Ортомиксовирусиды
| Ортомиксовирусиды | |
|---|---|
 | |
| гриппа А и гриппа В Геном, мРНК и вирионная диаграмма вирусов | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Сорт: | Инстовирицеты |
| Заказ: | Артикулавирусы |
| Семья: | Ортомиксовирусиды |
| Роды | |
Orthomyxoviridae (от греческого ὀρθός, или «прямой» + μύξα, mýxa « слизь ») [1] — семейство с отрицательным смыслом РНК-вирусов . Он включает семь родов : Alphainfluenzavirus , Betainfluenzavirus , Gammainfluenzavirus , Deltainfluenzavirus , Isavirus , Thogotovirus и Quaranjavirus . Первые четыре рода содержат вирусы, вызывающие ( см грипп птиц . также птичий грипп ) и млекопитающих , включая человека. Изавирусы поражают лосося ; Тоготовирусы представляют собой арбовирусы , поражающие позвоночных и беспозвоночных (таких как клещи и комары ). [2] [3] [4] Кваранджавирусы также являются арбовирусами , поражающими позвоночных (птиц) и беспозвоночных ( членистоногих ).
Четыре рода вируса гриппа, поражающие позвоночных, которые идентифицируются по антигенным различиям в их нуклеопротеинах и матриксных белках , следующие:
- Вирус альфагриппа заражает людей, других млекопитающих и птиц и вызывает все пандемии гриппа.
- Вирус бетаингриппа заражает людей и тюленей
- Вирус гаммагриппа поражает людей и свиней.
- Дельтавирус гриппа поражает свиней и крупный рогатый скот .
Структура
[ редактировать ]
вируса Вирион плеоморфен гриппа ; вирусная оболочка может иметь сферическую и нитевидную форму. В целом морфология вируса эллипсоидная с частицами диаметром 100–120 нм или нитевидная с частицами диаметром 80–100 нм и длиной до 20 мкм. [5] В оболочке имеется около 500 различных шипообразных выступов на поверхности, каждый из которых выступает на 10–14 нм от поверхности с различной поверхностной плотностью. Главный спайк гликопротеина (НА) расположен неравномерно между кластерами шипов нейраминидазы (НА), с соотношением НА и НА примерно 10 к 1. [6]
Вирусная оболочка, состоящая из двухслойной липидной мембраны, в которой закреплены гликопротеиновые шипы, окружает нуклеокапсиды ; нуклеопротеины разных размерных классов с петлей на каждом конце; расположение внутри вириона неясно. Рибонуклеарные белки имеют нитевидную форму и имеют длину 50–130 нм и диаметр 9–15 нм со спиральной симметрией. [ нужна ссылка ]
Геном
[ редактировать ]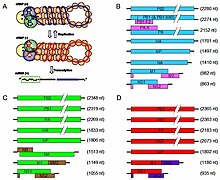
Вирусы семейства Orthomyxoviridae содержат от шести до восьми сегментов линейной с отрицательным смыслом одноцепочечной РНК . Их общая длина генома составляет 10 000–14 600 нуклеотидов (нт). [7] гриппа А геном Например, состоит из восьми фрагментов сегментированной РНК с отрицательным смыслом (всего 13,5 тыс. нуклеотидов). [8]
Наиболее изученными белками вируса гриппа являются гемагглютинин и нейраминидаза , два крупных гликопротеина, обнаруженные на внешней стороне вирусных частиц. Гемагглютинин представляет собой лектин , который опосредует связывание вируса с клетками-мишенями и проникновение вирусного генома в клетку-мишень. [9] Напротив, нейраминидаза представляет собой фермент, участвующий в высвобождении потомства вируса из инфицированных клеток путем расщепления сахаров, которые связывают зрелые вирусные частицы. гемагглютинин (H) и нейраминидаза (N) Белки являются ключевыми мишенями для антител и противовирусных препаратов. [10] [11] и они используются для классификации различных серотипов вирусов гриппа А, отсюда H и N в H5N1 .
