтому известен как слуховой путь «что». К функциям АВС относятся следующие.
Распознавание звука
[ редактировать ]Накопленные сходные данные указывают на то, что AVS участвует в распознавании слуховых объектов. На уровне первичной слуховой коры записи обезьян показали более высокий процент нейронов, избирательно реагирующих на выученные мелодические последовательности, в области R, чем в области A1. [60] а исследование на людях продемонстрировало большую избирательность к слышимым слогам в передней извилине Хешля (область hR), чем в задней извилине Хешля (область hA1). [61] В последующих ассоциативных слуховых полях исследования как на обезьянах, так и на людях показали, что граница между передними и задними слуховыми полями (рис. 1 — область PC у обезьяны и mSTG у человека) обрабатывает атрибуты высоты звука, необходимые для распознавания слуховых объектов. . [18] Передние слуховые поля обезьян также продемонстрировали избирательность в отношении специфических вокализаций с помощью внутрикорковых записей. [41] [19] [62] и функциональная визуализация [63] [42] [43] Одно исследование на обезьянах с помощью фМРТ дополнительно продемонстрировало роль аСТГ в распознавании отдельных голосов. [42] Роль человеческого mSTG-aSTG в распознавании звука была продемонстрирована с помощью исследований функциональной визуализации, которые коррелировали активность в этой области с изоляцией слуховых объектов от фонового шума. [64] [65] и с распознаванием произносимых слов, [66] [67] [68] [69] [70] [71] [72] голоса, [73] мелодии, [74] [75] звуки окружающей среды, [76] [77] [78] и неречевые коммуникативные звуки. [79] Метаанализ исследований фМРТ [80] далее продемонстрирована функциональная диссоциация между левым mSTG и aSTG: первый обрабатывает короткие речевые единицы (фонемы), а второй обрабатывает более длинные единицы (например, слова, звуки окружающей среды). Исследование, в котором регистрировалась нейронная активность непосредственно от левого pSTG и aSTG, показало, что aSTG, но не pSTG, был более активен, когда пациентка слушала речь на своем родном языке, чем незнакомый иностранный язык. [81] Соответственно, электростимуляция аСТГ у этого пациента приводила к нарушению восприятия речи. [81] (см. также [82] [83] для аналогичных результатов). Внутрикорковые записи правого и левого aSTG дополнительно продемонстрировали, что речь обрабатывается латерально по отношению к музыке. [81] При фМРТ- исследовании пациента с нарушением распознавания звуков ( слуховой агнозией ) вследствие поражения ствола мозга также было показано снижение активации в областях hR и aSTG обоих полушарий при прослушивании устной речи и звуков окружающей среды. [36] Записи передней слуховой коры обезьян при сохранении выученных звуков в рабочей памяти, [46] и изнурительный эффект индуцированных повреждений этой области на воспроизведение рабочей памяти, [84] [85] [86] еще больше вовлекают AVS в поддержание воспринимаемых слуховых объектов в рабочей памяти. Сообщалось также, что у людей область mSTG-aSTG активна во время повторения услышанных слогов с помощью MEG. [87] и фМРТ [88] Последнее исследование дополнительно продемонстрировало, что рабочая память в AVS отвечает за акустические свойства произносимых слов и что она не зависит от рабочей памяти в ADS, которая опосредует внутреннюю речь. Исследования рабочей памяти на обезьянах также показывают, что у обезьян, в отличие от людей, AVS является доминирующим хранилищем рабочей памяти. [89]
Считается, что у людей, расположенных ниже aSTG, MTG и TP составляют семантический словарь , который представляет собой хранилище долговременной памяти аудиовизуальных представлений, связанных между собой на основе семантических отношений. (См. также обзоры [3] [4] обсуждаем эту тему). Основным доказательством этой роли MTG-TP является то, что о пациентах с повреждением этой области (например, о пациентах с семантической деменцией или энцефалитом, вызванным вирусом простого герпеса ). сообщается [90] [91] с нарушением способности описывать зрительные и слуховые объекты и склонностью к совершению смысловых ошибок при назывании предметов (т. е. семантическая парафазия ). Семантические парафазии были выражены и у пациентов с афазией с повреждением левого МТГ-ТП. [14] [92] и было показано, что они возникают у неафазических пациентов после электростимуляции этой области. [93] [83] или основной путь белого вещества [94] Два метаанализа литературы по фМРТ также показали, что передние MTG и TP были последовательно активны во время семантического анализа речи и текста; [66] [95] а исследование внутрикортикальной записи коррелировало нейронные разряды в MTG с пониманием понятных предложений. [96]
Понимание предложений
[ редактировать ]Помимо извлечения значения из звуков, область MTG-TP AVS, по-видимому, играет роль в понимании предложений, возможно, путем слияния понятий (например, слияния понятий «синий» и «рубашка» для создания понятия «синий» и «рубашка» для создания понятия «синий» и «рубашка»). синяя рубашка»). Роль MTG в извлечении смысла из предложений была продемонстрирована в исследованиях функциональной визуализации, в которых сообщалось о более сильной активации передней MTG, когда правильные предложения противопоставляются спискам слов, предложениям на иностранном или бессмысленном языке, зашифрованным предложениям, предложениям со семантикой или синтаксисом. нарушения и предложения-подобные последовательности звуков окружающей среды. [97] [98] [99] [100] [101] [102] [103] [104] Одно фМРТ- исследование [105] в котором участникам было поручено прочитать рассказ, активность передней MTG дополнительно коррелировала с объемом семантического и синтаксического содержания, содержащегося в каждом предложении. ЭЭГ-исследование [106] сравнив корковую активность при чтении предложений с синтаксическими нарушениями и без них у здоровых участников и пациентов с повреждением MTG-TP, пришли к выводу, что MTG-TP в обоих полушариях участвуют в автоматическом (основанном на правилах) этапе синтаксического анализа (компонент ELAN) и что левый MTG-TP также участвует в более позднем контролируемом этапе синтаксического анализа (компонент P600). Также сообщалось о нарушениях понимания предложений у пациентов с повреждением области MTG-TP. [14] [107] [108] Посмотреть обзор [109] для получения дополнительной информации по этой теме.
Двусторонность
[ редактировать ]В отличие от модели Вернике-Лиххейма-Гешвинда, которая предполагает, что распознавание звуков происходит исключительно в левом полушарии, исследования, в которых изучались свойства правого или левого полушария изолированно посредством односторонней полушарной анестезии (т. е. процедуры ВАДА) [110] ) или внутрикорковые записи каждого полушария [96] предоставил доказательства того, что распознавание звука осуществляется на двусторонней основе. Более того, исследование, в котором пациентам с отключенными полушариями (т. е. пациентам с расщепленным мозгом ) предлагалось сопоставить произнесенные слова с письменными словами, представленными в правом или левом полушарии, показало, что словарный запас в правом полушарии почти совпадает по размеру с левым полушарием. [111] (Словарь правого полушария был эквивалентен словарю здорового 11-летнего ребенка). Такое двустороннее распознавание звуков также согласуется с выводом о том, что одностороннее поражение слуховой коры редко приводит к нарушению слухового восприятия (т. е. к слуховой агнозии ), тогда как второе поражение оставшегося полушария (которое может произойти спустя годы) приводит к этому. [112] [113] Наконец, как упоминалось ранее, фМРТ пациента со слуховой агнозией продемонстрировало двустороннее снижение активации передней слуховой коры. [36] и двусторонняя электростимуляция этих областей в обоих полушариях приводила к нарушению распознавания речи. [81]
Слуховой спинной поток
[ редактировать ]Слуховой дорсальный поток соединяет слуховую кору с теменной долей , которая, в свою очередь, соединяется с нижней лобной извилиной . И у людей, и у приматов слуховой дорсальный поток отвечает за локализацию звука и, соответственно, известен как слуховой путь «где». У людей этот путь (особенно в левом полушарии) также отвечает за производство речи, повторение речи, чтение по губам, а также фонологическую рабочую и долговременную память.
Речевое производство
[ редактировать ]Исследования современных людей продемонстрировали роль ADS в производстве речи, особенно в голосовом выражении названий объектов. Например, в серии исследований, в которых подкорковые волокна непосредственно стимулировались [94] вмешательство в левый pSTG и IPL приводило к ошибкам при выполнении задач по именованию объектов, а вмешательство в левый IFG приводило к остановке речи. Магнитные помехи в pSTG и IFG здоровых участников также вызывали речевые ошибки и остановку речи соответственно. [114] [115] В одном исследовании также сообщалось, что электрическая стимуляция левой IPL заставляла пациентов верить, что они говорили, хотя на самом деле это не так, и что стимуляция IFG заставляла пациентов бессознательно двигать губами. [116] Вклад ADS в процесс артикуляции названий объектов может зависеть от приема афферентов из семантического лексикона AVS, поскольку исследование внутрикорковой записи показало активацию в заднем MTG до активации в Spt. - Область IPL , когда пациенты называли объекты на картинках [117] Исследования внутрикорковой электростимуляции также показали, что электрические помехи в задней части MTG коррелируют с нарушением называния объектов. [118] [82]
Вокальная мимикрия
[ редактировать ]Хотя восприятие звука в первую очередь связано с AVS, ADS, по-видимому, связан с несколькими аспектами восприятия речи. Например, в метаанализе фМРТ исследований [119] (Turkeltaub, Coslett, 2010), в которых слуховое восприятие фонем противопоставлялось близко совпадающим звукам, а исследования оценивались по необходимому уровню внимания, авторы пришли к выводу, что внимание к фонемам коррелирует с сильной активацией в pSTG-pSTS. область. Исследование внутрикорковой записи, в котором участников просили идентифицировать слоги, также коррелировало слух каждого слога с его собственным паттерном активации в pSTG. [120] Участие ADS как в восприятии, так и в производстве речи было дополнительно освещено в нескольких новаторских исследованиях функциональной визуализации, в которых сравнивалось восприятие речи с явным или скрытым речевым производством. [121] [122] [123] Эти исследования показали, что pSTS активен только во время восприятия речи, тогда как область Spt активна как во время восприятия, так и во время производства речи. Авторы пришли к выводу, что pSTS проецируется в область Spt, которая преобразует слуховой сигнал в артикуляционные движения. [124] [125] Аналогичные результаты были получены в исследовании, в котором височные и теменные доли участников подвергались электрической стимуляции. В этом исследовании сообщалось, что электрическая стимуляция области pSTG мешает пониманию предложений, а стимуляция IPL мешает способности озвучивать названия объектов. [83] Авторы также сообщили, что стимуляция в области Spt и нижней IPL вызывает помехи как во время задач по названию объектов, так и при понимании речи. Роль ADS в повторении речи также согласуется с результатами других исследований функциональной визуализации, которые локализовали активацию во время задач повторения речи в областях ADS. [126] [127] [128] Исследование внутрикорковой записи, в ходе которого регистрировалась активность в большинстве височных, теменных и лобных долей, также показало активацию pSTG, Spt, IPL и IFG, когда повторение речи контрастирует с восприятием речи. [129] Нейропсихологические исследования также показали, что люди с дефицитом повторения речи, но с сохраненным слуховым пониманием (т. е. с афазией проводимости ) страдают от ограниченного повреждения области Spt-IPL. [130] [131] [132] [133] [134] [135] [136] или повреждение проекций, исходящих из этой области и нацеленных на лобную долю [137] [138] [139] [140] Исследования также сообщили о временном дефиците повторения речи у пациентов после прямой внутрикорковой электростимуляции той же области. [11] [141] [142] Понимание цели повторения речи в ADS дает лонгитудинальное исследование детей, в ходе которого изучение иностранной лексики коррелировало со способностью повторять бессмысленные слова. [143] [144]
Речевой мониторинг
[ редактировать ]Помимо повторения и воспроизведения речи, ADS, по-видимому, играет роль в контроле качества речевого вывода. Нейроанатомические данные свидетельствуют о том, что ADS оснащен нисходящими связями от IFG к pSTG, которые передают информацию о двигательной активности (т. е. вторичных разрядах) в голосовом аппарате (рот, язык, голосовые связки). Эта обратная связь отмечает звук, воспринимаемый во время произнесения речи, как произведённый самим человеком, и может использоваться для настройки голосового аппарата для увеличения сходства между воспринимаемыми и издаваемыми звуками. Доказательства нисходящих связей от IFG к pSTG были предложены в исследовании, в котором электрическая стимуляция IFG во время хирургических операций выявила распространение активации на область pSTG-pSTS-Spt. [145] Исследование [146] которые сравнивали способность пациентов с афазией с повреждением лобной, теменной или височной доли быстро и многократно артикулировать цепочку слогов, сообщили, что повреждение лобной доли мешает артикуляции как идентичных слоговых цепочек («бабаба»), так и неидентичных слоговых цепочек. струны («Бадага»), тогда как у пациентов с повреждением височной или теменной доли нарушения наблюдались только при артикуляции неидентичных слоговых строк. Поскольку пациенты с повреждением височных и теменных долей были способны повторять слоговую строку в первом задании, их речевое восприятие и производство, по-видимому, относительно сохранены, и их дефицит во втором задании, следовательно, обусловлен нарушением контроля. Демонстрируя роль нисходящих соединений ADS в мониторинге исходящих вызовов, исследование фМРТ давало участникам указание говорить в нормальных условиях или при прослушивании измененной версии своего собственного голоса (задержка первой форманты) и сообщало, что прослушивание искаженной версии собственного голоса приводит к в повышенной активации пСТГ. [147] Еще одной демонстрацией того, что ADS облегчает двигательную обратную связь во время мимики, является исследование внутрикорковой записи, в котором сравнивались восприятие речи и повторение. [129] Авторы сообщили, что, помимо активации в IPL и IFG, повторение речи характеризуется более сильной активацией в pSTG, чем при восприятии речи.
