Тропизм эндотелиальных клеток
Тропизм эндотелиальных клеток или эндотелиотропизм — это тип тропизма ткани или тропизма хозяина , который характеризует способность патогена распознавать и инфицировать эндотелиальную клетку. Патогены, такие как вирусы, могут поражать определенный тип ткани или несколько типов тканей. Как и другие клетки, эндотелиальная клетка обладает рядом особенностей, которые поддерживают продуктивную вирусную инфекцию клетки, включая рецепторы клеточной поверхности , иммунные реакции и другие факторы вирулентности . [ 1 ] Эндотелиальные клетки встречаются в различных типах тканей, таких как капилляры , вены и артерии человеческого тела. Поскольку эндотелиальные клетки выстилают эти кровеносные сосуды и критические сети, которые расширяют доступ к различным системам органов человека, проникновение вируса в эти клетки может оказаться пагубным для распространения вируса по системе хозяина и повлиять на клиническое течение заболевания. Понимание механизмов того, как вирусы прикрепляются, проникают и контролируют функции эндотелия и реакцию хозяина, дает информацию о понимании инфекционных заболеваний и медицинских контрмерах .
Клеточные особенности и механизмы
[ редактировать ]Существует множество особенностей эндотелиальных клеток, которые влияют на тропизм клеток и, в конечном итоге, способствуют активации и дисфункции эндотелиальных клеток , а также продолжению жизненного цикла вируса .
Рецепторы клеточной поверхности
[ редактировать ]
Вирусные патогены используют рецепторы клеточной поверхности, которые распространены повсеместно и могут распознавать множество разнообразных лигандов для прикрепления и, в конечном итоге, проникновения в клетку. Эти лиганды состоят не только из эндогенных белков, но также из бактериальных и вирусных продуктов. Как только вирус закрепляется на поверхности клетки, поглощение вируса обычно происходит с использованием механизмов хозяина, таких как эндоцитоз . [ 2 ] [ 3 ] Одним из методов захвата вируса является клатрин-опосредованный эндоцитоз (CME). [ 4 ] Рецепторы клеточной поверхности обеспечивают связывающий карман для прикрепления и проникновения в клетку и, следовательно, влияют на восприимчивость клетки к инфекции. Кроме того, плотность рецепторов на поверхности эндотелиальной клетки также влияет на эффективность проникновения вируса в клетку-хозяина. Например, более низкая плотность рецепторов на клеточной поверхности может сделать эндотелиальную клетку менее восприимчивой к вирусной инфекции, чем эндотелиальную клетку с более высокой плотностью рецепторов на клеточной поверхности. Эндотелий содержит множество рецепторов клеточной поверхности, связанных с такими функциями, как прикрепление и транспортировка иммунных клеток, свертывание крови , расширение сосудов и проницаемость барьера. [ 5 ] Учитывая эти жизненно важные функции, взаимодействие вируса с этими рецепторами позволяет понять симптомы, которые присутствуют во время вирусного патогенеза, такие как воспаление , повышенная проницаемость сосудов и тромбоз .

Факторы транскрипции и репликация вируса
[ редактировать ]После проникновения в клетку этим внутриклеточным паразитам требуются факторы в клетке-хозяине для поддержки репликации вируса и высвобождения вирионов-потомков. В частности, факторы-хозяева включают белки , такие как факторы транскрипции и полимеразы , которые помогают в репликации вирусного генома. [ 7 ] Следовательно, единственное проникновение в живого хозяина не обязательно приводит к размножению вирусного потомства, поскольку клетка может не содержать критических факторов транскрипции или полимераз для репликации вируса. Более того, внутри вирусного генома существуют не только инструкции по синтезу вирусных белков, но и другие факторы вирулентности, такие как гены , клеточные структуры и другие регуляторные процессы, которые позволяют патогену контролировать противовирусные реакции хозяина. [ 8 ] хозяина Эти факторы вирулентности могут противодействовать защитным механизмам хозяина, которые пытаются устранить инфекцию через иммунную систему .