Последовательность генома имеет терминальные повторяющиеся последовательности; повторяется с обоих концов. Терминальные повторы на 5'-конце длиной 12–13 нуклеотидов. Нуклеотидные последовательности 3'-конца идентичны; то же самое в роду того же семейства; больше всего на РНК (сегментах) или на всех видах РНК. Терминальные повторы на 3'-конце длиной 9–11 нуклеотидов. Инкапсидированная нуклеиновая кислота является исключительно геномной. Каждый вирион может содержать дефектные интерферирующие копии. При гриппе A (H1N1) PB1-F2 образуется из альтернативной рамки считывания в PB1. Гены M и NS производят два разных гена посредством альтернативного сплайсинга . [12]
Цикл репликации
[ редактировать ]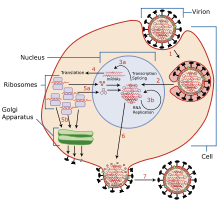
Обычно грипп передается от инфицированных млекопитающих воздушно-капельным путем при кашле или чихании, образуя аэрозоли, содержащие вирус, а также от инфицированных птиц через их помет . Грипп также может передаваться через слюну , носовые выделения , фекалии и кровь . Заражение происходит при контакте с этими биологическими жидкостями или загрязненными поверхностями. Вне хозяина вирусы гриппа могут оставаться заразными в течение примерно одной недели при температуре человеческого тела, более 30 дней при 0 °C (32 °F) и неопределенно долго при очень низких температурах (например, в озерах на северо-востоке Сибири ). Их можно легко инактивировать дезинфицирующими и моющими средствами . [13] [14] [15]
Вирусы связываются с клеткой посредством взаимодействия между ее гликопротеином гемагглютинина и сахарами сиаловой кислоты на поверхности эпителиальных клеток легких и горла (стадия 1 на рисунке инфекции). [16] Клетка импортирует вирус путем эндоцитоза . В кислой эндосоме часть белка гемагглютинина сливает вирусную оболочку с мембраной вакуоли, высвобождая вирусной РНК (вРНК), вспомогательные белки и РНК-зависимую РНК-полимеразу в цитоплазму молекулы (стадия 2). [17] Эти белки и вРНК образуют комплекс, который транспортируется в ядро клетки , где РНК-зависимая РНК-полимераза начинает транскрибировать комплементарную кРНК с положительным смыслом (шаги 3a и b). [18] кРНК либо экспортируется в цитоплазму и транслируется (этап 4), либо остается в ядре. Вновь синтезированные вирусные белки либо секретируются через аппарат Гольджи на поверхность клетки (в случае нейраминидазы и гемагглютинина, этап 5б), либо транспортируются обратно в ядро для связывания вРНК и формирования новых частиц вирусного генома (этап 5а). Другие вирусные белки оказывают множественное действие на клетку-хозяина, включая деградацию клеточной мРНК и использование высвободившихся нуклеотидов для синтеза вРНК, а также ингибирование трансляции мРНК клетки-хозяина. [19]
ВРНК с отрицательным смыслом, формирующие геномы будущих вирусов, РНК-зависимая РНК-транскриптаза и другие вирусные белки собираются в вирион. Молекулы гемагглютинина и нейраминидазы собираются в выпуклость клеточной мембраны. ВРНК и коровые белки вируса покидают ядро и попадают в выступ мембраны (шаг 6). Зрелый вирус отпочковывается от клетки в сфере фосфолипидной мембраны хозяина, приобретая гемагглютинин и нейраминидазу с помощью этой мембранной оболочки (этап 7). [20] Как и раньше, вирусы прикрепляются к клетке посредством гемагглютинина; зрелые вирусы отделяются, как только их нейраминидаза отщепляет остатки сиаловой кислоты от клетки-хозяина. [16] После высвобождения нового вируса гриппа клетка-хозяин погибает.

Ортомиксовирусные вирусы — один из двух РНК-вирусов, которые реплицируются в ядре (второй — ретровирусы ). Это связано с тем, что механизм ортомиксовирусов не может создавать собственные мРНК. Они используют клеточные РНК в качестве праймеров для инициации синтеза вирусной мРНК в процессе, известном как захват кэпа . [21] Попав в ядро, белок РНК-полимеразы PB2 находит клеточную пре-мРНК и связывается с ее 5'-концом. Затем РНК-полимераза PA отщепляет клеточную мРНК вблизи 5'-конца и использует этот кэпированный фрагмент в качестве праймера для транскрипции остальной части генома вирусной РНК в вирусную мРНК. [22] клетки Это связано с тем, что мРНК должна иметь 5'-кэп, чтобы ее можно было распознать рибосомой для трансляции.