Интеграция фонем с движениями губ.
[ редактировать ]Хотя восприятие звука в первую очередь связано с AVS, ADS, по-видимому, связан с несколькими аспектами восприятия речи. Например, в метаанализе фМРТ исследований [119] В которых слуховому восприятию фонем противопоставлялись близко совпадающие звуки, а исследования оценивались по необходимому уровню внимания, авторы пришли к выводу, что внимание к фонемам коррелирует с сильной активацией в области pSTG-pSTS. Исследование внутрикорковой записи, в котором участников просили идентифицировать слоги, также коррелировало слух каждого слога с его собственным паттерном активации в pSTG. [148] В соответствии с ролью ADS в различении фонем, [119] исследования приписали интеграцию фонем и соответствующих им движений губ (т. е. визем) pSTS ADS. Например, фМРТ- исследование [149] коррелировал активацию pSTS с иллюзией Макгерка (при которой слушание слога «ба» при виде виземы «га» приводит к восприятию слога «да»). Другое исследование показало, что использование магнитной стимуляции для вмешательства в обработку данных в этой области еще больше разрушает иллюзию Макгерка. [150] Связь pSTS с аудиовизуальной интеграцией речи также была продемонстрирована в исследовании, в котором участникам предлагались изображения лиц и произнесенных слов различного качества. В исследовании сообщается, что PSTS обеспечивает комбинированное увеличение четкости лиц и произнесенных слов. [151] Подтверждающие доказательства были предоставлены фМРТ. исследованием [152] которые противопоставляли восприятие аудиовизуальной речи аудиовизуальному неречи (изображениям и звукам инструментов). В этом исследовании сообщалось об обнаружении речевых селективных компартментов в pSTS. Кроме того, фМРТ- исследование [153] которые сравнивали конгруэнтную аудиовизуальную речь с неконгруэнтной речью (изображениями неподвижных лиц) сообщили об активации pSTS. Обзор, представляющий дополнительные сходящиеся данные относительно роли pSTS и ADS в интеграции фонем и визем, см. [154]
Фонологическая долговременная память
[ редактировать ]Растущее количество данных указывает на то, что люди, помимо наличия долговременного хранилища значений слов, расположенного в MTG-TP AVS (т. е. семантического лексикона), также имеют долгосрочное хранилище для названий объектов. расположен в области Spt-IPL ADS (т. е. фонологическом лексиконе). Например, исследование [155] [156] Обследование пациентов с повреждением AVS (повреждение MTG) или повреждением ADS (повреждение IPL) показало, что повреждение MTG приводит к тому, что люди неправильно идентифицируют объекты (например, называют «козу» «овцой», пример семантической парафазии ). И наоборот, повреждение IPL приводит к тому, что люди правильно идентифицируют объект, но неправильно произносят его имя (например, говорят «гоф» вместо «коза», пример фонематической парафазии ). Ошибки семантической парафазии также были зарегистрированы у пациентов, получавших внутрикорковую электрическую стимуляцию AVS (MTG), а ошибки фонематической парафазии были зарегистрированы у пациентов, чьи ADS (pSTG, Spt и IPL) получали внутрикорковую электрическую стимуляцию. [83] [157] [94] Дополнительным подтверждением роли ADS в именовании объектов является исследование MEG, в котором локализовалась активность IPL во время обучения и во время вспоминания имен объектов. [158] Исследование, в котором вызывалось магнитное вмешательство в IPL участников, когда они отвечали на вопросы об объекте, показало, что участники были способны отвечать на вопросы, касающиеся характеристик объекта или атрибутов восприятия, но у них были проблемы, когда их спрашивали, состоит ли слово из двух или трех слогов. [159] Исследование MEG также коррелирует выздоровление от аномии (расстройства, характеризующегося нарушением способности называть объекты) с изменениями активации IPL. [160] Еще одним подтверждением роли IPL в кодировании звуков слов являются исследования, в которых сообщается, что по сравнению с монолингвами билингвы имеют большую корковую плотность в IPL, но не в MTG. [161] [162] Поскольку данные показывают, что у билингвов разные фонологические представления одного и того же слова имеют одно и то же семантическое представление, [163] такое увеличение плотности IPL подтверждает существование фонологической лексики: ожидается, что семантическая лексика билингвов будет аналогична по размеру семантической лексике монолингвов, тогда как их фонологическая лексика должна быть в два раза больше. В соответствии с этим выводом, плотность коры IPL у монолингвов также коррелирует с размером словарного запаса. [164] [165] Примечательно, что функциональная диссоциация AVS и ADS в задачах по именованию объектов подтверждается совокупными данными исследований чтения, показывающих, что семантические ошибки коррелируют с нарушениями MTG, а фонематические ошибки - с нарушениями IPL. На основании этих ассоциаций семантический анализ текста был связан с нижневисочной извилиной и MTG, а фонологический анализ текста — с pSTG-Spt-IPL. [166] [167] [168]
Фонологическая рабочая память
[ редактировать ]Рабочую память часто трактуют как временную активацию хранящихся в долговременной памяти представлений, которые используются для речи (фонологические представления). Это совместное использование ресурсов между рабочей памятью и речью очевидно из открытия [169] [170] что разговор во время репетиции приводит к значительному уменьшению количества элементов, которые можно вспомнить из рабочей памяти ( артикуляционное подавление ). Об участии фонологической лексики в рабочей памяти свидетельствует также склонность лиц совершать больше ошибок при вспоминании слов из недавно выученного списка фонологически схожих слов, чем из списка фонологически несходных слов ( эффект фонологического сходства ). [169] Исследования также показали, что речевые ошибки, совершаемые при чтении, удивительно похожи на речевые ошибки, допущенные при воспроизведении недавно выученных фонологически похожих слов из рабочей памяти. [171] У пациентов с повреждением IPL также наблюдались как ошибки произношения речи, так и нарушения рабочей памяти. [172] [173] [174] [175] Наконец, точка зрения о том, что вербальная рабочая память является результатом временной активации фонологических представлений в ADS, совместима с недавними моделями, описывающими рабочую память как комбинацию сохранения представлений в механизме внимания параллельно с временной активацией представлений в долговременной памяти. [170] [176] [177] [178] Утверждалось, что роль ADS в повторении списков слов является причиной того, что этот путь активен во время понимания предложения. [179] Обзор роли АДС в рабочей памяти см. [180]

Лингвистические теории
[ редактировать ]Исследования в области языковой обработки служат основой для теорий языка . Главный теоретический вопрос заключается в том, вытекают ли лингвистические структуры из структур мозга или наоборот. Экстерналистские модели, такие как Фердинанда де Соссюра структурализм , утверждают, что язык как социальное явление является внешним по отношению к мозгу. Языковую систему человек получает извне, а данный язык формирует мозг человека. [181]
Этой идее противостоят интерналистские модели, в том числе Ноама Хомского трансформационная порождающая грамматика , Джорджа Лакоффа когнитивная лингвистика и гипотеза эффективности Джона А. Хокинса . По мнению Хомского, язык приобретается за счет врожденной структуры мозга независимо от значения. [182] Лакофф утверждает, что язык возникает из сенсорных систем . [183] Хокинс выдвигает гипотезу, что преобладающие в разных языках паттерны основаны на естественных предпочтениях мозга в обработке информации. [184]
Кроме того, модели, вдохновленные Ричарда Докинза меметикой , в том числе строительная грамматика и лингвистика, основанная на использовании , защищают двустороннюю модель, утверждая, что мозг формирует язык, а язык формирует мозг. [185] [186]
Данные нейровизуализационных исследований указывают на экстерналистскую позицию. Исследования ERP показывают, что языковая обработка основана на взаимодействии синтаксиса и семантики, и исследования не поддерживают врожденные грамматические структуры. [187] [188] Исследования МРТ ребенка показывают, что структурные характеристики первого языка формируют обрабатывающий коннектом мозга. [189] Исследования обработки не смогли найти подтверждения обратной идее о том, что синтаксические структуры отражают естественные предпочтения мозга в межлингвистической обработке. [190]
Эволюция языка
[ редактировать ]Слуховой дорсальный поток также выполняет функции, не связанные с языком, такие как локализация звука. [191] [192] [193] [194] [195] и управление движениями глаз. [196] [197] Недавние исследования также указывают на роль ADS в локализации членов семьи/племени. [198] Исследование, записанное в коре головного мозга пациента с эпилепсией, показало, что pSTG, но не aSTG, избирательно реагирует на присутствие новых говорящих. фМРТ [199] исследование плодов в третьем триместре также показало, что область Spt более избирательна к женской речи, чем чистые тона, а часть Spt избирательна к речи матери в отличие от незнакомых женских голосов.
В настоящее время неизвестно, почему АДС человека приписывают так много функций. Попытка объединить эти функции в рамках единой структуры была предпринята в модели эволюции языка «Откуда к чему». [200] [201] В соответствии с этой моделью каждая функция АДС указывает на различную промежуточную фазу эволюции языка. Роль локализации звука и интеграция местоположения звука с голосами и слуховыми объектами интерпретируется как свидетельство того, что происхождение речи - это обмен контактными звонками (сигналами, используемыми для сообщения о местоположении в случаях разлуки) между матерями и потомством. Роль АДС в восприятии и производстве интонаций интерпретируется как свидетельство того, что речь началась с модификации контактных звонков интонациями, возможно, для различения тревожных контактных звонков от безопасных контактных звонков. Роль АДС в кодировании названий предметов (долговременная фонологическая память) интерпретируется как свидетельство постепенного перехода от модификации призывов интонациями к полному речевому контролю. Роль ADS в интеграции движений губ с фонемами и в повторении речи интерпретируется как свидетельство того, что произнесенные слова были усвоены младенцами, имитирующими вокализации своих родителей, первоначально путем имитации движений их губ. Роль ADS в фонологической рабочей памяти интерпретируется как свидетельство того, что слова, выученные посредством мимики, оставались активными в ADS, даже когда они не произносились. В результате появились люди, способные репетировать список вокализаций, что позволило произносить слова из нескольких слогов. Дальнейшие разработки в ADS позволили воспроизводить списки слов, что обеспечило инфраструктуру для связи с предложениями.
Язык жестов в мозгу
[ редактировать ]Нейробиологические исследования обеспечили научное понимание того, как язык жестов обрабатывается в мозгу . В мире существует более 135 отдельных жестовых языков, в которых используются разные акценты, характерные для отдельных регионов страны. [202]
Прибегнув к анализу поражений и нейровизуализации, нейробиологи обнаружили, что будь то разговорный язык или язык жестов, человеческий мозг обрабатывает язык в целом аналогичным образом в зависимости от того, какая область мозга используется. [202] Анализ повреждений используется для изучения последствий повреждения определенных областей мозга, участвующих в речевой деятельности, в то время как нейровизуализация исследует области, участвующие в обработке речи. [202]
Ранее выдвигались гипотезы о том, что повреждение зоны Брока или зоны Вернике не влияет на восприятие языка жестов; однако это не так. Исследования показали, что повреждение этих областей приводит к одинаковым результатам в разговорной речи, где присутствуют и/или повторяются жестовые ошибки. [202] В обоих типах языков поражается левое полушарие мозга, а не правое, обычно связанное с искусством.