Защитные механизмы хозяина
[ редактировать ]Эндотелиальные клетки также обладают внутренними противовирусными реакциями, которые используют иммунную систему хозяина для борьбы с инфекцией или ограничения репликации вируса. [ 9 ] [ 10 ] В ответ на продукцию вируса в клетке клетка-хозяин может высвободить белок, такой как цитокин, подобный интерферону (ИФН) , который будет сигнализировать об иммунном ответе. ИФН «мешает» репликации вируса, передавая сигнал другим клеткам нашей иммунной системы, чтобы остановить инфекцию. Другие клеточные механизмы также находятся на различных субклеточных уровнях. В частности, существуют рецепторы распознавания клеточных образов, такие как TLR7 и TLR8 (обнаружение РНК) и TLR9 (обнаружение ДНК). [ 11 ] Эти toll-подобные рецепторы могут различать наличие вирусных нуклеиновых кислот в клетке-хозяине, а также запускать иммунный ответ, чтобы пометить клетку и попытаться устранить патоген. Сочетание этих механизмов, которые поддерживают успешное проникновение вируса, репликацию вируса и блокирование иммунного ответа хозяина, способствуют продуктивному заражению и репликации вируса.
Примеры и влияние на вирусный патогенез
[ редактировать ]Коронавирусы
[ редактировать ]
SARS-CoV-2 — это вирус, вызывающий заболевание COVID-19 и поражающий различные типы клеток, но также демонстрирующий поражение сосудов многих органов. В тяжелых случаях SARS-CoV-2 может вызвать эндотелиальную дисфункцию или повреждение. Эти вызванные вирусом реакции эндотелия могут привести к тромбозу, застою и микроангиопатии . [ 12 ] Рецепторы клеточной поверхности, связанные с проникновением вируса, включают ACE2 и корецептор TMPRSS2 . TMPRSS2 необходим для расщепления белка-шипа для слияния вируса с клеточной мембраной. Однако недавнее исследование показало, что низкая экспрессия ACE2 в эндотелиальных клетках связана с плохой способностью к размножению вируса из-за отсутствия точек входа на поверхности клетки. [ 13 ]
Флавивирусы
[ редактировать ]Денге является примером флавивируса , переносимого комарами , который вызывает лихорадку Денге . Хотя эндотелиальные клетки не являются основными типами клеток-мишеней лихорадки Денге, вирус связывается с различными рецепторами клеточной поверхности на эндотелиальных клетках с особенно продуктивным инфицированием через рецепторы клеточной поверхности, содержащие гепарансульфат . [ 14 ] Было показано, что инфицирование эндотелия через эти рецепторы нарушает критические иммунные реакции и изменяет проницаемость капилляров, что, в свою очередь, поддерживает клиническое течение заболевания. [ 15 ]
Филовирусы
[ редактировать ]Эбола — один из вирусов геморрагической лихорадки, вызывающий болезнь, вызванную вирусом Эбола (БВВЭ) . Анализ образцов человека, не перенесших заболевание, показал, что эндотелий значительно изменен по сравнению со здоровым состоянием. [ 16 ] [ 17 ] Другие изменения гомеостаза включают широко распространенную экспрессию вирусных антигенов в эндотелиальных клетках. [ 18 ] вируса Было установлено, что гликопротеин , который служит «ключом» вируса к клетке, серьезно повреждает эндотелий. [ 16 ] Например, печень часто считается зоной поражения при инфекции. Синусоидальные эндотелиальные клетки печени (LSEC) экспрессируют различные рецепторы-поглотители , включая FcγRIIb2 и рецептор маннозы , которые имеют решающее значение для устранения молекул отходов в печени, но также поглощают лиганды по пути CME. [ 19 ] Помимо поддержки проникновения вируса, взаимодействие с этими рецепторами также может препятствовать выведению фармацевтических препаратов, применяемых для смягчения инфекции.