Поскольку ферменты , проверяющие РНК , отсутствуют, РНК-зависимая РНК-транскриптаза допускает ошибку вставки одного нуклеотида примерно каждые 10 тысяч нуклеотидов, что соответствует приблизительной длине вРНК гриппа. Следовательно, почти каждый вновь созданный вирус гриппа будет содержать мутацию в своем геноме. [23] Разделение генома на восемь отдельных сегментов вРНК позволяет смешивать ( реассортировать ) гены, если одну и ту же клетку инфицирует более одной разновидности вируса гриппа ( суперинфекция ). Возникающее в результате изменение сегментов генома, упакованных в вирусное потомство, придает новое поведение, иногда способность заражать новые виды хозяев или преодолевать защитный иммунитет популяций хозяев к старому геному (в этом случае это называется антигенным сдвигом ). [10]
Классификация
[ редактировать ]В филогенетической оцРНК-вирусов с таксономии категория РНК-вирусов включает подкатегорию отрицательным смыслом , которая включает порядок Articulavirales и семейство Orthomyxoviridae . Связанные с родами виды и серотипы Orthomyxoviridae . показаны в следующей таблице
| Род | Виды (* указывает типовой вид ) | Серотипы или подтипы | Хозяева |
|---|---|---|---|
| Альфагрипповирус | Вирус гриппа А * | H1N1 , H1N2 , H2N2 , H3N1 , H3N2 , H3N8 , H5N1 , H5N2 , H5N3 , H5N8 , H5N9 , H7N1 , H7N2 , H7N3 , H7N4 , H7N7 , H7N9 , H9N2 , H10N7 | Человек , свинья , птица , лошадь , летучая мышь |
| Бетаинвирус гриппа | Вирус гриппа B * | Победа, Ямагата [24] | Человек, тюлень |
| Гаммагрипповирус | Вирус гриппа С * | Человек, свинья | |
| Дельтавирус гриппа | Вирус гриппа D * | Свинья, крупный рогатый скот | |
| Исавирус | Вирус инфекционной анемии лосося * | Атлантический лосось | |
| Тоготовирус | Тоготовирус * | Клещ , комар , млекопитающее (в том числе и человек) | |
| Вирус Дори | Вирус Баткен , вирус Бурбона , вирус Джоса. | ||
| Каранджавирус [25] | |||
| Вирус Quaranfil ,* вирус атолла Джонстон |
Типы
[ редактировать ]Существует четыре рода вирусов гриппа, каждый из которых содержит только один вид или тип. Грипп A и C поражают различные виды животных (включая человека), тогда как грипп B поражает почти исключительно людей, а грипп D поражает крупный рогатый скот и свиней. [26] [27] [28]
Грипп А
[ редактировать ]
Вирусы гриппа А дополнительно классифицируются на основе вирусных поверхностных белков гемагглютинина (HA или H) и нейраминидазы (NA или N). В природе выделено 18 подтипов (или серотипов) НА и 11 подтипов NA вируса гриппа А. Среди них подтипы НА 1–16 и подтипы NA 1–9 обнаружены у диких водоплавающих и околоводных птиц, а подтипы HA 17–18 и подтипы NA 10–11 были выделены только у летучих мышей. [29] [30]
Существуют и другие вариации; таким образом, специфические изоляты штаммов гриппа идентифицируются по номенклатуре вирусов гриппа, [31] с указанием типа вируса, вида хозяина (если не человека), географического местоположения, где впервые был выделен, лабораторной ссылки, года выделения, а также подтипов HA и NA. [32] [33]
Примеры номенклатуры:
- A/Brisbane/59/2007 (H1N1) - выделен от человека.
- A/swine/South Dakota/152B/2009 (H1N2) - выделен от свиньи.
Вирусы гриппа типа А являются наиболее вирулентными патогенами для человека среди трех типов гриппа и вызывают наиболее тяжелое заболевание. Считается, что все вирусы гриппа А, вызывающие вспышки или пандемии, происходят от диких водоплавающих птиц. [34] Все пандемии вируса гриппа А, начиная с 1900-х годов, были вызваны птичьим гриппом в результате реассортации с другими штаммами гриппа, поражающими либо людей (сезонный грипп), либо других животных (см. пандемию свиного гриппа 2009 г. ). [35] Серотипы, подтвержденные у людей , упорядочены по количеству подтвержденных смертей людей:
- H1N1 вызвал « испанку » в 1918 году и « свиной грипп » в 2009 году. [36]
- H2N2 вызвал «азиатский грипп».
- H3N2 вызвал « гонконгский грипп ».
- H5N1 , «птичий» или «птичий грипп». [37]
- H7N7 обладает необычным зоонозным потенциалом. [38]
- H1N2 заражает свиней и людей. [39]
- Х9Н2 , Х7Н2 , Х7Н3 , Х10Н7 .
| Название пандемии | Дата | Летальные исходы | Уровень смертности | Задействован подтип | Индекс серьезности пандемии |
|---|---|---|---|---|---|
| Пандемия гриппа 1889–1890 гг. (Азиатский или Русский грипп) [42] | 1889–1890 | 1 миллион | 0.15% | Возможно H3N8 или H2N2 | — |
| Пандемия гриппа 1918 года (Испанка) [43] | 1918–1920 | от 20 до 100 миллионов | 2% | H1N1 | 5 |
| Азиатский грипп | 1957–1958 | от 1 до 1,5 миллиона | 0.13% | H2N2 | 2 |
| Гонконгский грипп | 1968–1969 | от 0,75 до 1 миллиона | <0,1% | H3N2 | 2 |
| Русский грипп | 1977–1978 | Нет точного подсчета | — | H1N1 | — |
| пандемия гриппа 2009 г. [44] [45] | 2009–2010 | 105,700–395,600 [46] | 0.03% | H1N1 | Н/Д |
Грипп Б
[ редактировать ]
Вирус гриппа B почти исключительно является человеческим возбудителем и встречается реже, чем вирус гриппа A. Известно, что единственным другим животным, восприимчивым к инфекции гриппа B, является тюлень . [47] Этот тип гриппа мутирует со скоростью в 2–3 раза меньшей, чем тип А. [48] и, следовательно, менее генетически разнообразен, имея только один серотип гриппа B. [26] В результате отсутствия антигенного разнообразия определенная степень иммунитета к гриппу B обычно приобретается в раннем возрасте. Однако грипп B настолько мутирует, что устойчивый иммунитет невозможен. [49] Эта сниженная скорость антигенных изменений в сочетании с ограниченным кругом хозяев (ингибирование межвидового антигенного сдвига ) гарантирует отсутствие пандемий гриппа B. [50]
Грипп С
[ редактировать ]Вирус гриппа С поражает людей и свиней и может вызывать тяжелые заболевания и местные эпидемии . [51] Однако грипп С встречается реже, чем другие типы, и обычно вызывает легкое заболевание у детей. [52] [53]
Грипп Д
[ редактировать ]Это род, классифицированный в 2016 году, представители которого впервые были выделены в 2011 году. [54] Этот род, по-видимому, наиболее близок к гриппу C, от которого он отделился несколько сотен лет назад. [55] Существует как минимум два современных штамма этого рода. [56] Основными хозяевами, по-видимому, является крупный рогатый скот, но известно, что вирус заражает и свиней.