Существуют очевидные закономерности использования и обработки языка. В языке жестов активируется зона Брока, в то время как при обработке языка жестов используется зона Вернике, аналогичная той, что используется в разговорной речи. [202]
Были и другие гипотезы о латерализации двух полушарий. В частности, считалось, что правое полушарие способствует общему общению на языке в глобальном масштабе, тогда как левое полушарие будет доминировать в создании языка на местном уровне. [203] В ходе исследований афазий у лиц, подписывающих знаки RHD, было обнаружено, что они испытывают проблемы с сохранением пространственной части своих знаков, из-за чего они путают одинаковые знаки в разных местах, необходимые для правильного общения с другим человеком. [203] С другой стороны, у лиц, подписавших ЛХД, результаты были аналогичны результатам, полученным у слышащих пациентов. Более того, другие исследования подчеркнули, что язык жестов присутствует на двусторонней основе, но необходимо продолжить исследования, чтобы прийти к выводу. [203]
Пишу в мозгу
[ редактировать ]Существует сравнительно небольшой объем исследований по неврологии чтения и письма. [204] Большинство проведенных исследований посвящено чтению, а не письму или правописанию, и большинство исследований обоих видов сосредоточено исключительно на английском языке. [205] Английская орфография менее прозрачна, чем орфография других языков, использующих латиницу . [204] Другая трудность заключается в том, что некоторые исследования сосредоточены на написании английских слов и опускают несколько логографических символов, встречающихся в письме. [204]
С точки зрения написания английские слова можно разделить на три категории: обычные, неправильные и «новые слова» или «неслова». существует регулярное, однозначное соответствие между графемой и фонемой Правильные слова — это те, в которых в написании . Неправильные слова – это слова, в которых такого соответствия нет. Неслова — это те, которые демонстрируют ожидаемую орфографию обычных слов, но не несут смысла, например слова-нонсы и звукоподражания . [204]
Проблема когнитивного и неврологического исследования чтения и правописания на английском языке заключается в том, какая модель лучше всего описывает, как грамотные носители могут читать и писать все три категории английских слов в соответствии с принятыми стандартами орфографической правильности. Модели с одним маршрутом предполагают, что лексическая память используется для хранения всех вариантов написания слов для поиска в одном процессе. Модели двойного маршрута предполагают, что лексическая память используется для обработки нерегулярных и высокочастотных регулярных слов, в то время как низкочастотные регулярные слова и неслова обрабатываются с использованием сублексического набора фонологических правил. [204]
Модель однонаправленного чтения нашла поддержку в исследованиях компьютерного моделирования, которые предполагают, что читатели идентифицируют слова по их орфографическому сходству с фонологически похожими словами. [204] Однако исследования когнитивных функций и поражений склоняются к модели двойного пути. Исследования когнитивного правописания на детях и взрослых показывают, что орфографы используют фонологические правила при написании обычных слов и неслов, в то время как лексическая память используется для написания неправильных слов и высокочастотных слов всех типов. [204] Аналогичным образом, исследования повреждений показывают, что лексическая память используется для хранения неправильных слов и некоторых обычных слов, в то время как фонологические правила используются для написания неслов. [204]
Совсем недавно исследования нейровизуализации с использованием позитронно-эмиссионной томографии и фМРТ предложили сбалансированную модель, в которой чтение всех типов слов начинается в области визуальной словоформы , но впоследствии разветвляется на разные маршруты в зависимости от того, имеется ли доступ к лексической памяти или семантической памяти. необходима информация (чего можно было бы ожидать при наличии неправильных слов в модели с двойным маршрутом). [204] 2007 года Исследование фМРТ показало, что испытуемые, которых просили произносить правильные слова в орфографическом задании, демонстрировали большую активацию левой задней STG , области, используемой для фонологической обработки, в то время как написание неправильных слов вызывало большую активацию областей, используемых для лексической памяти и семантической обработки. , такие как левый IFG и левый SMG и оба полушария MTG . [204] Было обнаружено, что правописание неслов дает доступ к членам обоих путей, таким как левый STG и двусторонний MTG и ITG . [204] Примечательно, что было обнаружено, что правописание вызывает активацию в таких областях, как левая веретенообразная извилина и левый SMG, которые также важны при чтении, что позволяет предположить, что аналогичный путь используется как для чтения, так и для письма. [204]
Гораздо меньше информации существует о познании и неврологии неалфавитных и неанглийских сценариев. Каждый язык имеет морфологический и фонологический компоненты, каждый из которых может быть записан системой письма . Сценарии, записывающие слова и морфемы, считаются логографическими , а сценарии, записывающие фонологические сегменты, такие как слоговые письма и алфавиты , — фонографическими. [205] Большинство систем сочетают в себе эти две системы и имеют как логографические, так и фонографические символы. [205]
По сложности системы письма можно охарактеризовать как «прозрачные» или «непрозрачные», а также как «поверхностные» или «глубокие». В «прозрачной» системе наблюдается очевидное соответствие между графемой и звуком, тогда как в «непрозрачной» системе эта связь менее очевидна. Термины «поверхностный» и «глубокий» относятся к степени, в которой орфография системы представляет морфемы, а не фонологические сегменты. [205] Системы, которые записывают более крупные морфосинтаксические или фонологические сегменты, такие как логографические системы и слоговые письма, предъявляют большие требования к памяти пользователей. [205] Таким образом, можно было бы ожидать, что непрозрачная или глубокая система письма потребует большего внимания к областям мозга, используемым для лексической памяти, чем система с прозрачной или поверхностной орфографией.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Зайденберг М.С., Петитто Л.А. (1987). «Коммуникация, символическая коммуникация и язык: комментарий к Сэвиджу-Рамбо, Макдональду, Севчику, Хопкинсу и Руперту (1986)». Журнал экспериментальной психологии: Общие сведения . 116 (3): 279–287. дои : 10.1037/0096-3445.116.3.279 . S2CID 18329599 .
- ^ Перейти обратно: а б Гешвинд Н. (июнь 1965 г.). «Синдромы отключения у животных и человека. I» . обзор. Мозг . 88 (2): 237–94. дои : 10.1093/мозг/88.2.237 . ПМИД 5318481 .
- ^ Перейти обратно: а б Хикок Г., Поппель Д. (май 2007 г.). «Корковая организация обработки речи». обзор. Обзоры природы. Нейронаука . 8 (5): 393–402. дои : 10.1038/nrn2113 . ПМИД 17431404 . S2CID 6199399 .
- ^ Перейти обратно: а б Гоу Д.В. (июнь 2012 г.). «Корковая организация лексических знаний: двойственная лексическая модель обработки разговорной речи» . обзор. Мозг и язык . 121 (3): 273–88. дои : 10.1016/j.bandl.2012.03.005 . ПМЦ 3348354 . ПМИД 22498237 .
- ^ Полива О (20 сентября 2017 г.). «Откуда к чему: нейроанатомически обоснованная эволюционная модель возникновения речи у человека» . обзор. F1000Исследования . 4 : 67. doi : 10.12688/f1000research.6175.3 . ПМК 5600004 . ПМИД 28928931 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Полива О (2016). «От мимикрии к языку: нейроанатомическая эволюционная модель возникновения голосового языка» . обзор. Границы в неврологии . 10 : 307. дои : 10.3389/fnins.2016.00307 . ПМЦ 4928493 . ПМИД 27445676 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Пиклз Дж.О. (2015). «Глава 1: Слуховые пути: анатомия и физиология». В Аминофф М.Дж., Боллер Ф., Свааб Д.Ф. (ред.). Справочник по клинической неврологии . обзор. Том. 129. стр. 3–25. дои : 10.1016/B978-0-444-62630-1.00001-9 . ISBN 978-0-444-62630-1 . ПМИД 25726260 .
- ^ Лихтейм Л. (1 января 1885 г.). «Об афазии». Мозг . 7 (4): 433–484. дои : 10.1093/мозг/7.4.433 . hdl : 11858/00-001M-0000-002C-5780-B .
- ^ Вернике С (1974). Афазический симптомокомплекс . Шпрингер Берлин Гейдельберг. стр. 1–70. ISBN 978-3-540-06905-8 .
- ^ Абоитис Ф., Гарсиа В.Р. (декабрь 1997 г.). «Эволюционное происхождение языковых областей в человеческом мозге. Нейроанатомический взгляд». Исследования мозга. Обзоры исследований мозга . 25 (3): 381–96. дои : 10.1016/s0165-0173(97)00053-2 . ПМИД 9495565 . S2CID 20704891 .
- ^ Перейти обратно: а б Андерсон Дж.М., Гилмор Р., Ропер С., Кроссон Б., Бауэр Р.М., Надо С., Беверсдорф Д.К., Сибула Дж., Рогиш М., Кортенкамп С., Хьюз Дж.Д., Гонсалес Роти Л.Дж., Хейлман К.М. (октябрь 1999 г.). «Проводниковая афазия и дугообразный пучок: пересмотр модели Вернике-Гешвинда». Мозг и язык . 70 (1): 1–12. дои : 10.1006/brln.1999.2135 . ПМИД 10534369 . S2CID 12171982 .
- ^ ДеВитт И., Раушекер Дж.П. (ноябрь 2013 г.). «Возвращение к области Вернике: параллельные потоки и обработка текста» . Мозг и язык . 127 (2): 181–91. дои : 10.1016/j.bandl.2013.09.014 . ПМК 4098851 . ПМИД 24404576 .
- ^ Дронкерс НФ (январь 2000 г.). «Стремление к связям мозга и языка». Мозг и язык . 71 (1): 59–61. дои : 10.1006/brln.1999.2212 . ПМИД 10716807 . S2CID 7224731 .
- ^ Перейти обратно: а б с Дронкерс Н.Ф., Уилкинс Д.П., Ван Валин Р.Д., Редферн Б.Б., Джагер Дж.Дж. (май 2004 г.). «Анализ повреждений областей мозга, участвующих в понимании языка». Познание . 92 (1–2): 145–77. дои : 10.1016/j.cognition.2003.11.002 . hdl : 11858/00-001M-0000-0012-6912-A . ПМИД 15037129 . S2CID 10919645 .
- ^ Месулам М.М., Томпсон К.К., Вайнтрауб С., Рогальски Э.Дж. (август 2015 г.). «Загадка Вернике и анатомия понимания речи при первичной прогрессирующей афазии» . Мозг . 138 (Часть 8): 2423–37. дои : 10.1093/brain/awv154 . ПМК 4805066 . ПМИД 26112340 .
- ^ Поппель Д. , Эммори К. , Хикок Г., Пюлкканен Л. (октябрь 2012 г.). «На пути к новой нейробиологии языка» . Журнал неврологии . 32 (41): 14125–31. doi : 10.1523/jneurosci.3244-12.2012 . ПМК 3495005 . ПМИД 23055482 .
- ^ Виньоло Л.А., Боккарди Э., Каверни Л. (март 1986 г.). «Неожиданные результаты компьютерной томографии при глобальной афазии» . Кора; Журнал, посвященный изучению нервной системы и поведения . 22 (1): 55–69. дои : 10.1016/s0010-9452(86)80032-6 . ПМИД 2423296 . S2CID 4479679 .
- ^ Перейти обратно: а б Бендор Д., Ван X (август 2006 г.). «Корковые представления слуха у обезьян и людей» . Современное мнение в нейробиологии . 16 (4): 391–9. дои : 10.1016/j.conb.2006.07.001 . ПМЦ 4325365 . ПМИД 16842992 .
- ^ Перейти обратно: а б Раушекер Дж. П., Тиан Б., Хаузер М. (апрель 1995 г.). «Обработка сложных звуков в непервичной слуховой коре макака». Наука . 268 (5207): 111–4. Бибкод : 1995Sci...268..111R . дои : 10.1126/science.7701330 . ПМИД 7701330 . S2CID 19590708 .
- ^ Перейти обратно: а б де ла Мот Л.А., Блюмелл С., Кадзикава Ю., Хакетт Т.А. (май 2006 г.). «Корковые связи слуховой коры мартышек: области ядра и медиального пояса». Журнал сравнительной неврологии . 496 (1): 27–71. дои : 10.1002/cne.20923 . ПМИД 16528722 . S2CID 38393074 .
- ^ де ла Мот Л.А., Блюмелл С., Кадзикава Ю., Хакетт Т.А. (май 2012 г.). «Корковые связи слуховой коры мартышек: латеральный пояс и парапоясной отдел» . Анатомическая запись . 295 (5): 800–21. дои : 10.1002/ar.22451 . ПМЦ 3379817 . ПМИД 22461313 .
- ^ Каас Дж. Х., Хакетт Т. А. (октябрь 2000 г.). «Подразделения слуховой коры и потоки обработки информации у приматов» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (22): 11793–9. Бибкод : 2000PNAS...9711793K . дои : 10.1073/pnas.97.22.11793 . ПМК 34351 . ПМИД 11050211 .
- ^ Петков К.И., Кайзер С., Аугат М., Логотетис Н.К. (июль 2006 г.). «Функциональная визуализация выявляет многочисленные поля в слуховой коре обезьян» . ПЛОС Биология . 4 (7): е215. doi : 10.1371/journal.pbio.0040215 . ПМЦ 1479693 . ПМИД 16774452 .
- ^ Морел А., Гаррагти П.Е., Каас Дж.Х. (сентябрь 1993 г.). «Тонотопическая организация, архитектонические поля и связи слуховой коры у макак». Журнал сравнительной неврологии . 335 (3): 437–59. дои : 10.1002/cne.903350312 . ПМИД 7693772 . S2CID 22872232 .
- ^ Раушекер Дж. П., Тиан Б. (октябрь 2000 г.). «Механизмы и потоки обработки «что» и «где» в слуховой коре» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (22): 11800–6. Бибкод : 2000PNAS...9711800R . дои : 10.1073/pnas.97.22.11800 . ПМК 34352 . ПМИД 11050212 .
- ^ Раушекер Дж. П., Тиан Б., Понс Т., Мишкин М. (май 1997 г.). «Последовательная и параллельная обработка данных в слуховой коре макак-резус». Журнал сравнительной неврологии . 382 (1): 89–103. doi : 10.1002/(sici)1096-9861(19970526)382:1<89::aid-cne6>3.3.co;2-y . ПМИД 9136813 .
- ^ Свит Р.А., Дорф-Петерсен К.А., Льюис Д.А. (октябрь 2005 г.). «Картирование слухового ядра, латерального пояса и парабелковой коры верхней височной извилины человека». Журнал сравнительной неврологии . 491 (3): 270–89. дои : 10.1002/cne.20702 . ПМИД 16134138 . S2CID 40822276 .