Ортомиксовирусиды
[ редактировать ]Грипп A H1N1 — это подтип вируса гриппа , который поражает эндотелиальные клетки дыхательной системы, например, легких, и инфицирует их. Вирус также может поражать эпителий слизистых оболочек этих систем органов. [ 20 ] Вирусные частицы имеют тенденцию выходить из просвета эндотелия, что приводит к обнаружению вирусных антигенов в крови и лимфатических эндотелиальных клетках. Однако по мере распространения этого вируса он будет поражать эндотелиальные клетки легких, а не, например, головного мозга.
Приложения
[ редактировать ]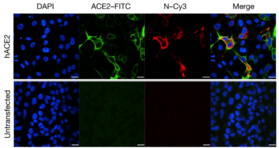
Технологии обучения
[ редактировать ]
В зависимости от уровня биобезопасности (BSL), также известного как уровень патогена или защиты, существуют различные уровни биологической защиты и разрешения, необходимые для изучения патогена; этот уровень защиты влияет на то, как и где изучается возбудитель. [ 21 ] Хотя в этих обзорах основное внимание уделяется тропизму эндотелиальных клеток, эти методы также широко применимы к различным методам вирусологии . Эти резюме не содержат полного списка, но представляют собой общие платформы для изучения новых инфекционных заболеваний.
Подходы in vitro
[ редактировать ]Иммортализованные клетки представляют собой возобновляемый ресурс для изучения различных патогенов. Характеристика эндотелиального тропизма позволяет исследователям модифицировать любую клетку, чтобы отобразить рецептор, с которым взаимодействует гликопротеин вируса, для прикрепления к клетке. Однако эти 2D-клеточные культуры не обязательно предназначены для имитации распространения вируса или реакций хозяина in vivo . Эти форматы биоанализов позволяют исследовать вирус и потенциально идентифицировать рецепторы клеточной поверхности или другие факторы, участвующие в тропизме клеток. Обычно методы молекулярной биологии, такие как иммунофлуоресценция или иммуногистохимия , позволяют исследователям визуализировать, где рецептор присутствует в клетке. И наоборот, использование суррогатного или псевдотипированного вируса также является методом понимания клеточного тропизма. Короче говоря, эти подходы обычно используют другой и хорошо охарактеризованный патоген, такой как вирус везикулярного стоматита (ВВС) , и модифицируют его так, чтобы он отображал гликопротеин другого представляющего интерес вируса. [ 22 ] Поскольку гликопротеин служит «ключом» к клетке, этот метод позволяет изучать проникновение в клетку независимо от других процессов жизненного цикла вируса. Дальнейшее выращивание или серийное пассирование этого рекомбинантного вируса может продемонстрировать, как вирус развивается или мутирует для поддержания эффективности заражения.
Модели заражения in vivo
[ редактировать ]
Приматы, не относящиеся к человеку, такие как макаки-резусы, служат «золотым стандартом» для животных моделей многих патогенов BSL4, когда биологический феномен не может быть изучен на других видах. [ 23 ] Поскольку многие инфекционные заболевания носят зоонозный характер, моделирование этих заболеваний у видов макак, которые имеют некоторое биологическое сходство с людьми, дает представление о понимании болезней в обстоятельствах, когда вирус плохо изучен, а варианты лечения ограничены или отсутствуют. Показания этих моделей можно оценить, например, с помощью образцов тканей или образцов крови. Однако эти модели инфекции in vivo, такие как модели грызунов и приматов, не являющихся человеком, представляют собой этические проблемы и недостатки, поскольку они включают лабораторное заключение животного и введение ему инфекции. [ 24 ] Появление передовых моделей in vivo, в том числе гуманизированных или трансгенных грызунов, обеспечивает альтернативу серии макак, но также вызывает опасения, повторяют ли эти модели физиологию человека или предсказывают человеческие реакции на заболевание или лечение. Эти модели включают генетическую модификацию и/или трансплантацию человеческой ткани в модель грызуна. [ 25 ] В сочетании с in vitro клеточными анализами эти модели in vivo имеют решающее значение для проверки эффективности терапии во время открытия и разработки лекарств.