Жизнеспособность и дезинфекция
[ редактировать ]Вирусы гриппа млекопитающих, как правило, лабильны, но могут сохраняться в слизи в течение нескольких часов. [57] Вирус птичьего гриппа может выжить 100 дней в дистиллированной воде при комнатной температуре и 200 дней при 17 °C (63 °F). Птичий вирус быстрее инактивируется в навозе, но может сохраняться до двух недель в фекалиях клеток. Вирусы птичьего гриппа могут сохраняться в замороженном состоянии неопределенно долго. [57] Вирусы гриппа чувствительны к отбеливателю, 70% этанолу, альдегидам, окислителям и соединениям четвертичного аммония. Они инактивируются при нагревании до 133 °F (56 °C) в течение минимум 60 минут, а также при низком pH <2. [57]
Вакцинация и профилактика
[ редактировать ]
Доступны вакцины и лекарства для профилактики и лечения инфекций, вызванных вирусом гриппа. Вакцины состоят из инактивированных или живых аттенуированных вирионов вирусов человеческого гриппа A H1N1 и H3N2, а также вирусов гриппа B. Поскольку антигенность диких вирусов меняется, состав вакцин ежегодно обновляется путем обновления посевных штаммов. [ нужна ссылка ]
Когда антигенность семенных штаммов и диких вирусов не совпадает, вакцины не могут защитить привитых. [ нужна ссылка ] Кроме того, даже если они совпадают, часто возникают мутанты-побеги. [ нужна ссылка ]
Лекарства, доступные для лечения гриппа, включают амантадин и римантадин , которые ингибируют снятие оболочки вирионов, вмешиваясь в М2, и осельтамивир (продаваемый под торговой маркой Тамифлю ), занамивир и перамивир , которые ингибируют высвобождение вирионов из инфицированных клеток, вмешиваясь в процесс. с Н.А. Однако для первого препарата часто возникают ускользающие мутанты, а для второго — реже. [58]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ вирусов Международного комитета по таксономии Индекс вирусов — ортомиксовирус (2006). В: ICTVdB — Универсальная база данных вирусов, версия 4. Бюхен-Осмонд, C (ред), Колумбийский университет, Нью-Йорк.
- ^ Джонс Л.Д., Наттолл, Пенсильвания (1989). «Невиремическая передача вируса Тогото: влияние времени и расстояния». Пер. Р. Сок. Троп. Мед. Хиг . 83 (5): 712–14. дои : 10.1016/0035-9203(89)90405-7 . ПМИД 2617637 .
- ^ Эли Б. (1999). «Инфекционная лососевая анемия» . Очерки Милл-Хилла . Национальный институт медицинских исследований . Архивировано из оригинала 24 августа 2007 г. Проверено 14 сентября 2007 г.
- ^ Рейнард Р.С., Мюррей А.Г., Грегори А. (2001). «Вирус инфекционной анемии лосося у диких рыб из Шотландии» . Дис. Акват. Орг . 46 (2): 93–100. дои : 10.3354/dao046093 . ПМИД 11678233 .
- ^ Нода Т (3 января 2012 г.). «Нативная морфология вирионов гриппа» . Границы микробиологии . 2 : 269. дои : 10.3389/fmicb.2011.00269 . ПМЦ 3249889 . ПМИД 22291683 .
- ^ Эйнав Т., Джентлес Л.Е., Блум Дж.Д. (23 июля 2020 г.). «SnapShot: Грипп в цифрах» (PDF) . Клетка . 182 (2): 532–532.e1. doi : 10.1016/j.cell.2020.05.004 . ПМИД 32707094 . S2CID 220715148 .