- ^ Уоллес М.Н., Джонстон П.В., Палмер А.Р. (апрель 2002 г.). «Гистохимическая идентификация корковых областей слуховой области головного мозга человека». Экспериментальное исследование мозга . 143 (4): 499–508. дои : 10.1007/s00221-002-1014-z . ПМИД 11914796 . S2CID 24211906 .
- ^ Да Коста С., ван дер Цвааг В., Маркес Дж.П., Фраковяк Р.С., Кларк С., Саенс М. (октябрь 2011 г.). «Первичная слуховая кора человека повторяет форму извилины Хешля» . Журнал неврологии . 31 (40): 14067–75. doi : 10.1523/jneurosci.2000-11.2011 . ПМК 6623669 . ПМИД 21976491 .
- ^ Хамфрис К., Либенталь Э., Биндер-младший (апрель 2010 г.). «Тонотопическая организация слуховой коры человека» . НейроИмидж . 50 (3): 1202–11. doi : 10.1016/j.neuroimage.2010.01.046 . ПМЦ 2830355 . ПМИД 20096790 .
- ^ Лангерс Д.Р., ван Дейк П. (сентябрь 2012 г.). «Картирование тонотопической организации слуховой коры человека при минимально выраженной акустической стимуляции» . Кора головного мозга . 22 (9): 2024–38. дои : 10.1093/cercor/bhr282 . ПМЦ 3412441 . ПМИД 21980020 .
- ^ Стрием-Амит Э., Герц У., Амеди А. (март 2011 г.). «Обширное кохлеотопическое картирование полей слуховой коры человека, полученное с помощью фМРТ с фазовым кодированием» . ПЛОС ОДИН . 6 (3): e17832. Бибкод : 2011PLoSO...617832S . дои : 10.1371/journal.pone.0017832 . ПМК 3063163 . ПМИД 21448274 .
- ^ Вудс Д.Л., Херрон Т.Дж., Кейт А.Д., Юнд Э.В., Стекер Г.К., Ринне Т., Канг X (2010). «Функциональные свойства полей слуховой коры человека» . Границы системной нейронауки . 4 : 155. дои : 10.3389/fnsys.2010.00155 . ПМК 3001989 . ПМИД 21160558 .
- ^ Гуревич Б., Ле Букен Жаннес Р., Фокон Ж., Льежуа-Шовель С. (март 2008 г.). «Обработка временной оболочки в слуховой коре человека: реакция и взаимосвязи областей слуховой коры» (PDF) . Исследование слуха . 237 (1–2): 1–18. дои : 10.1016/j.heares.2007.12.003 . ПМИД 18255243 . S2CID 15271578 .
- ^ Геген М., Ле Букен-Жаннес Р., Фокон Ж., Шовель П., Льежуа-Шовель С. (февраль 2007 г.). «Доказательства функциональной связи между областями слуховой коры, выявленные при обработке звука с амплитудной модуляцией» . Кора головного мозга . 17 (2): 304–13. дои : 10.1093/cercor/bhj148 . ПМК 2111045 . ПМИД 16514106 .
- ^ Перейти обратно: а б с Полива О, Бестельмейер П.Е., Холл М., Бултитьюд Дж.Х., Коллер К., Рафаль Р.Д. (сентябрь 2015 г.). «Функциональное картирование слуховой коры человека: фМРТ-исследование пациента со слуховой агнозией от травмы нижнего холмика» (PDF) . Когнитивная и поведенческая неврология . 28 (3): 160–80. doi : 10.1097/wnn.0000000000000072 . ПМИД 26413744 . S2CID 913296 .
- ^ Чанг Э.Ф., Эдвардс Э., Нагараджан СС, Фогельсон Н., Далал СС, Канолти РТ, Кирш Х.Э., Барбаро Н.М., Найт РТ (июнь 2011 г.). «Корковая пространственно-временная динамика, лежащая в основе обнаружения фонологических целей у человека» . Журнал когнитивной нейронауки . 23 (6): 1437–46. дои : 10.1162/jocn.2010.21466 . ПМЦ 3895406 . ПМИД 20465359 .
- ^ Муньос М., Мишкин М., Сондерс Р.К. (сентябрь 2009 г.). «Резекция медиальной височной доли отключает ростральную верхнюю височную извилину от некоторых ее проекционных целей в лобной доле и таламусе» . Кора головного мозга . 19 (9): 2114–30. дои : 10.1093/cercor/bhn236 . ПМЦ 2722427 . ПМИД 19150921 .
- ^ Перейти обратно: а б Романски Л.М., Бейтс Дж.Ф., Гольдман-Ракич П.С. (январь 1999 г.). «Слуховой пояс и парабелтные проекции префронтальной коры макаки-резуса». Журнал сравнительной неврологии . 403 (2): 141–57. doi : 10.1002/(sici)1096-9861(19990111)403:2<141::aid-cne1>3.0.co;2-v . ПМИД 9886040 . S2CID 42482082 .
- ^ Танака Д. (июнь 1976 г.). «Таламические проекции дорсомедиальной префронтальной коры макаки-резуса (Macaca mulatta)». Исследования мозга . 110 (1): 21–38. дои : 10.1016/0006-8993(76)90206-7 . ПМИД 819108 . S2CID 21529048 .
- ^ Перейти обратно: а б Перродин С., Кайзер С., Логотетис Н.К., Петков С.И. (август 2011 г.). «Голосовые клетки височной доли приматов» . Современная биология . 21 (16): 1408–15. дои : 10.1016/j.cub.2011.07.028 . ПМК 3398143 . ПМИД 21835625 .
- ^ Перейти обратно: а б с Петков К.И., Кайзер С., Стодел Т., Уиттингстолл К., Огат М., Логотетис Н.К. (март 2008 г.). «Голосовая область в мозгу обезьяны». Природная неврология . 11 (3): 367–74. дои : 10.1038/nn2043 . ПМИД 18264095 . S2CID 5505773 .
- ^ Перейти обратно: а б Поремба А., Маллой М., Сондерс Р.К., Карсон Р.Э., Херскович П., Мишкин М. (январь 2004 г.). «Видоспецифичные крики вызывают асимметричную активность височных полюсов обезьяны». Природа . 427 (6973): 448–51. Бибкод : 2004Natur.427..448P . дои : 10.1038/nature02268 . ПМИД 14749833 . S2CID 4402126 .
- ^ Романски Л.М., Авербек Б.Б., Дилц М. (февраль 2005 г.). «Нейронное представление вокализаций в вентролатеральной префронтальной коре приматов». Журнал нейрофизиологии . 93 (2): 734–47. дои : 10.1152/jn.00675.2004 . ПМИД 15371495 .
- ^ Расс Б.Е., Акельсон А.Л., Бейкер А.Е., Коэн Ю.Е. (январь 2008 г.). «Кодирование идентичности слухового стимула в потоке слуховой непространственной обработки» . Журнал нейрофизиологии . 99 (1): 87–95. дои : 10.1152/Jn.01069.2007 . ПМК 4091985 . ПМИД 18003874 .
- ^ Перейти обратно: а б Цунада Дж., Ли Дж. Х., Коэн Ю. Е. (июнь 2011 г.). «Представление речевых категорий в слуховой коре приматов» . Журнал нейрофизиологии . 105 (6): 2634–46. дои : 10.1152/jn.00037.2011 . ПМЦ 3118748 . ПМИД 21346209 .
- ^ Кьюсик К.Г., Зельцер Б., Кола М., Григгс Э. (сентябрь 1995 г.). «Хемоархитектоника и корково-кортикальные окончания в верхней височной борозде макаки-резуса: свидетельства подразделения верхней височной полисенсорной коры». Журнал сравнительной неврологии . 360 (3): 513–35. дои : 10.1002/cne.903600312 . ПМИД 8543656 . S2CID 42281619 .
- ^ Коэн Ю.Е., Расс Б.Е., Гиффорд Г.В., Кирингода Р., Маклин К.А. (декабрь 2004 г.). «Избирательность пространственных и непространственных атрибутов слуховых стимулов в вентролатеральной префронтальной коре» . Журнал неврологии . 24 (50): 11307–16. doi : 10.1523/jneurosci.3935-04.2004 . ПМК 6730358 . ПМИД 15601937 .
- ^ Диакон Т.В. (февраль 1992 г.). «Корковые соединения коры нижней дугообразной борозды мозга макака». Исследования мозга . 573 (1): 8–26. дои : 10.1016/0006-8993(92)90109-м . ISSN 0006-8993 . ПМИД 1374284 . S2CID 20670766 .
- ^ Льюис Дж.В., Ван Эссен, округ Колумбия (декабрь 2000 г.). «Кортикокортикальные связи зрительных, сенсомоторных и мультимодальных областей обработки в теменной доле макаки». Журнал сравнительной неврологии . 428 (1): 112–37. doi : 10.1002/1096-9861(20001204)428:1<112::aid-cne8>3.0.co;2-9 . ПМИД 11058227 . S2CID 16153360 .
- ^ Робертс AC, Томик Д.Л., Паркинсон CH, Роелинг Т.А., Каттер DJ, Роббинс Т.В., Эверитт Б.Дж. (май 2007 г.). «Связность переднего мозга префронтальной коры у мартышки (Callithrix jacchus): исследование антероградного и ретроградного отслеживания путей». Журнал сравнительной неврологии . 502 (1): 86–112. дои : 10.1002/cne.21300 . ПМИД 17335041 . S2CID 18262007 .
- ^ Перейти обратно: а б Шмахманн Дж.Д., Пандия Д.Н., Ван Р., Дай Дж., Д'Арсей Х.Э., де Креспиньи А.Дж., Ведин В.Дж. (март 2007 г.). «Ассоциативные пути волокон головного мозга: параллельные наблюдения с помощью визуализации диффузионного спектра и авторадиографии» . Мозг . 130 (Часть 3): 630–53. дои : 10.1093/brain/awl359 . ПМИД 17293361 .
- ^ Зельцер Б., Пандия Д.Н. (июль 1984 г.). «Дальнейшие наблюдения над теменно-височными связями у макак-резус». Экспериментальное исследование мозга . 55 (2): 301–12. дои : 10.1007/bf00237280 . ПМИД 6745368 . S2CID 20167953 .
- ^ Катани М., Джонс Д.К., Фитче Д.Х. (январь 2005 г.). «Перисильвийские языковые сети человеческого мозга» . Анналы неврологии . 57 (1): 8–16. дои : 10.1002/ana.20319 . ПМИД 15597383 . S2CID 17743067 .
- ^ Фрей С., Кэмпбелл Дж. С., Пайк ГБ, Петридес М. (ноябрь 2008 г.). «Диссоциация путей человеческой речи с помощью диффузионной волоконной трактографии с высоким угловым разрешением» . Журнал неврологии . 28 (45): 11435–44. doi : 10.1523/jneurosci.2388-08.2008 . ПМК 6671318 . ПМИД 18987180 .
- ^ Макрис Н., Пападимитриу ГМ, Кайзер-младший, Сорг С., Кеннеди Д.Н., Пандия Д.Н. (апрель 2009 г.). «Ограничение среднего продольного пучка у человека: количественное исследование in vivo с помощью DT-MRI» . Кора головного мозга . 19 (4): 777–85. дои : 10.1093/cercor/bhn124 . ПМК 2651473 . ПМИД 18669591 .
- ^ Менжо де Шампфлер Н., Лима Мальдонадо И., Мориц-Гассер С., Мачи П., Ле Барс Е., Бонафе А., Даффо Х. (январь 2013 г.). «Очертание среднего продольного пучка речевых путей: исследование тензорной диффузионной визуализации у человека» . Европейский журнал радиологии . 82 (1): 151–7. дои : 10.1016/j.ejrad.2012.05.034 . ПМИД 23084876 .
- ^ Туркен АУ, Дронкерс НФ (2011). «Нейронная архитектура сети понимания языка: сходящиеся данные анализа повреждений и связности» . Границы системной нейронауки . 5 : 1. дои : 10.3389/fnsys.2011.00001 . ПМК 3039157 . ПМИД 21347218 .
- ^ Саур Д., Креер Б.В., Шнелл С., Кюммерер Д., Келлмейер П., Ври М.С., Умарова Р., Муссо М., Глауче В., Абель С., Хубер В., Райнтьес М., Хенниг Дж., Вейллер С. (ноябрь 2008 г.). «Вентральные и дорсальные пути речи» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (46): 18035–40. Бибкод : 2008PNAS..10518035S . дои : 10.1073/pnas.0805234105 . ПМЦ 2584675 . ПМИД 19004769 .
- ^ Инь П., Мишкин М., Саттер М., Фриц Дж.Б. (декабрь 2008 г.). «Ранние стадии обработки мелодии: последовательность стимулов и зависящая от задачи активность нейронов в полях слуховой коры обезьян A1 и R» . Журнал нейрофизиологии . 100 (6): 3009–29. дои : 10.1152/jn.00828.2007 . ПМК 2604844 . ПМИД 18842950 .