Открытие и разработка лекарств
[ редактировать ]Тропизм эндотелиальных клеток определяет медицинские контрмеры в ответ на возникающие инфекционные заболевания. таких терапевтических средств, как низкомолекулярные соединения и вакцины Эти медицинские контрмеры включают разработку .
Противовирусные препараты и другие низкомолекулярные препараты
[ редактировать ]Противовирусные препараты — это терапевтические средства, которые помогают организму человека устранить инфекцию, смягчить симптомы инфекции и/или уменьшить клиническое течение заболевания. Понимание тропизма эндотелиальных клеток используется при открытии мишеней противовирусных препаратов. Многие механизмы действия этих терапевтических средств в первую очередь нацелены на жизненный цикл вируса. [ 26 ] Эти препараты выпускаются в форме низкомолекулярных соединений или других биотерапевтических средств (например, терапии моноклональными антителами ). клеточном При высокопроизводительном скрининге лекарств тропизм клеток является важным фактором при выборе типа клеток. Тип клеток в этих анализах должен отображать целевой рецептор, чтобы репрезентативно подтвердить предполагаемый механизм действия препарата и определить его эффективность , безопасность и эффективность in vitro . [ 27 ] Более того, другие аспекты тропизма эндотелиальных клеток поддаются терапевтическим подходам. Эти аспекты включают разнообразные механизмы того, как эндотелиальные клетки обнаруживают вирусы и реагируют на инфекцию. Например, эндотелиальный барьер служит одновременно защитным барьером и медиатором иммунных ответов против инородных тел. Однако эндотелиальный барьер подвергается повреждению в результате вирусной инфекции. Терапия, которая усиливает или восстанавливает целостность эндотелиального барьера после его повреждения, рассматривается как потенциальная мишень для новых инфекционных заболеваний, таких как COVID-19, Эбола, лихорадка Денге и других. [ 1 ] [ 28 ] В целом, исследование тропизма эндотелиальных клеток может дать представление о соответствующих терапевтических вмешательствах.
| Группа | Drug(s) | Механизм действия | Ссылки |
|---|---|---|---|
| Ингибиторы проникновения вируса | Маравирок | Блокирует взаимодействие рецепторов, эндоцитоз/макропиноцитоз, прикрепление, слияние или передачу сигналов, связанных с поглощением вируса. | [ 26 ] [ 29 ] [ 30 ] |
| Ингибиторы синтеза вирусного белка | Лопинавир/ритонавир | Подавляет или замедляет репликацию вируса, нарушая процессы, связанные с трансляцией или генерацией белка. | [ 31 ] |
| Ингибиторы вирусной полимеразы | Молнупиравир | Вмешивается в регуляцию транскрипции вирусных белков во время репликации вируса. | [ 32 ] |
| Иммуномодуляторы | Нитазоксанид | Вмешиваться и противодействовать регулируемым хозяином путям во время репликации вируса (например, путям IFN и механизму восприятия вирусной РНК) | [ 26 ] [ 33 ] |
Вакцина
[ редактировать ]Вакцины – это терапевтические средства, которые являются профилактической мерой инфекционных заболеваний. организма Эти методы лечения обеспечивают адаптивный иммунитет к определенному патогену. По сути, вакцины обеспечивают защиту пациентов, вызывая иммунный ответ, в результате которого у них вырабатываются антитела, которые помогут защититься от вторгающегося патогена. Разработка, производство и глобальное распространение этих вакцин необходимы для предотвращения, контроля и искоренения потенциально пандемических патогенов. В частности, в технологиях вакцин, основанных на культивировании клеток, используются клеточные линии, обладающие