- ^ «Девятый отчет ICTV; выпуск таксономии 2009 г.: Orthomyxoviridae » . ICTV . Проверено 19 сентября 2020 г.
- ^ Гедин Э, Сенгамалай Н.А., Шамуэй М, Заборский Дж, Фельдблюм Т, Суббу В, Спиро DJ, Ситц Дж, Ку Х, Болотов П, Дерновой Д, Татусова Т, Бао Ю, Сент-Джордж К, Тейлор Дж, Липман DJ, Фрейзер КМ, Таубенбергер Дж.К., Зальцберг С.Л. (октябрь 2005 г.). «Крупномасштабное секвенирование человеческого гриппа раскрывает динамическую природу эволюции вирусного генома» . Природа . 437 (7062): 1162–6. Бибкод : 2005Natur.437.1162G . дои : 10.1038/nature04239 . ПМИД 16208317 .
- ^ Сузуки Y (март 2005 г.). «Сиалобиология гриппа: молекулярный механизм изменения диапазона хозяев вирусов гриппа» . Биологический и фармацевтический вестник . 28 (3): 399–408. дои : 10.1248/bpb.28.399 . ПМИД 15744059 .
- ^ Jump up to: а б с Хиллеман М.Р. (август 2002 г.). «Реалии и загадки вирусного гриппа человека: патогенез, эпидемиология и контроль». Вакцина . 20 (25–26): 3068–87. дои : 10.1016/S0264-410X(02)00254-2 . ПМИД 12163258 .
- ^ Уилсон Дж.К., фон Ицштайн М. (июль 2003 г.). «Последние стратегии в поиске новых методов лечения гриппа». Текущие цели по борьбе с наркотиками . 4 (5): 389–408. дои : 10.2174/1389450033491019 . ПМИД 12816348 .
- ^ Бувье Н.М., Палезе П. (сентябрь 2008 г.). «Биология вирусов гриппа» . Вакцина . 26 (Приложение 4): D49–53. doi : 10.1016/j.vaccine.2008.07.039 . ПМК 3074182 . ПМИД 19230160 .
- ^ Суарес Д.Л., Спакман Э., Сенне Д.А., Булага Л., Уэлш А.С., Фроберг К. (2003). «Влияние различных дезинфицирующих средств на обнаружение вируса птичьего гриппа с помощью RT-PCR в реальном времени». Птичьи болезни . 47 (3 приложения): 1091–5. дои : 10.1637/0005-2086-47.s3.1091 . ПМИД 14575118 . S2CID 8612187 .
- ^ «Последствия птичьего гриппа (птичьего гриппа) для заболеваний человека. Физические характеристики вирусов гриппа А» . CIDRAP — Центр исследований и политики в области инфекционных заболеваний . Университет Миннесоты. 12 марта 2024 г.
- ^ «Вирусы гриппа могут жить на льду десятилетиями» . Новозеландский Вестник . Рейтер . 30 ноября 2006 года . Проверено 1 ноября 2011 г.
- ^ Jump up to: а б Вагнер Р., Матросович М., Кленк Х. (май – июнь 2002 г.). «Функциональный баланс между гемагглютинином и нейраминидазой при инфекциях вируса гриппа». Преподобный Мед Вирол . 12 (3): 159–66. дои : 10.1002/rmv.352 . ПМИД 11987141 . S2CID 30876482 .
- ^ Лакадамиали М., Раст М., Бэбкок Х., Чжуан Икс (5 августа 2003 г.). «Визуализация заражения отдельными вирусами гриппа» . Proc Natl Acad Sci США . 100 (16): 9280–85. Бибкод : 2003PNAS..100.9280L . дои : 10.1073/pnas.0832269100 . ПМК 170909 . ПМИД 12883000 .
- ^ Крос Дж., Палезе П. (сентябрь 2003 г.). «Торговля вирусной геномной РНК в ядро и из него: вирусы гриппа, болезни Тогото и Борна». Вирус Рес . 95 (1–2): 3–12. дои : 10.1016/S0168-1702(03)00159-X . ПМИД 12921991 .
- ^ Каш Дж., Гудман А., Корт М., Катце М. (июль 2006 г.). «Похищение реакции клетки-хозяина и контроль трансляции во время заражения вирусом гриппа». Вирус Рес . 119 (1): 111–20. doi : 10.1016/j.virusres.2005.10.013 . ПМИД 16630668 .
- ^ Наяк Д., Хуэй Э., Барман С. (декабрь 2004 г.). «Сборка и почкование вируса гриппа» . Вирус Рес . 106 (2): 147–65. doi : 10.1016/j.virusres.2004.08.012 . ПМЦ 7172797 . ПМИД 15567494 .
- ^ «Похищение кепки» . ВиралЗона . экспази . Проверено 11 сентября 2014 г.