- ^ Штайншнейдер М., Волков И.О., Фишман Ю.И., Оя Х., Ареццо Дж.К., Ховард М.А. (февраль 2005 г.). «Внутрикорковые реакции в первичной слуховой коре человека и обезьяны поддерживают механизм временной обработки для кодирования фонетического параметра времени начала голоса» . Кора головного мозга . 15 (2): 170–86. дои : 10.1093/cercor/bhh120 . ПМИД 15238437 .
- ^ Расс Б.Е., Акельсон А.Л., Бейкер А.Е., Коэн Ю.Е. (январь 2008 г.). «Кодирование идентичности слухового стимула в потоке слуховой непространственной обработки» . Журнал нейрофизиологии . 99 (1): 87–95. дои : 10.1152/Jn.01069.2007 . ПМК 4091985 . ПМИД 18003874 .
- ^ Жоли О., Пальер С., Рамус Ф., Прессницер Д., Вандуффель В., Орбан Г.А. (сентябрь 2012 г.). «Обработка вокализаций у людей и обезьян: сравнительное исследование фМРТ» (PDF) . НейроИмидж . 62 (3): 1376–89. doi : 10.1016/j.neuroimage.2012.05.070 . ПМИД 22659478 . S2CID 9441377 .
- ^ Шейх Х., Баумгарт Ф., Гашлер-Маркефски Б., Тегелер С., Темпельманн С., Хайнце Х.Дж., Шиндлер Ф., Стиллер Д. (февраль 1998 г.). «Функциональная магнитно-резонансная томография области слуховой коры человека, участвующей в разложении переднего плана и фона». Европейский журнал неврологии . 10 (2): 803–9. дои : 10.1046/j.1460-9568.1998.00086.x . ПМИД 9749748 . S2CID 42898063 .
- ^ Заторре Р.Дж., Буффар М., Белин П. (апрель 2004 г.). «Чувствительность к особенностям слуховых объектов в височной коре головного мозга человека» . Журнал неврологии . 24 (14): 3637–42. doi : 10.1523/jneurosci.5458-03.2004 . ПМК 6729744 . ПМИД 15071112 .
- ^ Перейти обратно: а б Биндер-младший, Десаи Р.Х., Грейвс В.В., Конант Л.Л. (декабрь 2009 г.). «Где семантическая система? Критический обзор и метаанализ 120 исследований функциональной нейровизуализации» . Кора головного мозга . 19 (12): 2767–96. дои : 10.1093/cercor/bhp055 . ПМЦ 2774390 . ПМИД 19329570 .
- ^ Дэвис М.Х., Джонсруд И.С. (апрель 2003 г.). «Иерархическая обработка при понимании разговорной речи» . Журнал неврологии . 23 (8): 3423–31. doi : 10.1523/jneurosci.23-08-03423.2003 . ПМК 6742313 . ПМИД 12716950 .
- ^ Либенталь Э., Биндер-младший, Спитцер С.М., Поссинг Э.Т., Медлер Д.А. (октябрь 2005 г.). «Нейронные субстраты фонематического восприятия» . Кора головного мозга . 15 (10): 1621–31. дои : 10.1093/cercor/bhi040 . ПМИД 15703256 .
- ^ Нараин С., Скотт С.К., Уайз Р.Дж., Розен С., Лефф А., Иверсен С.Д., Мэтьюз П.М. (декабрь 2003 г.). «Определение леволатеральной реакции, специфичной для разборчивой речи, с помощью фМРТ» . Кора головного мозга . 13 (12): 1362–8. дои : 10.1093/cercor/bhg083 . ПМИД 14615301 .
- ^ Облесер Дж., Бекер Х., Джезга А., Хаслингер Б., Хенненлоттер А., Реттингер М., Юлиц С., Раушекер Дж. П. (июль 2006 г.). «Извлечение гласных звуков в передней верхней височной коре» . Картирование человеческого мозга . 27 (7): 562–71. дои : 10.1002/hbm.20201 . ПМК 6871493 . ПМИД 16281283 .
- ^ Облесер Дж., Циммерманн Дж., Ван Метер Дж., Раушекер Дж. П. (октябрь 2007 г.). «Множественные стадии слухового восприятия речи, отраженные в событийно-связанном FMRI» . Кора головного мозга . 17 (10): 2251–7. дои : 10.1093/cercor/bhl133 . ПМИД 17150986 .
- ^ Скотт С.К., Бланк CC, Розен С., Уайз Р.Дж. (декабрь 2000 г.). «Определение пути разборчивой речи в левой височной доле» . Мозг . 123 (12): 2400–6. дои : 10.1093/мозг/123.12.2400 . ПМК 5630088 . ПМИД 11099443 .
- ^ Белин П., Заторре Р.Дж. (ноябрь 2003 г.). «Адаптация к голосу говорящего в правой передней височной доле». НейроОтчет . 14 (16): 2105–2109. дои : 10.1097/00001756-200311140-00019 . ПМИД 14600506 . S2CID 34183900 .
- ^ Бенсон Р.Р., Уэлен Д.Х., Ричардсон М., Суэйнсон Б., Кларк В.П., Лай С., Либерман А.М. (сентябрь 2001 г.). «Параметрическое разделение речевого и неречевого восприятия в мозге с помощью фМРТ». Мозг и язык . 78 (3): 364–96. дои : 10.1006/brln.2001.2484 . ПМИД 11703063 . S2CID 15328590 .
- ^ Ливер А.М., Раушекер Дж.П. (июнь 2010 г.). «Корковое представление сложных естественных звуков: влияние акустических особенностей и категорий слуховых объектов» . Журнал неврологии . 30 (22): 7604–12. doi : 10.1523/jneurosci.0296-10.2010 . ПМЦ 2930617 . ПМИД 20519535 .
- ^ Льюис Дж.В., Финни Р.Э., Брефчински-Льюис Дж.А., ДеЙо Э.А. (август 2006 г.). «Левши понимают «правильно», когда слышат звуки инструментов». Журнал когнитивной нейронауки . 18 (8): 1314–30. дои : 10.1162/jocn.2006.18.8.1314 . ПМИД 16859417 . S2CID 14049095 .
- ^ Маедер П.П., Меули Р.А., Адриани М., Беллманн А., Форнари Э., Тиран Дж.П., Питтет А., Кларк С. (октябрь 2001 г.). «Различные пути, участвующие в распознавании и локализации звука: исследование фМРТ человека» (PDF) . НейроИмидж . 14 (4): 802–16. дои : 10.1006/нимг.2001.0888 . ПМИД 11554799 . S2CID 1388647 .
- ^ Вицеич Д., Форнари Э., Тиран Дж.П., Маедер П.П., Меули Р., Адриани М., Кларк С. (ноябрь 2006 г.). «Области слухового пояса человека, специализирующиеся на распознавании звуков: исследование функциональной магнитно-резонансной томографии» (PDF) . НейроОтчет . 17 (16): 1659–62. doi : 10.1097/01.wnr.0000239962.75943.dd . ПМИД 17047449 . S2CID 14482187 .
- ^ Шульц С., Вулуманос А., Пелфри К. (май 2012 г.). «Верхняя височная борозда различает коммуникативные и некоммуникативные слуховые сигналы». Журнал когнитивной нейронауки . 24 (5): 1224–32. дои : 10.1162/jocn_a_00208 . ПМИД 22360624 . S2CID 10784270 .
- ^ ДеВитт И., Раушекер Дж. П. (февраль 2012 г.). «Распознавание фонем и слов в слуховом вентральном потоке» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (8): Е505-14. Бибкод : 2012PNAS..109E.505D . дои : 10.1073/pnas.1113427109 . ПМК 3286918 . ПМИД 22308358 .
- ^ Перейти обратно: а б с д Лашо Дж.П., Джерби К., Бертран О., Минотти Л., Хоффманн Д., Шендорф Б., Кахане П. (октябрь 2007 г.). «Проект функционального картирования в реальном времени с помощью внутричерепных записей человека» . ПЛОС ОДИН . 2 (10): е1094. Бибкод : 2007PLoSO...2.1094L . дои : 10.1371/journal.pone.0001094 . ПМК 2040217 . ПМИД 17971857 .
- ^ Перейти обратно: а б Мацумото Р., Имамура Х., Инучи М., Накагава Т., Ёкояма Ю., Мацухаши М., Микуни Н., Миямото С., Фукуяма Х., Такахаши Р., Икеда А. (апрель 2011 г.). «Левая передняя височная кора активно участвует в восприятии речи: исследование прямой кортикальной стимуляции». Нейропсихология . 49 (5): 1350–1354. doi : 10.1016/j.neuropsychologia.2011.01.023 . hdl : 2433/141342 . ПМИД 21251921 . S2CID 1831334 .
- ^ Перейти обратно: а б с д Ру Ф.Е., Мискин К., Дюран Ж.Б., Сако О., Рео Э., Танова Р., Демонет Ж.Ф. (октябрь 2015 г.). «Электростимуляционное картирование понимания слуховых и зрительных слов». Кора; Журнал, посвященный изучению нервной системы и поведения . 71 : 398–408. дои : 10.1016/j.cortex.2015.07.001 . ПМИД 26332785 . S2CID 39964328 .
- ^ Фриц Дж., Мишкин М., Сондерс Р.К. (июнь 2005 г.). «В поисках аудиальной инграммы» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (26): 9359–64. Бибкод : 2005PNAS..102.9359F . дои : 10.1073/pnas.0503998102 . ПМЦ 1166637 . ПМИД 15967995 .
- ^ Степьен Л.С., Кордо Дж.П., Расмуссен Т. (1960). «Влияние поражений височной доли и гиппокампа на слуховую и зрительную недавнюю память у обезьян» . Мозг . 83 (3): 470–489. дои : 10.1093/мозг/83.3.470 . ISSN 0006-8950 .
- ^ Стромингер Н.Л., Остеррайх Р.Э., Нефф В.Д. (июнь 1980 г.). «Последовательная слуховая и зрительная дискриминация после абляции височной доли у обезьян». Физиология и поведение . 24 (6): 1149–56. дои : 10.1016/0031-9384(80)90062-1 . ПМИД 6774349 . S2CID 7494152 .
- ^ Кайзер Дж., Риппер Б., Бирбаумер Н., Луценбергер В. (октябрь 2003 г.). «Динамика активности гамма-диапазона в магнитоэнцефалограмме человека при работе слуховой модели рабочей памяти». НейроИмидж . 20 (2): 816–27. дои : 10.1016/s1053-8119(03)00350-1 . PMID 14568454 . S2CID 19373941 .
- ^ Бухсбаум Б.Р., Олсен Р.К., Кох П., Берман К.Ф. (ноябрь 2005 г.). «Спинные и вентральные слуховые потоки человека служат репетиционным и эхическим процессам во время вербальной рабочей памяти» . Нейрон . 48 (4): 687–97. дои : 10.1016/j.neuron.2005.09.029 . ПМИД 16301183 . S2CID 13202604 .
- ^ Скотт Б.Х., Мишкин М., Инь П. (июль 2012 г.). «Обезьяны имеют ограниченную форму кратковременной памяти на прослушивании» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (30): 12237–41. Бибкод : 2012PNAS..10912237S . дои : 10.1073/pnas.1209685109 . ПМК 3409773 . ПМИД 22778411 .
- ^ Ноппени У., Паттерсон К., Тайлер Л.К., Мосс Х., Стаматакис Э.А., Брайт П., Маммери С., Прайс СиДжей (апрель 2007 г.). «Поражения височных долей и семантические нарушения: сравнение энцефалита, вызванного вирусом простого герпеса, и семантической деменции» . Мозг . 130 (Часть 4): 1138–47. дои : 10.1093/brain/awl344 . ПМИД 17251241 .
- ^ Паттерсон К., Нестор П.Дж., Роджерс Т.Т. (декабрь 2007 г.). «Откуда вы знаете то, что знаете? Представление семантических знаний в человеческом мозгу». Обзоры природы. Нейронаука . 8 (12): 976–87. дои : 10.1038/nrn2277 . ПМИД 18026167 . S2CID 7310189 .
- ^ Шварц М.Ф., Кимберг Д.Ю., Уокер Г.М., Фасейтан О., Бречер А., Делл Г.С., Кослетт Х.Б. (декабрь 2009 г.). «Вовлечение передней височной части в семантический поиск слов: свидетельства картирования симптомов и поражений на основе вокселей при афазии» . Мозг . 132 (Часть 12): 3411–27. дои : 10.1093/brain/awp284 . ПМЦ 2792374 . ПМИД 19942676 .
- ^ Хамбергер М.Дж., Макклелланд С., Макханн Г.М., Уильямс А.К., Гудман Р.Р. (март 2007 г.). «Распределение слуховых и зрительных участков именования у пациентов с неочищенной височной эпилепсией и пациентов с объемными поражениями височной доли» . Эпилепсия . 48 (3): 531–8. дои : 10.1111/j.1528-1167.2006.00955.x . ПМИД 17326797 . S2CID 12642281 .
- ^ Перейти обратно: а б с Даффо Х (март 2008 г.). «Возврат к анатомо-функциональной связности языка. Новые идеи, полученные с помощью электростимуляции и трактографии». Нейропсихология . 46 (4): 927–34. doi : 10.1016/j.neuropsychologia.2007.10.025 . ПМИД 18093622 . S2CID 40514753 .