широким диапазоном вирусного тропизма, для адаптации штаммов вируса, используемых при разработке вакцин, к новым клеткам. [ 34 ] Это применение клеточного тропизма оценивает различные пути проникновения вируса и рецепторы хозяина для достижения этой цели. Более того, аспекты активации и дисфункции эндотелиальных клеток становятся важными показателями во время разработки вакцин, поскольку они являются отличительными чертами многих клинических проявлений инфекционных заболеваний. Одним из наиболее многообещающих кандидатов на вакцину против Эболы является компании Merck рекомбинантная вакцина VSV-EBOV Ervebo . в конце Вакцина имела решающее значение во время вспышки Эболы в Гвинее 2014–2015 гг . Эффективность Эрвебо была доказана на приматах, а затем и в Гвинее в ходе санкционированного исследования эффективности на людях, которое показало, что Эрвебо также обладает высокой защитной способностью у людей. В вакцине ВСВ используется в качестве суррогата для отображения гликопротеина Эболы. [ 35 ] ВВС не вызывает заболеваний у людей, что делает его полезным стержнем для хранения важного белка заирского вируса Эбола . При введении вакцины рекомбинантный VSV вводит функциональный гликопротеин вируса Эбола, который взаимодействует с барьером эндотелиальных клеток и вызывает быстрый иммунный ответ, не вызывая заболевания у пациентов. Таким образом, разработка и масштабирование вакцин учитывают важные аспекты тропизма эндотелиальных клеток.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Фосс, Джоанна Хол; Харальдсен, Гутторм; Фальк, Кнут; Эдельманн, Рейдунн (2021). «Эндотелиальные клетки при возникающих вирусных инфекциях» . Границы сердечно-сосудистой медицины . 8 : 95. дои : 10.3389/fcvm.2021.619690 . ISSN 2297-055X . ПМЦ 7943456 . ПМИД 33718448 .
- ^ МакМахон, Харви Т.; Букро, Эммануэль (август 2011 г.). «Молекулярный механизм и физиологические функции клатрин-опосредованного эндоцитоза» . Nature Reviews Молекулярно-клеточная биология . 12 (8): 517–533. дои : 10.1038/nrm3151 . ISSN 1471-0080 . ПМИД 21779028 . S2CID 15235357 .
- ^ Каксонен, Марко; Ру, Орельен (май 2018 г.). «Механизмы клатрин-опосредованного эндоцитоза» . Nature Reviews Молекулярно-клеточная биология . 19 (5): 313–326. дои : 10.1038/номер.2017.132 . ISSN 1471-0080 . ПМИД 29410531 . S2CID 4380108 .
- ^ Коссар, Паскаль; Хелениус, Ари (01 августа 2014 г.). «Эндоцитоз вирусов и бактерий» . Перспективы Колд-Спринг-Харбор в биологии . 6 (8): а016972. doi : 10.1101/cshperspect.a016972 . ISSN 1943-0264 . ПМК 4107984 . ПМИД 25085912 .
- ^ Юань, Сара Ю.; Строгость, Роберт Р. (2010). Сигнальные механизмы регуляции эндотелиальной проницаемости . Морган и Клейпул Науки о жизни.
- ^ Бернар, Изабель; Лимонта, Дэниел; Махал, Лара; Хобман, Том (26 декабря 2020 г.). «Инфекция и нарушение регуляции эндотелия, вызванные SARS-CoV-2: данные и предостережения относительно COVID-19» . Вирусы . 13 (1): 29. дои : 10.3390/v13010029 . ПМЦ 7823949 . ПМИД 33375371 .
- ^ Феннер, Фрэнк; Бахманн, Питер А.; Гиббс, Э. Пол Дж.; Мерфи, Фредерик А.; Стаддерт, Майкл Дж.; Уайт, Дэвид О. (1987), Феннер, Фрэнк; Бахманн, Питер А.; Гиббс, Э. Пол Дж.; Мерфи, Фредерик А. (ред.), «Глава 4 – Репликация вируса» , Ветеринарная вирусология , Academic Press, стр. 55–88, ISBN 978-0122530555
- ^ Бэгген, Джим; Ванстрилс, Элс; Янсен, Сандер; Далеманс, Дирк (октябрь 2021 г.). «Клеточные факторы-хозяева инфекции SARS-CoV-2» . Природная микробиология . 6 (10): 1219–1232. дои : 10.1038/s41564-021-00958-0 . ISSN 2058-5276 . ПМИД 34471255 . S2CID 237387636 .