- ^ Диас А., Бувье Д., Крепен Т., Маккарти А.А., Харт DJ, Боден Ф., Кьюсак С., Рюигрок Р.В. (апрель 2009 г.). «Эндонуклеаза, захватывающая кепку полимеразы вируса гриппа, находится в субъединице PA». Природа . 458 (7240): 914–8. Бибкод : 2009Natur.458..914D . дои : 10.1038/nature07745 . ПМИД 19194459 . S2CID 4421958 .
- ^ Дрейк Дж. (1 мая 1993 г.). «Уровень спонтанных мутаций среди РНК-вирусов» . Proc Natl Acad Sci США . 90 (9): 4171–5. Бибкод : 1993PNAS...90.4171D . дои : 10.1073/pnas.90.9.4171 . ПМК 46468 . ПМИД 8387212 .
- ^ Бьер Б., Бауэр Б., Швайгер Б. (апрель 2010 г.). «Дифференциация линий вируса гриппа B Ямагата и Виктория с помощью ПЦР в реальном времени» (PDF) . Журнал клинической микробиологии . 48 (4): 1425–7. дои : 10.1128/JCM.02116-09 . ПМЦ 2849545 . ПМИД 20107085 .
- ^ История таксономии ICTV , ICTV, 2014, заархивировано из оригинала 2 апреля 2015 года , получено 6 июня 2006 года.
- ^ Jump up to: а б Хэй А., Грегори В., Дуглас А., Лин Ю. (29 декабря 2001 г.). «Эволюция вирусов гриппа человека» . Philos Trans R Soc Lond B Biol Sci . 356 (1416): 1861–70. дои : 10.1098/rstb.2001.0999 . ПМЦ 1088562 . ПМИД 11779385 .
- ^ «Птичий грипп (птичий грипп)» . Центры по контролю и профилактике заболеваний . Проверено 15 сентября 2007 г.
- ^ Кумар, Бинод; Аша, Кумари; Кханна, Мадху; Ронсар, Ларанс; Месеко, Клемент Адебахо; Саникас, Мелвин (апрель 2018 г.). «Новая угроза вируса гриппа: состояние и новые перспективы его терапии и контроля» . Архив вирусологии . 163 (4): 831–844. дои : 10.1007/s00705-018-3708-y . ISSN 1432-8798 . ПМК 7087104 . ПМИД 29322273 .
- ^ Александр, Деннис Дж. (26 июля 2007 г.). «Обзор эпидемиологии птичьего гриппа» . Вакцина . 25 (30): 5637–5644. doi : 10.1016/j.vaccine.2006.10.051 . ISSN 0264-410X . ПМИД 17126960 .
- ^ Ву, Ин; Ву, Ян; Тефсен, Борис; Ши, Йи; Гао, Джордж Ф. (апрель 2014 г.). «Гриппоподобные вирусы летучих мышей H17N10 и H18N11» . Тенденции в микробиологии . 22 (4): 183–191. дои : 10.1016/j.tim.2014.01.010 . ISSN 0966-842X . ПМЦ 7127364 . ПМИД 24582528 .
- ^ «Пересмотр системы номенклатуры вирусов гриппа: Меморандум ВОЗ» . Бюллетень Всемирного органа здравоохранения . 58 (4): 585–591. 1980. ПМК 2395936 . ПМИД 6969132 .
Этот Меморандум был составлен подписантами, перечисленными на странице 590, по случаю встречи, состоявшейся в Женеве в феврале 1980 года.
- ^ Аткинсон В., Хамборски Дж., Макинтайр Л., Вулф С., ред. (2007). Эпидемиология и профилактика болезней, предупреждаемых с помощью вакцин (10-е изд.). Вашингтон, округ Колумбия: Центры по контролю и профилактике заболеваний.
- ^ «Птичий грипп (птичий грипп): последствия для болезней человека» . Центр исследований и политики в области инфекционных заболеваний, Университет Миннесоты . 27 июня 2007 г. Проверено 14 сентября 2007 г.
- ^ Вебстер, Р.Г.; Бин, Вашингтон; Горман, ОТ; Чемберс, ТМ; Каваока, Ю. (март 1992 г.). «Эволюция и экология вирусов гриппа А» . Микробиологические обзоры . 56 (1): 152–179. дои : 10.1128/мр.56.1.152-179.1992 . ISSN 0146-0749 . ПМЦ 372859 . ПМИД 1579108 .
- ^ Таубенбергер, Джеффри К.; Моренс, Дэвид М. (апрель 2010 г.). «Грипп: пандемия прошлого и будущего» . Отчеты общественного здравоохранения . 125 (Приложение 3): 16–26. дои : 10.1177/00333549101250S305 . ISSN 0033-3549 . ПМЦ 2862331 . ПМИД 20568566 .
- ^ Ван Т.Т., Палезе П. (июнь 2009 г.). «Разгадка тайны вируса свиного гриппа» . Клетка . 137 (6): 983–85. дои : 10.1016/j.cell.2009.05.032 . ПМИД 19524497 .