- ^ Виньо М., Бокузен В., Эрве П.Ю., Дюффо Х., Кривелло Ф., Уде О., Мазойер Б., Цурио-Мазуайер Н. (май 2006 г.). «Метаанализ языковых областей левого полушария: фонология, семантика и обработка предложений». НейроИмидж . 30 (4): 1414–32. doi : 10.1016/j.neuroimage.2005.11.002 . ПМИД 16413796 . S2CID 8870165 .
- ^ Перейти обратно: а б Крейцфельдт О, Ожеманн Г, Леттич Э (октябрь 1989 г.). «Нейрональная активность в боковой височной доле человека. I. Реакция на речь». Экспериментальное исследование мозга . 77 (3): 451–75. дои : 10.1007/bf00249600 . hdl : 11858/00-001M-0000-002C-89EA-3 . ПМИД 2806441 . S2CID 19952034 .
- ^ Мазойер Б.М., Цурио Н., Фрак В., Сирота А., Мураяма Н., Левье О., Саламон Г., Дехан С., Коэн Л., Мелер Дж. (октябрь 1993 г.). «Корковое представление речи» (PDF) . Журнал когнитивной нейронауки . 5 (4): 467–79. дои : 10.1162/jocn.1993.5.4.467 . ПМИД 23964919 . S2CID 22265355 .
- ^ Хамфрис С., Лав Т., Суинни Д., Хикок Дж. (октябрь 2005 г.). «Реакция передней височной коры на синтаксические и просодические манипуляции во время обработки предложения» . Картирование человеческого мозга . 26 (2): 128–38. дои : 10.1002/hbm.20148 . ПМК 6871757 . ПМИД 15895428 .
- ^ Хамфрис К., Уиллард К., Буксбаум Б., Хикок Дж. (июнь 2001 г.). «Роль передней височной коры в понимании слуховых предложений: исследование фМРТ». НейроОтчет . 12 (8): 1749–52. дои : 10.1097/00001756-200106130-00046 . ПМИД 11409752 . S2CID 13039857 .
- ^ Ванденберге Р., Нобре AC, Прайс CJ (май 2002 г.). «Реакция левой височной коры на предложения». Журнал когнитивной нейронауки . 14 (4): 550–60. дои : 10.1162/08989290260045800 . ПМИД 12126497 . S2CID 21607482 .
- ^ Фридеричи А.Д., Рюшемейер С.А., Хане А., Фибах С.Дж. (февраль 2003 г.). «Роль левой нижней лобной и верхней височной коры в понимании предложений: локализация синтаксических и семантических процессов» . Кора головного мозга . 13 (2): 170–7. дои : 10.1093/cercor/13.2.170 . hdl : 11858/00-001M-0000-0010-E3AB-B . ПМИД 12507948 .
- ^ Сюй Дж., Кемени С., Пак Дж., Фраттали С., Браун А. (2005). «Язык в контексте: новые особенности понимания слов, предложений и повествования». НейроИмидж . 25 (3): 1002–15. doi : 10.1016/j.neuroimage.2004.12.013 . ПМИД 15809000 . S2CID 25570583 .
- ^ Рогальский С., Хикок Г. (апрель 2009 г.). «Избирательное внимание к семантическим и синтаксическим особенностям модулирует сети обработки предложений в передней височной коре» . Кора головного мозга . 19 (4): 786–96. дои : 10.1093/cercor/bhn126 . ПМЦ 2651476 . ПМИД 18669589 .
- ^ Пальер С., Девошель А.Д., Деэн С. (февраль 2011 г.). «Корковое представление составной структуры предложения» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (6): 2522–7. дои : 10.1073/pnas.1018711108 . ПМК 3038732 . ПМИД 21224415 .
- ^ Бреннан Дж., Нир Ю., Хассон У., Малах Р., Хигер Д.Д., Пюлкканен Л. (февраль 2012 г.). «Построение синтаксической структуры в передней височной доле при прослушивании естественного рассказа» . Мозг и язык . 120 (2): 163–73. дои : 10.1016/j.bandl.2010.04.002 . ПМЦ 2947556 . ПМИД 20472279 .
- ^ Коц С.А., фон Крамон Д.Ю., Фридеричи А.Д. (октябрь 2003 г.). «Дифференциация синтаксических процессов в левой и правой передней височной доле: данные о потенциале мозга, связанные с событиями, от пациентов с поражением». Мозг и язык . 87 (1): 135–136. дои : 10.1016/s0093-934x(03)00236-0 . S2CID 54320415 .
- ^ Мартин Р.К., Шелтон-младший, Яффи Л.С. (февраль 1994 г.). «Языковая обработка и рабочая память: нейропсихологические доказательства отдельных фонологических и семантических способностей». Журнал памяти и языка . 33 (1): 83–111. дои : 10.1006/jmla.1994.1005 .
- ^ Магнусдоттир С., Филлмор П., ден Оуден Д.Б., Хьялтасон Х., Рорден С., Кьяртанссон О., Бонилья Л., Фридрикссон Дж. (октябрь 2013 г.). «Повреждение левой передней височной коры предсказывает нарушение сложной синтаксической обработки: исследование картирования симптомов поражения» . Картирование человеческого мозга . 34 (10): 2715–23. дои : 10.1002/hbm.22096 . ПМК 6869931 . ПМИД 22522937 .
- ^ Борнкессель-Шлезевски И., Шлезевски М., Смолл С.Л., Раушекер Дж.П. (март 2015 г.). «Нейробиологические корни языка при слухе у приматов: общие вычислительные свойства» . Тенденции в когнитивных науках . 19 (3): 142–50. дои : 10.1016/j.tics.2014.12.008 . ПМК 4348204 . ПМИД 25600585 .
- ^ Хикок Г., Окада К., Барр В., Па Дж., Рогальски С., Доннелли К., Бард Л., Грант А. (декабрь 2008 г.). «Двусторонняя способность обработки звуков речи при слуховом понимании: данные процедур ВАДА» . Мозг и язык . 107 (3): 179–84. дои : 10.1016/j.bandl.2008.09.006 . ПМК 2644214 . ПМИД 18976806 .
- ^ Зайдель Э. (сентябрь 1976 г.). «Слуховой словарь правого полушария после рассечения мозга или гемидекортикации» . Кортекс . 12 (3): 191–211. дои : 10.1016/s0010-9452(76)80001-9 . ISSN 0010-9452 . ПМИД 1000988 . S2CID 4479925 .
- ^ Поппель Д. (октябрь 2001 г.). «Чистая словесная глухота и двусторонняя обработка речевого кода» . Когнитивная наука . 25 (5): 679–693. дои : 10.1016/s0364-0213(01)00050-7 .
- ^ Ульрих Г. (май 1978 г.). «Межполушарные функциональные взаимоотношения при слуховой агнозии. Анализ предпосылок и концептуальная модель». Мозг и язык . 5 (3): 286–300. дои : 10.1016/0093-934x(78)90027-5 . ПМИД 656899 . S2CID 33841186 .
- ^ Стюарт Л., Уолш В., Фрит У., Ротвелл Дж. К. (март 2001 г.). «ТМС вызывает два разделяемых типа нарушений речи» (PDF) . НейроИмидж . 13 (3): 472–8. дои : 10.1006/нимг.2000.0701 . ПМИД 11170812 . S2CID 10392466 .
- ^ Ачесон DJ, Хамиди М, Биндер JR, Postle BR (июнь 2011 г.). «Общий нейронный субстрат для производства речи и вербальной рабочей памяти» . Журнал когнитивной нейронауки . 23 (6): 1358–67. дои : 10.1162/jocn.2010.21519 . ПМК 3053417 . ПМИД 20617889 .
- ^ Десмургет М., Рейли К.Т., Ричард Н., Сатмари А., Моттолезе С., Сиригу А. (май 2009 г.). «Намерение движения после стимуляции теменной коры у людей». Наука . 324 (5928): 811–3. Бибкод : 2009Sci...324..811D . дои : 10.1126/science.1169896 . ПМИД 19423830 . S2CID 6555881 .
- ^ Эдвардс Э., Нагараджан СС, Далал СС, Канолти РТ, Кирш Х.Э., Барбаро Н.М., Найт РТ (март 2010 г.). «Пространственно-временная визуализация активации коры во время генерации глаголов и называния изображений» . НейроИмидж . 50 (1): 291–301. doi : 10.1016/j.neuroimage.2009.12.035 . ПМЦ 2957470 . ПМИД 20026224 .
- ^ Ботман Д., Гордон Б., Харт Дж., Селнес О., Мильоретти Д. , Ленц Ф. (август 2000 г.). «Транскортикальная сенсорная афазия: пересмотренная и пересмотренная» . Мозг . 123 (8): 1634–42. дои : 10.1093/мозг/123.8.1634 . ПМИД 10908193 .
- ^ Перейти обратно: а б с Туркельтауб PE, Кослетт HB (июль 2010 г.). «Локализация сублексических компонентов восприятия речи» . Мозг и язык . 114 (1): 1–15. дои : 10.1016/j.bandl.2010.03.008 . ПМЦ 2914564 . ПМИД 20413149 .
- ^ Чанг Э.Ф., Ригер Дж.В., Джонсон К., Бергер М.С., Барбаро Н.М., Найт RT (ноябрь 2010 г.). «Категорическое речевое представление в верхней височной извилине человека» . Природная неврология . 13 (11): 1428–32. дои : 10.1038/nn.2641 . ПМЦ 2967728 . ПМИД 20890293 .
- ^ Бухсбаум Б.Р., Хикок Г., Хамфрис К. (сентябрь 2001 г.). «Роль левой задней верхней височной извилины в фонологической обработке восприятия и производства речи» . Когнитивная наука . 25 (5): 663–678. дои : 10.1207/s15516709cog2505_2 . ISSN 0364-0213 .
- ^ Уайз Р.Дж., Скотт С.К., Бланк СК, Маммери СиДжей, Мерфи К., Уорбертон Э.А. (январь 2001 г.). «Отдельные нейронные подсистемы в «зоне Вернике» » . Мозг . 124 (Часть 1): 83–95. дои : 10.1093/мозг/124.1.83 . ПМИД 11133789 .
- ^ Хикок Г., Буксбаум Б., Хамфрис С., Муфтулер Т. (июль 2003 г.). «Слухо-моторное взаимодействие, выявленное с помощью фМРТ: речь, музыка и рабочая память в области Spt». Журнал когнитивной нейронауки . 15 (5): 673–82. дои : 10.1162/089892903322307393 . ПМИД 12965041 .
- ^ Уоррен Дж. Э., Уайз Р. Дж., Уоррен Дж. Д. (декабрь 2005 г.). «Звучит выполнимо: слухо-моторные трансформации и задняя височная плоскость». Тенденции в нейронауках . 28 (12): 636–43. дои : 10.1016/j.tins.2005.09.010 . ПМИД 16216346 . S2CID 36678139 .
- ^ Хикок Г., Поппель Д. (май 2007 г.). «Корковая организация обработки речи». Обзоры природы. Нейронаука . 8 (5): 393–402. дои : 10.1038/nrn2113 . ПМИД 17431404 . S2CID 6199399 .
- ^ Карбе Х., Герхольц К., Вебер-Люксенбургер Г., Гаеми М., Хейсс В.Д. (июнь 1998 г.). «Церебральные сети и функциональная асимметрия мозга: данные о региональных метаболических изменениях во время повторения слов». Мозг и язык . 63 (1): 108–21. дои : 10.1006/брлн.1997.1937 . ПМИД 9642023 . S2CID 31335617 .
- ^ Жиро А.Л., Прайс CJ (август 2001 г.). «Ограничения функциональной нейровизуализации на классические модели слуховой обработки текста». Журнал когнитивной нейронауки . 13 (6): 754–65. дои : 10.1162/08989290152541421 . ПМИД 11564320 . S2CID 13916709 .
- ^ Грейвс В.В., Грабовски Т.Дж., Мехта С., Гупта П. (сентябрь 2008 г.). «Левая задняя верхняя височная извилина конкретно участвует в доступе к лексической фонологии» . Журнал когнитивной нейронауки . 20 (9): 1698–710. дои : 10.1162/jocn.2008.20113 . ПМК 2570618 . ПМИД 18345989 .
- ^ Перейти обратно: а б Таул В.Л., Юн Х.А., Кастель М., Эдгар Дж.К., Биассу Н.М., Фрим Д.М., Спайр Дж.П., Корман М.Х. (август 2008 г.). «Гамма-активность ECoG во время языкового задания: дифференциация экспрессивных и рецептивных областей речи» . Мозг . 131 (Часть 8): 2013–27. дои : 10.1093/brain/awn147 . ПМК 2724904 . ПМИД 18669510 .
- ^ Селнес О.А., Кнопман Д.С., Никкум Н., Рубенс А.Б. (июнь 1985 г.). «Критическая роль области Вернике в повторении предложений». Анналы неврологии . 17 (6): 549–57. дои : 10.1002/ana.410170604 . ПМИД 4026225 . S2CID 12914191 .
- ^ Аксер Х., фон Кейзерлингк АГ, Беркс Г., фон Кейзерлингк Д.Г. (март 2001 г.). «Супра- и инфрасильвианская афазия проводимости». Мозг и язык . 76 (3): 317–31. дои : 10.1006/brln.2000.2425 . ПМИД 11247647 . S2CID 25406527 .