- ^ Макфадден, Грант; Мохамед, Мохамед Р.; Рахман, Масмудур М.; Барти, Эрик (сентябрь 2009 г.). «Цитокиновые детерминанты вирусного тропизма» . Обзоры природы Иммунология . 9 (9): 645–655. дои : 10.1038/nri2623 . ISSN 1474-1741 . ПМЦ 4373421 . ПМИД 19696766 .
- ^ Маждул, Салиха; Комптон, Алекс А. (13 октября 2021 г.). «Уроки самообороны: подавление проникновения вируса за счет внутреннего иммунитета» . Обзоры природы Иммунология . 22 (6): 339–352. дои : 10.1038/s41577-021-00626-8 . ISSN 1474-1741 . ПМК 8511856 . ПМИД 34646033 .
- ^ Пандей, Сурья; Каваи, Таро; Акира, Шизуо (01 января 2015 г.). «Микробное зондирование с помощью Toll-подобных рецепторов и внутриклеточных сенсоров нуклеиновых кислот» . Перспективы Колд-Спринг-Харбор в биологии . 7 (1): а016246. doi : 10.1101/cshperspect.a016246 . ISSN 1943-0264 . ПМЦ 4292165 . ПМИД 25301932 .
- ^ Джин, Юэфэй; Цзи, Ванцюань; Ян, Хайян; Чен, Шуайинь; Чжан, Вэйго; Дуань, Гуанкай (24 декабря 2020 г.). «Эндотелиальная активация и дисфункция при COVID-19: от основных механизмов к потенциальным терапевтическим подходам» . Сигнальная трансдукция и таргетная терапия . 5 (1): 293. дои : 10.1038/s41392-020-00454-7 . ISSN 2059-3635 . ПМЦ 7758411 . ПМИД 33361764 .
- ^ Шиммель, Лилиан; Чу, Кенг Йи; Стокс, Клаудия Дж.; Йорданов, Теодор Э.; Эссебье, Патрисия; Куласингхе, Арута; Монкман, Джеймс; Миджиоларо, Анна Флавия; Купер, Кэролайн; Норонья, Люсия; Шредер, Кейт (2021). «Эндотелиальные клетки не заражаются SARS-CoV-2 продуктивно» . Клиническая и трансляционная иммунология . 10 (10):е1 дои : 10.1002/cti2.1350 . ISSN 2050-0068 . ПМЦ 8542944 . ПМИД 34721846 .
- ^ Нойфельдт, Кристофер Дж.; Кортезе, Мирко; Акоста, Элиана Г.; Бартеншлагер, Ральф (март 2018 г.). «Перестройка сотовых сетей членами семейства Flaviviridae» . Обзоры природы Микробиология . 16 (3): 125–142. дои : 10.1038/nrmicro.2017.170 . ISSN 1740-1534 . ПМК 7097628 . ПМИД 29430005 .
- ^ Басу, Атану; Чатурведи, Умеш К. (август 2008 г.). «Сосудистый эндотелий: поле битвы вирусов денге» . FEMS Иммунология и медицинская микробиология . 53 (3): 287–299. дои : 10.1111/j.1574-695x.2008.00420.x . ISSN 0928-8244 . ПМК 7110366 . ПМИД 18522648 .
- ^ Перейти обратно: а б Фаласка, Л.; Аграти, К.; Петросилло, Н.; Ди Каро, А.; Капобьянки, MR; Ипполито, Г.; Пьячентини, М. (август 2015 г.). «Молекулярные механизмы патогенеза вируса Эбола: фокус на гибели клеток» . Смерть клеток и дифференциация . 22 (8): 1250–1259. дои : 10.1038/cdd.2015.67 . ISSN 1476-5403 . ПМЦ 4495366 . ПМИД 26024394 .
- ^ Валь-Йенсен, Виктория М.; Афанасьева Татьяна Александровна; Зеебах, Йохен; Стрёрер, Юте; Фельдманн, Хайнц; Шниттлер, Ханс-Иоахим (август 2005 г.). «Влияние гликопротеинов вируса Эбола на активацию эндотелиальных клеток и барьерную функцию» . Журнал вирусологии . 79 (16): 10442–10450. doi : 10.1128/JVI.79.16.10442-10450.2005 . ISSN 0022-538X . ПМЦ 1182673 . ПМИД 16051836 .