- ^ Таубенбергер, Дж. К., Моренс, Д. М. (апрель 2009 г.). «Пандемический грипп – включая оценку риска H5N1» . Преподобный учёный. Тех. Выключенный. Межд. Эпиз . 28 (1): 187–202. doi : 10.20506/rst.28.1.1879 . ПМК 2720801 . ПМИД 19618626 .
- ^ Фушье Р., Шнебергер П., Розендал Ф., Брукман Дж., Кеминк С., Мюнстер В., Куикен Т., Риммельцваан Г., Шуттен М., Ван Дорнум Г., Кох Г., Босман А., Купманс М., Остерхаус А. (2004). «Вирус птичьего гриппа А (H7N7), связанный с конъюнктивитом человека и смертельным случаем острого респираторного дистресс-синдрома» . Proc Natl Acad Sci США . 101 (5): 1356–61. Стартовый код : 2004PNAS..101.1356F . дои : 10.1073/pnas.0308352100 . ПМК 337057 . ПМИД 14745020 .
- ^ Малик-Пейрис Дж. С., Пун Л. Л., Гуань Ю (июль 2009 г.). «Появление нового вируса свиного гриппа А (S-OIV) H1N1 у людей» . Дж. Клин Вирол . 45 (3): 169–173. дои : 10.1016/j.jcv.2009.06.006 . ПМЦ 4894826 . ПМИД 19540800 .
- ^ Поттер CW (октябрь 2001 г.). «История гриппа». Журнал прикладной микробиологии . 91 (4): 572–9. дои : 10.1046/j.1365-2672.2001.01492.x . ПМИД 11576290 . S2CID 26392163 .
- ^ «Десять вещей, которые вам нужно знать о пандемическом гриппе» . Всемирная организация здравоохранения. 14 октября 2005 г. Архивировано из оригинала 23 сентября 2009 г. Проверено 26 сентября 2009 г.
- ^ Валлерон А.Дж., Кори А., Валта С., Мерисс С., Каррат Ф., Боэль П.Ю. (май 2010 г.). «Заразность и географическое распространение пандемии гриппа 1889 года» . Учеб. Натл. акад. наук. США . 107 (19): 8778–81. Бибкод : 2010PNAS..107.8778V . дои : 10.1073/pnas.1000886107 . ПМЦ 2889325 . ПМИД 20421481 .
- ^ Миллс С.Э., Робинс Дж.М. , Липсич М. (декабрь 2004 г.). «Заразность пандемического гриппа 1918 года» . Природа . 432 (7019): 904–06. Бибкод : 2004Natur.432..904M . дои : 10.1038/nature03063 . ПМК 7095078 . ПМИД 15602562 .
- ^ Дональдсон Л.Дж., Раттер П.Д., Эллис Б.М. и др. (2009). «Смертность от пандемического гриппа A/H1N1 2009 в Англии: исследование общественного здравоохранения» . БМЖ . 339 : b5213. дои : 10.1136/bmj.b5213 . ПМК 2791802 . ПМИД 20007665 .
- ^ «Ежедневный обзор ECDC – Пандемия (H1N1) 2009 г. – 18 января 2010 г.» (PDF) . Европейский центр профилактики и контроля заболеваний . 18 января 2010 г. Архивировано из оригинала (PDF) 22 января 2010 года . Проверено 18 января 2010 г.
- ^ Давуд Ф.С., Юлиано А.Д., Рид С., Мельцер М.И., Шей Д.К., Ченг П.Ю., Бандаранаяке Д., Брейман Р.Ф., Брукс В.А., Бучи П., Фейкин Д.Р., Фаулер К.Б., Гордон А., Хиен Н.Т., Хорби П., Хуанг К.С., Кац М.А. , Кришнан А., Лал Р., Монтгомери Дж. М., Молбак К., Пебоди Р., Пресанис А. М., Разури Х., Стинс А., Тиноко Ю. О., Валлинга Дж., Ю Х., Вонг С., Брези Дж., Виддоусон М. А. (сентябрь 2012 г.). «Оценочная глобальная смертность, связанная с первыми 12 месяцами циркуляции вируса пандемического гриппа A H1N1 в 2009 году: моделирование» . «Ланцет». Инфекционные болезни (Представлена рукопись). 12 (9): 687–95. дои : 10.1016/S1473-3099(12)70121-4 . ПМИД 22738893 . Архивировано (PDF) из оригинала 9 апреля 2024 г. – через Zenodo.
- ^ Остерхаус А.Д., Риммельцваан Г.Ф., Мартина Б.Е., Бестеброер Т.М., Фушье Р.А. (май 2000 г.). «Вирус гриппа B у тюленей». Наука . 288 (5468): 1051–3. Бибкод : 2000Sci...288.1051O . дои : 10.1126/science.288.5468.1051 . ПМИД 10807575 .
- ^ Нобусава Э., Сато К. (апрель 2006 г.). «Сравнение частоты мутаций вирусов гриппа человека А и В» . Журнал вирусологии . 80 (7): 3675–8. doi : 10.1128/JVI.80.7.3675-3678.2006 . ПМК 1440390 . ПМИД 16537638 .