- ^ Барта Л., Бенке Т. (апрель 2003 г.). «Острая афазия проводимости: анализ 20 случаев». Мозг и язык . 85 (1): 93–108. дои : 10.1016/s0093-934x(02)00502-3 . ПМИД 12681350 . S2CID 18466425 .
- ^ Бальдо Й.В., Кацевф С., Дронкерс Н.Ф. (март 2012 г.). «Области мозга, лежащие в основе повторения и дефицита слухо-вербальной кратковременной памяти при афазии: данные картирования симптомов поражения на основе вокселей» . Афазиология . 26 (3–4): 338–354. дои : 10.1080/02687038.2011.602391 . ПМК 4070523 . ПМИД 24976669 .
- ^ Baldo JV, Klostermann EC, Dronkers NF (май 2008 г.). «Это либо повар, либо булочник: пациенты с афазией проводимости улавливают суть, но теряют след». Мозг и язык . 105 (2): 134–40. дои : 10.1016/j.bandl.2007.12.007 . ПМИД 18243294 . S2CID 997735 .
- ^ Фридрикссон Дж., Кьяртанссон О., Морган П.С., Хьялтасон Х., Магнусдоттир С., Бонилья Л., Рорден К. (август 2010 г.). «Нарушение повторения речи и повреждение левой теменной доли» . Журнал неврологии . 30 (33): 11057–61. doi : 10.1523/jneurosci.1120-10.2010 . ПМЦ 2936270 . ПМИД 20720112 .
- ^ Бухсбаум Б.Р., Бальдо Дж., Окада К., Берман К.Ф., Дронкерс Н., Д'Эспозито М., Хикок Г. (декабрь 2011 г.). «Кондуктивная афазия, сенсомоторная интеграция и фонологическая кратковременная память - совокупный анализ поражения и данные фМРТ» . Мозг и язык . 119 (3): 119–28. дои : 10.1016/j.bandl.2010.12.001 . ПМК 3090694 . ПМИД 21256582 .
- ^ Ямада К., Нагаканэ Ю., Мизуно Т., Хосоми А., Накагава М., Нисимура Т. (март 2007 г.). «МР-трактография, показывающая повреждение дугообразного пучка у пациента с проводниковой афазией» . Неврология . 68 (10): 789. doi : 10.1212/01.wnl.0000256348.65744.b2 . ПМИД 17339591 .
- ^ Брейер Дж.И., Хасан К.М., Чжан В., Мен Д., Папаниколау А.С. (март 2008 г.). «Языковая дисфункция после инсульта и повреждение участков белого вещества, оцениваемая с помощью диффузионно-тензорной визуализации» . АДЖНР. Американский журнал нейрорадиологии . 29 (3): 483–7. дои : 10.3174/ajnr.A0846 . ПМК 3073452 . ПМИД 18039757 .
- ^ Чжан И, Ван С, Чжао X, Чен Х, Хань Цз, Ван Ю (сентябрь 2010 г.). «Диффузионно-тензорная визуализация, изображающая повреждение дугообразного пучка у пациентов с афазией проводимости: исследование модели Вернике-Гешвинда». Неврологические исследования . 32 (7): 775–8. дои : 10.1179/016164109x12478302362653 . ПМИД 19825277 . S2CID 22960870 .
- ^ Джонс О.П., Прежава С., Хоуп Т.М., Оберхубер М., Сегир М.Л., Лефф А.П., Грин Д.В., Прайс CJ (2014). «Сенсорно-моторная интеграция во время слухового повторения: комбинированное исследование фМРТ и повреждений» . Границы человеческой неврологии . 8:24 . дои : 10.3389/fnhum.2014.00024 . ПМК 3908611 . ПМИД 24550807 .
- ^ Куигг М., Fountain NB (март 1999 г.). «Проводниковая афазия, вызванная стимуляцией левой задней верхней височной извилины» . Журнал неврологии, нейрохирургии и психиатрии . 66 (3): 393–6. дои : 10.1136/jnnp.66.3.393 . ПМЦ 1736266 . ПМИД 10084542 .
- ^ Куигг М., Гельдмахер Д.С., Элиас В.Дж. (май 2006 г.). «Кондуктивная афазия как функция доминирующей задней перисильвиевой коры. Отчет о двух случаях». Журнал нейрохирургии . 104 (5): 845–8. дои : 10.3171/jns.2006.104.5.845 . ПМИД 16703895 .
- ^ Сервис Е, Кохонен В (апрель 1995 г.). «Обусловлена ли связь между фонологической памятью и изучением иностранного языка приобретением словарного запаса?». Прикладная психолингвистика . 16 (2): 155–172. дои : 10.1017/S0142716400007062 . S2CID 143974128 .
- ^ Служба Е (июль 1992 г.). «Фонология, рабочая память и изучение иностранного языка». Ежеквартальный журнал экспериментальной психологии. А. Экспериментальная психология человека . 45 (1): 21–50. дои : 10.1080/14640749208401314 . ПМИД 1636010 . S2CID 43268252 .
- ^ Мацумото Р., Наир Д.Р., ЛаПресто Э., Наджм И., Бингаман В., Шибасаки Х., Людерс Х.О. (октябрь 2004 г.). «Функциональная связность в языковой системе человека: исследование кортико-кортикального вызванного потенциала» . Мозг . 127 (Часть 10): 2316–30. дои : 10.1093/brain/awh246 . ПМИД 15269116 .
- ^ Кимура Д., Уотсон Н. (ноябрь 1989 г.). «Связь между контролем оральных движений и речью». Мозг и язык . 37 (4): 565–90. дои : 10.1016/0093-934x(89)90112-0 . ПМИД 2479446 . S2CID 39913744 .
- ^ Турвиль Дж.А., Рейли К.Дж., Гюнтер Ф.Х. (февраль 2008 г.). «Нейронные механизмы, лежащие в основе слухового контроля речи» . НейроИмидж . 39 (3): 1429–43. doi : 10.1016/j.neuroimage.2007.09.054 . ПМЦ 3658624 . ПМИД 18035557 .
- ^ Чанг Э.Ф., Ригер Дж.В., Джонсон К., Бергер М.С., Барбаро Н.М., Найт RT (ноябрь 2010 г.). «Категорическое речевое представление в верхней височной извилине человека» . Природная неврология . 13 (11): 1428–32. дои : 10.1038/nn.2641 . ПМЦ 2967728 . ПМИД 20890293 .
- ^ Нат А.Р., Бошан М.С. (январь 2012 г.). «Нейронная основа межиндивидуальных различий в эффекте Макгерка, мультисенсорной речевой иллюзии» . НейроИмидж . 59 (1): 781–7. doi : 10.1016/j.neuroimage.2011.07.024 . ПМК 3196040 . ПМИД 21787869 .
- ^ Бошан М.С., Нат А.Р., Пасалар С. (февраль 2010 г.). «Транскраниальная магнитная стимуляция под контролем фМРТ показывает, что верхняя височная борозда является корковым локусом эффекта МакГерка» . Журнал неврологии . 30 (7): 2414–7. doi : 10.1523/JNEUROSCI.4865-09.2010 . ПМЦ 2844713 . ПМИД 20164324 .
- ^ МакГеттиган С., Фолкнер А., Алтарелли И., Облесер Дж., Баверсток Х., Скотт С.К. (апрель 2012 г.). «Понимание речи с помощью нескольких модальностей: поведенческих и нейронных взаимодействий» . Нейропсихология . 50 (5): 762–76. doi : 10.1016/j.neuropsychologia.2012.01.010 . ПМК 4050300 . ПМИД 22266262 .
- ^ Стивенсон Р.А., Джеймс Т.В. (февраль 2009 г.). «Аудиовизуальная интеграция в верхней височной борозде человека: обратная эффективность и нейронная обработка речи и распознавание объектов». НейроИмидж . 44 (3): 1210–23. doi : 10.1016/j.neuroimage.2008.09.034 . ПМИД 18973818 . S2CID 8342349 .
- ^ Бернштейн Л.Е., Цзян Дж., Пантазис Д., Лу З.Л., Джоши А. (октябрь 2011 г.). «Визуальная фонетическая обработка, локализованная с помощью речевых и неречевых жестов лица на видео и точечных световых дисплеях» . Картирование человеческого мозга . 32 (10): 1660–76. дои : 10.1002/hbm.21139 . ПМК 3120928 . ПМИД 20853377 .
- ^ Кэмпбелл Р. (март 2008 г.). «Обработка аудиовизуальной речи: эмпирические и нейронные основы» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 363 (1493): 1001–10. дои : 10.1098/rstb.2007.2155 . ПМК 2606792 . ПМИД 17827105 .
- ^ Шварц М.Ф., Фасейитан О., Ким Дж., Кослетт Х.Б. (декабрь 2012 г.). «Вклад спинного потока в фонологический поиск при именовании объектов» . Мозг . 135 (Часть 12): 3799–814. дои : 10.1093/brain/aws300 . ПМК 3525060 . ПМИД 23171662 .
- ^ Шварц М.Ф., Кимберг Д.Ю., Уокер Г.М., Фасейтан О., Бречер А., Делл Г.С., Кослетт Х.Б. (декабрь 2009 г.). «Вовлечение передней височной части в семантический поиск слов: свидетельства картирования симптомов и поражений на основе вокселей при афазии» . Мозг . 132 (Часть 12): 3411–27. дои : 10.1093/brain/awp284 . ПМЦ 2792374 . ПМИД 19942676 .
- ^ Оджеманн Г.А. (июнь 1983 г.). «Организация мозга для речи с точки зрения картирования электрической стимуляции». Поведенческие и мозговые науки . 6 (2): 189–206. дои : 10.1017/S0140525X00015491 . ISSN 1469-1825 . S2CID 143189089 .
- ^ Корнелиссен К., Лайне М., Ренвалл К., Сааринен Т., Мартин Н., Салмелин Р. (июнь 2004 г.). «Изучение новых названий новых объектов: корковые эффекты, измеренные с помощью магнитоэнцефалографии». Мозг и язык . 89 (3): 617–22. дои : 10.1016/j.bandl.2003.12.007 . ПМИД 15120553 . S2CID 32224334 .
- ^ Хартвигсен Г., Баумгертнер А., Прайс С.Дж., Кенке М., Ульмер С., Зибнер Х.Р. (сентябрь 2010 г.). «Фонологические решения требуют как левой, так и правой надмаргинальной извилины» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (38): 16494–9. Бибкод : 2010PNAS..10716494H . дои : 10.1073/pnas.1008121107 . ПМЦ 2944751 . ПМИД 20807747 .
- ^ Корнелиссен К., Лайне М., Таркиайнен А., Ярвенсиву Т., Мартин Н., Салмелин Р. (апрель 2003 г.). «Пластичность мозга взрослого человека, вызванная лечением аномии» . Журнал когнитивной нейронауки . 15 (3): 444–61. дои : 10.1162/089892903321593153 . ПМИД 12729495 . S2CID 1597939 .
- ^ Мечелли А., Крайнион Дж.Т., Ноппени Ю., О'Доэрти Дж., Эшбернер Дж., Фраковяк Р.С., Прайс СиДжей (октябрь 2004 г.). «Нейролингвистика: структурная пластичность двуязычного мозга». Природа . 431 (7010): 757. Бибкод : 2004Natur.431..757M . дои : 10.1038/431757a . hdl : 11858/00-001M-0000-0013-D79B-1 . ПМИД 15483594 . S2CID 4338340 .
- ^ Грин Д.В., Крайнион Дж., Прайс СиДжей (июль 2007 г.). «Изучение влияния межъязыковой лексики на структуры мозга с использованием воксельной морфометрии» . Двуязычие . 10 (2): 189–199. дои : 10.1017/S1366728907002933 . ПМК 2312335 . ПМИД 18418473 .
- ^ Уиллнесс С (08 января 2016 г.). «Оксфордский справочник по организационному климату и культуре. Бенджамин Шнайдер и Карен М. Барбера (ред.) Нью-Йорк, штат Нью-Йорк: Oxford University Press, 2014. ISBN 978-0-19-986071-5». Рецензии на книги. Британский журнал психологии . 107 (1): 201–202. дои : 10.1111/bjop.12170 .
- ^ Ли Х, Девлин Дж.Т., Шейкшафт К., Стюарт Л.Х., Бреннан А., Гленсман Дж., Питчер К., Крайнион Дж., Мечелли А., Фраковяк Р.С., Грин Д.В., Прайс СиДжей (январь 2007 г.). «Анатомические следы приобретения словарного запаса в мозге подростка» . Журнал неврологии . 27 (5): 1184–9. doi : 10.1523/JNEUROSCI.4442-06.2007 . ПМК 6673201 . ПМИД 17267574 .
- ^ Ричардсон Ф.М., Томас М.С., Филиппи Р., Харт Х., Прайс СиДжей (май 2010 г.). «Контрастное влияние словарного запаса на височную и теменную структуру мозга на протяжении всей жизни» . Журнал когнитивной нейронауки . 22 (5): 943–54. дои : 10.1162/jocn.2009.21238 . ПМЦ 2860571 . ПМИД 19366285 .