- ^ Джайн, Сахил; Хайбуллина Светлана Ф.; Баранвал, Манодж (17 октября 2020 г.). «Иммунологический взгляд на инфекцию, вызванную вирусом Эбола, и различные лечебные меры, принимаемые для борьбы с этой болезнью» . Патогены (Базель, Швейцария) . 9 (10): Е850. doi : 10.3390/pathogens9100850 . ISSN 2076-0817 . ПМЦ 7603231 . ПМИД 33080902 .
- ^ Бхандари, Сабин; Ларсен, Анетт Кристин; МакКорт, Питер; Смедсрёд, Борд; Соренсен, Карен Кристин (2021). «Функция мусорщика синусоидальных эндотелиальных клеток печени в здоровом состоянии и при заболеваниях» . Границы в физиологии . 12 : 1711. doi : 10.3389/fphys.2021.757469 . ISSN 1664-042X . ПМЦ 8542980 . ПМИД 34707514 .
- ^ Шорт, Кирсти Р.; Вельдхус Кроэзе, Эдвин Дж.Б.; Реперант, Лесли А.; Ричард, Матильда; Куикен, Тейс (2014). «Вирус гриппа и эндотелиальные клетки: видоспецифические взаимоотношения» . Границы микробиологии . 5 : 653. дои : 10.3389/fmicb.2014.00653 . ISSN 1664-302X . ПМЦ 4251441 . ПМИД 25520707 .
- ^ Байот, Марлон Л.; Кинг, Кевин С. (2021 г.), «Уровни биологической опасности» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 30570972 , получено 1 ноября 2021 г.
- ^ Джоглекар, Алок В.; Сандовал, Салемиз (декабрь 2017 г.). «Псевдотипированные лентивирусные векторы: один вектор, множество обличий» . Методы генной терапии человека . 28 (6): 291–301. дои : 10.1089/hgtb.2017.084 . ISSN 1946-6536 . ПМИД 28870117 .
- ^ Беннетт, Ричард С.; Лог, Джеймс; Лю, Дэвид X.; Ридер, Ребекка Дж.; Яноско, Кристина Б.; Перри, Донна Л.; Купер, Тимоти К.; Байрам, Рассел; Рэгланд, Дэнни; Сент-Клер, Мариса; Адамс, Рики (14 июля 2020 г.). «Прогрессирование заболевания, вызванного вирусом Эбола Киквит, на модели животного-резус» . Вирусы . 12 (7): 753. дои : 10.3390/v12070753 . ISSN 1999-4915 . ПМЦ 7411891 . ПМИД 32674252 .
- ^ Карвалью, Констанца; Гаспар, Огаста; Найт, Эндрю; Висенте, Луис (29 декабря 2018 г.). «Этические и научные ошибки, связанные с лабораторными исследованиями на приматах, не являющихся людьми, и возможные решения» . Животные . 9 (1): 12. дои : 10.3390/ani9010012 . ISSN 2076-2615 . ПМК 6356609 . ПМИД 30597951 .
- ^ Масеманн, Дёрте; Людвиг, Стефан; Бёргелинг, Ивонн (январь 2020 г.). «Достижения в области моделей трансгенных мышей для изучения инфекций, вызываемых патогенными вирусами человека» . Международный журнал молекулярных наук . 21 (23): 9289. doi : 10.3390/ijms21239289 . ПМЦ 7730764 . ПМИД 33291453 .
- ^ Перейти обратно: а б с Каусар, Шамайла; Саид Хан, Фахад; Исхак Муджиб Ур Рехман, Мухаммед; Акрам, Мухаммед; Риаз, Мухаммед; Расул, Гулам; Хамид Хан, Абдул; Салим, Икра; Шамим, Саба; Малик, Ариф (01 января 2021 г.). «Обзор: Механизм действия противовирусных препаратов» . Международный журнал иммунопатологии и фармакологии . 35 : 20587384211002621. дои : 10.1177/20587384211002621 . ISSN 2058-7384 . ПМЦ 7975490 . ПМИД 33726557 .