- ^ Вебстер Р.Г., Бин В.Дж., Горман О.Т., Чемберс Т.М., Каваока Ю. (март 1992 г.). «Эволюция и экология вирусов гриппа А» . Микробиологические обзоры . 56 (1): 152–79. дои : 10.1128/MMBR.56.1.152-179.1992 . ПМЦ 372859 . ПМИД 1579108 .
- ^ Zambon MC (ноябрь 1999 г.). «Эпидемиология и патогенез гриппа» . Журнал антимикробной химиотерапии . 44 Приложение Б (Приложение Б): 3–9. дои : 10.1093/jac/44.suppl_2.3 . ПМИД 10877456 . S2CID 15968981 .
- ^ Мацузаки Ю, Сугавара К, Мизута К, Цучия Э, Мураки Ю, Хонго С, Сузуки Х, Накамура К (2002). «Антигенная и генетическая характеристика вирусов гриппа С, вызвавших две вспышки в городе Ямагата, Япония, в 1996 и 1998 годах» . J Clin Микробиол . 40 (2): 422–29. doi : 10.1128/JCM.40.2.422-429.2002 . ПМЦ 153379 . ПМИД 11825952 .
- ^ Мацузаки Ю, Кацусима Н, Нагай Ю, Сёдзи М, Итагаки Т, Сакамото М, Китаока С, Мизута К, Нисимура Х (1 мая 2006 г.). «Клинические особенности инфекции вируса гриппа С у детей» . J Заразить Дис . 193 (9): 1229–35. дои : 10.1086/502973 . ПМИД 16586359 .
- ^ Катагири С., Охизуми А., Хомма М. (июль 1983 г.). «Вспышка гриппа типа С в детском доме». J Заразить Дис . 148 (1): 51–56. дои : 10.1093/infdis/148.1.51 . ПМИД 6309999 .
- ^ Хаус Б.М., Дукатес М., Коллин Э.А., Ран З., Лю Р., Шэн З., Армиен А., Каплан Б., Чакраварти С., Хоппе А.Д., Уэбби Р.Дж., Симонсон Р.Р., Ли Ф (февраль 2013 г.). «Выделение нового вируса свиного гриппа из Оклахомы в 2011 году, который имеет отдаленное родство с вирусами человеческого гриппа С» . ПЛОС Патогены . 9 (2): e1003176. дои : 10.1371/journal.ppat.1003176 . ПМК 3567177 . ПМИД 23408893 .
- ^ Шэн З., Ран З., Ван Д., Хоппе А.Д., Симонсон Р., Чакраварти С., Хаус Б.М., Ли Ф (февраль 2014 г.). «Геномная и эволюционная характеристика нового вируса гриппа С-подобного свиного происхождения» . Архив вирусологии . 159 (2): 249–55. дои : 10.1007/s00705-013-1815-3 . ПМЦ 5714291 . ПМИД 23942954 .
- ^ Коллин Э.А., Шэн З., Ланг Й., Ма В., Хаус Б.М., Ли Ф. (январь 2015 г.). «Коциркуляция двух различных генетических и антигенных линий предполагаемого вируса гриппа D у крупного рогатого скота» . Журнал вирусологии . 89 (2): 1036–42. дои : 10.1128/JVI.02718-14 . ПМК 4300623 . ПМИД 25355894 .
- ^ Jump up to: а б с Спиклер А.Р. (февраль 2016 г.). «Грипп» (PDF) . Центр продовольственной безопасности и общественного здравоохранения . Университет штата Айова. п. 7.
- ^ Сузуки Y (октябрь 2006 г.). «Естественный отбор в геноме вируса гриппа» . Молекулярная биология и эволюция . 23 (10). Оксфордский академический: 1902–11. дои : 10.1093/molbev/msl050 . ПМИД 16818477 . Архивировано из оригинала 21 января 2022 года.
Дальнейшее чтение
[ редактировать ]- Хойл, Л. (1969). Вирусы гриппа . Монографии по вирусологии. Том. 4. Шпрингер-Верлаг. ISBN 978-3-211-80892-4 . ISSN 0083-6591 . OCLC 4053391 .
Внешние ссылки
[ редактировать ]- Портал Здоровье-ЕС : ЕС работает над подготовкой глобального ответа на грипп.
- База данных исследований гриппа : база данных геномных последовательностей гриппа и сопутствующей информации.
- Европейская комиссия – Общественное здравоохранение : координация ЕС по борьбе с пандемией (H1N1), 2009 г.
- 3D-структуры, связанные с вирусом гриппа, из Банка данных EM (EMDB)
- Вирусная зона: Ортомиксовирусы.
- Таксономия вирусов: выпуск 2020 г .: Международный комитет по таксономии вирусов (ICTV)

| Базы данных органов управления : Национальные |
|---|