- ^ Джобард Дж., Кривелло Ф., Цурио-Мазойер Н. (октябрь 2003 г.). «Оценка теории двойного маршрута чтения: метаанализ 35 исследований нейровизуализации». НейроИмидж . 20 (2): 693–712. дои : 10.1016/s1053-8119(03)00343-4 . ПМИД 14568445 . S2CID 739665 .
- ^ Болджер DJ, Perfetti CA, Schneider W (май 2005 г.). «Пересмотр межкультурного воздействия на мозг: универсальные структуры плюс вариации системы письма» . Картирование человеческого мозга (на французском языке). 25 (1): 92–104. дои : 10.1002/hbm.20124 . ПМЦ 6871743 . ПМИД 15846818 .
- ^ Брамбати С.М., Огар Дж., Нойхаус Дж., Миллер Б.Л., Горно-Темпини М.Л. (июль 2009 г.). «Нарушения чтения при первичной прогрессирующей афазии: поведенческое и нейровизуализирующее исследование» . Нейропсихология . 47 (8–9): 1893–900. doi : 10.1016/j.neuropsychologia.2009.02.033 . ПМЦ 2734967 . ПМИД 19428421 .
- ^ Перейти обратно: а б Баддели А., Льюис В., Валлар Дж. (май 1984 г.). «Изучение артикуляционной петли» . Ежеквартальный журнал экспериментальной психологии Раздел А. 36 (2): 233–252. дои : 10.1080/14640748408402157 . S2CID 144313607 .
- ^ Перейти обратно: а б Коуэн Н. (февраль 2001 г.). «Магическое число 4 в кратковременной памяти: переосмысление умственных способностей» . Поведенческие и мозговые науки . 24 (1): 87–114, обсуждение 114–85. дои : 10.1017/S0140525X01003922 . ПМИД 11515286 .
- ^ Каплан Д., Рочон Э., Уотерс Г.С. (август 1992 г.). «Артикуляционные и фонологические детерминанты влияния длины слова в задачах с промежутком». Ежеквартальный журнал экспериментальной психологии. А. Экспериментальная психология человека . 45 (2): 177–92. дои : 10.1080/14640749208401323 . ПМИД 1410554 . S2CID 32594562 .
- ^ Уотерс Г.С., Рошон Э., Каплан Д. (февраль 1992 г.). «Роль высокого уровня планирования речи на репетиции: данные пациентов с апраксией речи». Журнал памяти и языка . 31 (1): 54–73. дои : 10.1016/0749-596x(92)90005-i .
- ^ Коэн Л., Башуд-Леви AC (сентябрь 1995 г.). «Роль выходного фонологического буфера в контроле синхронизации речи: отдельный пример». Кора; Журнал, посвященный изучению нервной системы и поведения . 31 (3): 469–86. дои : 10.1016/s0010-9452(13)80060-3 . ПМИД 8536476 . S2CID 4480375 .
- ^ Шаллис Т., Румиати Р.И., Задини А. (сентябрь 2000 г.). «Выборочное ухудшение фонологического выходного буфера». Когнитивная нейропсихология . 17 (6): 517–46. дои : 10.1080/02643290050110638 . ПМИД 20945193 . S2CID 14811413 .
- ^ Шу Х, Сюн Х, Хань З, Би Ю, Бай Икс (2005). «Избирательное нарушение фонологического выходного буфера: свидетельства китайского пациента» . Поведенческая неврология . 16 (2–3): 179–89. дои : 10.1155/2005/647871 . ПМЦ 5478832 . ПМИД 16410633 .
- ^ Оберауер К (2002). «Доступ к информации в рабочей памяти: исследование фокуса внимания». Журнал экспериментальной психологии: обучение, память и познание . 28 (3): 411–421. дои : 10.1037/0278-7393.28.3.411 . ПМИД 12018494 .
- ^ Ансворт Н., Энгл Р.В. (январь 2007 г.). «Природа индивидуальных различий в объеме рабочей памяти: активное поддержание первичной памяти и контролируемый поиск во вторичной памяти». Психологический обзор . 114 (1): 104–32. дои : 10.1037/0033-295x.114.1.104 . ПМИД 17227183 .
- ^ Барруйе П., Камос V (декабрь 2012 г.). «По прошествии времени» . Современные направления психологической науки . 21 (6): 413–419. дои : 10.1177/0963721412459513 . S2CID 145540189 .
- ^ Борнкессель-Шлезевски И., Шлезевски М., Смолл С.Л., Раушекер Дж.П. (март 2015 г.). «Нейробиологические корни языка при слухе у приматов: общие вычислительные свойства» . Тенденции в когнитивных науках . 19 (3): 142–50. дои : 10.1016/j.tics.2014.12.008 . ПМК 4348204 . ПМИД 25600585 .
- ^ Бухсбаум Б.Р., Д'Эспозито М. (май 2008 г.). «Поиск фонологического хранилища: от петли к свертке». Журнал когнитивной нейронауки . 20 (5): 762–78. дои : 10.1162/jocn.2008.20501 . ПМИД 18201133 . S2CID 17878480 .
- ^ де Соссюр, Фердинанд (1959) [Впервые опубликовано в 1916 году]. Курс общего языкознания (PDF) . Нью-Йорк: Философская библиотека. ISBN 9780231157278 . Архивировано из оригинала (PDF) 14 апреля 2020 г. Проверено 16 июня 2020 г.
- ^ Смит, Нил (2002). Хомский: идеи и идеалы (2-е изд.). Издательство Кембриджского университета. ISBN 0-521-47517-1 .
- ^ Лакофф, Джордж (1990). «Гипотеза инвариантности: основаны ли абстрактные рассуждения на схемах изображений?». Когнитивная лингвистика . 1 (1): 39–74. дои : 10.1515/cogl.1990.1.1.39 . S2CID 144380802 .
- ^ Сон, Джэ Юнг (2012). Порядок слов . Издательство Кембриджского университета. ISBN 9781139033930 .
- ^ Кристиансен, Мортен Х.; Чейтер, Ник (2008). «Язык, сформированный мозгом» (PDF) . Поведенческие и мозговые науки . 31 (5): 489–558. дои : 10.1017/S0140525X08004998 . ПМИД 18826669 . Проверено 22 декабря 2020 г.
- ^ Блэкмор, Сьюзен (2008). «Мемы формируют мозг, формируют мемы» . Поведенческие и мозговые науки . 31 (5): 513. doi : 10.1017/S0140525X08005037 . Проверено 22 декабря 2020 г.
- ^ Клюендер, Р.; Кутас, М. (1993). «Подчиненность как феномен обработки» (PDF) . Язык и когнитивные процессы . 8 (4): 573–633. дои : 10.1080/01690969308407588 . Проверено 28 февраля 2020 г.
- ^ Баркли, К.; Клюендер, Р.; Кутас, М. (2015). «Обработка ссылок в человеческом мозге: исследование потенциала, связанного с событиями (ERP)» (PDF) . Исследования мозга . 1629 : 143–159. дои : 10.1016/j.brainres.2015.09.017 . ПМИД 26456801 . S2CID 17053154 . Проверено 28 февраля 2020 г.
- ^ Вэй, Сюэху; Адамсон, Хелин; Швендеманн, Матиас; Гуча, Томас; Фридеричи, Анжела Д.; Анвандер, Альфред (19 февраля 2023 г.). «Родные языковые различия в структурном коннектоме человеческого мозга» . НейроИмидж . 270 (270): 119955. doi : 10.1016/j.neuroimage.2023.119955 . ПМИД 36805092 .
- ^ Коидзуми, Масатоши; Ясуги, Ёсихо; Тамаока, Кацуо; Кияма, Сатико; Ким, Чонхо; Айсивинац Сиан, Хуан Эстебан; Гарсиа Мацар, Педро Оскар (сентябрь 2014 г.). «О (не) универсальности предпочтения субъектно-объектного порядка слов при понимании предложения: исследование обработки предложений в Какчикель Майя» . Язык . 90 (3): 722–736. дои : 10.1353/lan.2014.0068 . JSTOR 24672044 . S2CID 146776347 . Проверено 15 мая 2023 г.
- ^ Миллер Л.М., Recanzone GH (апрель 2009 г.). «Популяции слуховых кортикальных нейронов могут точно кодировать акустическое пространство в зависимости от интенсивности стимула» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (14): 5931–5. Бибкод : 2009PNAS..106.5931M . дои : 10.1073/pnas.0901023106 . ПМК 2667094 . ПМИД 19321750 .
- ^ Тиан Б., Резер Д., Дарем А., Кустов А., Раушекер Дж. П. (апрель 2001 г.). «Функциональная специализация слуховой коры макаки-резус». Наука . 292 (5515): 290–3. Бибкод : 2001Sci...292..290T . дои : 10.1126/science.1058911 . ПМИД 11303104 . S2CID 32846215 .
- ^ Ален С., Арнотт С.Р., Хевенор С., Грэм С., Грейди С.Л. (октябрь 2001 г.). « Что» и «где» в слуховой системе человека» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (21): 12301–6. Бибкод : 2001PNAS...9812301A . дои : 10.1073/pnas.211209098 . ПМК 59809 . ПМИД 11572938 .
- ^ Де Сантис Л., Кларк С., Мюррей М.М. (январь 2007 г.). «Автоматическая и внутренняя слуховая обработка слов «что» и «где» у людей, выявленная с помощью электрической нейровизуализации» . Кора головного мозга . 17 (1): 9–17. дои : 10.1093/cercor/bhj119 . ПМИД 16421326 .
- ^ Барретт DJ, Hall DA (август 2006 г.). «Предпочтения реакции на «что» и «где» в непервичной слуховой коре человека». НейроИмидж . 32 (2): 968–77. doi : 10.1016/j.neuroimage.2006.03.050 . ПМИД 16733092 . S2CID 19988467 .
- ^ Линден Дж. Ф., Грюневальд А., Андерсен Р. А. (июль 1999 г.). «Ответы на слуховые стимулы в латеральной интратеменной области макака. II. Поведенческая модуляция». Журнал нейрофизиологии . 82 (1): 343–58. дои : 10.1152/jn.1999.82.1.343 . ПМИД 10400963 . S2CID 5317446 .
- ^ Маццони П., Брейсуэлл Р.М., Бараш С., Андерсен Р.А. (март 1996 г.). «Пространственно настроенные слуховые реакции в области LIP макак, выполняющих задержанные саккады памяти на акустические цели». Журнал нейрофизиологии . 75 (3): 1233–41. дои : 10.1152/jn.1996.75.3.1233 . ПМИД 8867131 .
- ^ Лашо Дж.П., Джерби К., Бертран О., Минотти Л., Хоффманн Д., Шендорф Б., Кахане П. (октябрь 2007 г.). «Проект функционального картирования в реальном времени с помощью внутричерепных записей человека» . ПЛОС ОДИН . 2 (10): е1094. Бибкод : 2007PLoSO...2.1094L . дои : 10.1371/journal.pone.0001094 . ПМК 2040217 . ПМИД 17971857 .
- ^ Жардри Р., Уффлин-Дебарж В., Делион П., Пруво Дж.П., Томас П., Пинс Д. (апрель 2012 г.). «Оценка реакции плода на материнскую речь с использованием метода неинвазивной функциональной визуализации мозга». Международный журнал нейробиологии развития . 30 (2): 159–61. дои : 10.1016/j.ijdevneu.2011.11.002 . ПМИД 22123457 . S2CID 2603226 .
- ^ Полива О (20 сентября 2017 г.). «Откуда к чему: нейроанатомически обоснованная эволюционная модель возникновения речи у человека» . F1000Исследования . 4 : 67. doi : 10.12688/f1000research.6175.3 . ПМК 5600004 . ПМИД 28928931 .
- ^ Полива О (30 июня 2016 г.). «От мимикрии к языку: нейроанатомическая эволюционная модель возникновения голосового языка» . Границы в неврологии . 10 : 307. дои : 10.3389/fnins.2016.00307 . ПМЦ 4928493 . ПМИД 27445676 .
- ^ Перейти обратно: а б с д и Сури, Сана (25 июля 2014 г.). «Чему язык жестов учит нас о мозге» . Разговор . Проверено 7 октября 2019 г.
- ^ Перейти обратно: а б с Научный американец. (2002). Язык жестов в мозгу. [Брошюра]. Получено с http://lcn.salk.edu/Brochure/SciAM%20ASL.pdf.
- ^ Перейти обратно: а б с д и ж г час я дж к л Нортон Э.С., Ковельман И., Петитто Л.А. (март 2007 г.). «Существуют ли отдельные нейронные системы для правописания? Новое понимание роли правил и памяти в правописании на основе функциональной магнитно-резонансной томографии» . Разум, мозг и образование . 1 (1): 48–59. дои : 10.1111/j.1751-228X.2007.00005.x . ПМК 2790202 . ПМИД 20011680 .
- ^ Перейти обратно: а б с д и Трейман Р., Кесслер Б. (2007). Системы письма и развитие правописания . Blackwell Publishing Ltd., стр. 120–134. дои : 10.1002/9780470757642.ch7 . ISBN 978-0-470-75764-2 .
{{cite book}}:|work=игнорируется ( помогите )