- ^ Шиманский, Павел; Маркович, Магдалена; Микичук-Оласик, Эльжбета (29 декабря 2011 г.). «Адаптация высокопроизводительного скрининга при поиске лекарств — токсикологические скрининговые тесты» . Международный журнал молекулярных наук . 13 (1): 427–452. дои : 10.3390/ijms13010427 . ISSN 1422-0067 . ПМК 3269696 . ПМИД 22312262 .
- ^ Макоу, Эрих Р.; Горбунова Елена Евгеньевна; Гавриловская, Ирина Н. (2015). «Дисфункция эндотелиальных клеток при вирусных кровоизлияниях и отеках» . Границы микробиологии . 5 : 733. дои : 10.3389/fmicb.2014.00733 . ISSN 1664-302X . ПМК 4283606 . ПМИД 25601858 .
- ^ Чжоу, Янчен; Симмонс, Грэм (октябрь 2012 г.). «Разработка новых ингибиторов проникновения, нацеленных на новые вирусы» . Экспертный обзор противоинфекционной терапии . 10 (10): 1129–1138. дои : 10.1586/eri.12.104 . ISSN 1478-7210 . ПМЦ 3587779 . ПМИД 23199399 .
- ^ Маркхэм, Энтони (01 января 2021 г.). «REGN-EB3: Первое одобрение» . Наркотики . 81 (1): 175–178. дои : 10.1007/s40265-020-01452-3 . ISSN 1179-1950 . ПМЦ 7799152 . ПМИД 33432551 .
- ^ Ли, Бохэ Рэйчел; Паинг, Мэй Хнин; Шарма-Валия, Нилам (2021). «Циклопентеноновые простагландины: биологически активные липидные медиаторы, воздействующие на воспаление» . Границы в физиологии . 12 : 801. дои : 10.3389/fphys.2021.640374 . ISSN 1664-042X . ПМЦ 8320392 . ПМИД 34335286 .
- ^ Сингх, Авадеш Кумар; Сингх, Акрити; Сингх, Риту; Мисра, Ануп (01 ноября 2021 г.). «Молнупиравир при COVID-19: систематический обзор литературы» . Диабет и метаболический синдром: клинические исследования и обзоры . 15 (6): 102329. doi : 10.1016/j.dsx.2021.102329 . ISSN 1871-4021 . ПМЦ 8556684 . ПМИД 34742052 .
- ^ Аннунциата, Джузеппе; Сандуцци Зампарелли, Марко; Санторо, Чиро; Чампалья, Роберто; Сторнаиуоло, Мариано; Тенор, Джан Карло; Сандуцци, Алессандро; Новеллино, Этторе (2020). «Могут ли полифенолы сыграть роль в борьбе с коронавирусной инфекцией? Обзор данных in vitro» . Границы в медицине . 7 : 240. дои : 10.3389/fmed.2020.00240 . ISSN 2296-858X . ПМЦ 7243156 . ПМИД 32574331 .
- ^ Дай, Сяофэн; Чжан, Сюаньхао; Остриков, Костя; Абрамян, Левон (03.03.2020). «Рецепторы хозяина: ключ к созданию клеток с широким вирусным тропизмом для производства вакцин» . Критические обзоры по микробиологии . 46 (2): 147–168. дои : 10.1080/1040841X.2020.1735992 . ISSN 1040-841X . ПМЦ 7113910 . ПМИД 32202955 .
- ^ Чаппелл, Кейт Дж.; Уоттерсон, Дэниел (12 января 2017 г.). «Борьба с Эболой: окно для переоценки вакцин?» . ПЛОС Патогены . 13 (1): e1006037. дои : 10.1371/journal.ppat.1006037 . ISSN 1553-7374 . ПМК 5230751 . ПМИД 28081225 .