Сомит
| Сомит | |
|---|---|
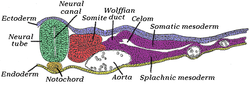 Поперечный срез половины куриного эмбриона сорокапятичасовой инкубации. Дорсальная (задняя) поверхность эмбриона находится вверху этой страницы, а вентральная (передняя) поверхность — внизу. | |
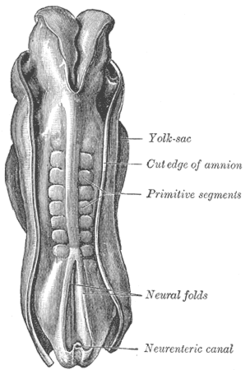 Дорсум человеческого эмбриона, длина 2,11 мм. более старый термин «примитивные сегменты ».) ( Для идентификации сомитов используется | |
| Подробности | |
| Этап Карнеги | 9 |
| Дни | 20 [1] |
| Предшественник | Параксиальная мезодерма |
| Дает начало | Дерматом , миотом , склеротом , синдетом |
| Идентификаторы | |
| латинский | прорицатель |
| МеШ | Д019170 |
| ТО | Е5.0.2.2.2.0.3 |
| ФМА | 85522 |
| Анатомическая терминология | |
Сомиты эмбриональной (устаревший термин: примитивные сегменты ) — совокупность билатерально парных блоков параксиальной мезодермы , образующихся на стадии сомитогенеза , вдоль оси голова-хвост у сегментированных животных. У позвоночных сомиты подразделяются на дерматомы , миотомы , склеротомы и синдетомы , дающие начало позвонкам позвоночного столба , грудной клетке , части затылочной кости , скелетным мышцам , хрящам , сухожилиям и коже (спины). [2]
Слово «сомит» иногда также используется вместо слова «метамер» . В этом определении сомит представляет собой гомологично животного -парную структуру в строении тела , например, наблюдаемую у кольчатых червей и членистоногих . [3]
Разработка
[ редактировать ]
Мезодерма зародышевым формируется одновременно с двумя другими листками — эктодермой и энтодермой . Мезодерма по обе стороны нервной трубки называется параксиальной мезодермой . Она отличается от мезодермы под нервной трубкой, которая называется хордамезодермой и становится хордой. Параксиальную мезодерму первоначально называют «сегментарной пластинкой» у куриного эмбриона или «несегментированной мезодермой» у других позвоночных. По мере того как примитивная полоска регрессирует и нервные складки собираются (что в конечном итоге становится нервной трубкой ), параксиальная мезодерма разделяется на блоки, называемые сомитами. [4]
Формирование
[ редактировать ]
Пресомитная мезодерма принимает на себя сомитную судьбу до того, как мезодерма становится способной образовывать сомиты. Клетки внутри каждого сомита определяются в зависимости от их местоположения внутри сомита. Кроме того, они сохраняют способность превращаться в любую структуру, происходящую из сомитов, до относительно поздней стадии процесса сомитогенеза . [4]
Развитие сомитов зависит от часового механизма, описываемого моделью часов и волнового фронта . В одном описании модели Notch и Wnt тактовые сигналы обеспечивают колеблющиеся сигналы . Волна представляет собой градиент белка фактора роста фибробластов от рострального к каудальному (градиент от носа к хвосту). Сомиты образуются один за другим по всей длине зародыша от головы до хвоста, причем каждый новый сомит формируется на каудальной (хвостовой) стороне предыдущего. [5] [6]
Время интервала не является универсальным. У разных видов разные интервалы времени. У куриного эмбриона сомиты образуются каждые 90 минут. У мыши интервал составляет 2 часа. [7]
Для некоторых видов количество сомитов может использоваться для определения стадии эмбрионального развития более надежно, чем количество часов после оплодотворения, поскольку на скорость развития могут влиять температура или другие факторы окружающей среды. Сомиты появляются по обе стороны нервной трубки одновременно. Экспериментальные манипуляции с развивающимися сомитами не изменят ростральную/каудальную ориентацию сомитов, поскольку судьбы клеток определяются до сомитогенеза. Образование сомитов может быть индуцировано клетками, секретирующими Noggin . Количество сомитов зависит от вида и не зависит от размера эмбриона (например, если он модифицирован хирургическим путем или генной инженерией). Куриные эмбрионы имеют 50 сомитов; у мышей их 65, а у змей 500. [4] [8]
Когда клетки внутри параксиальной мезодермы начинают объединяться, их называют сомитомерами , что указывает на отсутствие полного разделения между сегментами. Наружные клетки подвергаются мезенхимально-эпителиальному переходу с образованием эпителия вокруг каждого сомита. Внутренние клетки остаются в виде мезенхимы .
Сигнализация метки
[ редактировать ]Система Notch, как часть модели часов и волнового фронта, формирует границы сомитов. DLL1 и DLL3 являются лигандами Notch , мутации которых вызывают различные дефекты. Notch регулирует HES1 , который формирует каудальную половину сомитов. Активация Notch включает LFNG , который, в свою очередь, ингибирует рецептор Notch. Активация Notch также включает ген HES1, который инактивирует LFNG, повторно активируя рецептор Notch и, таким образом, объясняя модель осциллирующих часов. MESP2 индуцирует ген EPHA4 , который вызывает отталкивающее взаимодействие, которое разделяет сомиты, вызывая сегментацию. EPHA4 ограничен границами сомитов. EPHB2 также важен для границ.
Мезенхимально-эпителиальный переход
[ редактировать ]Фибронектин и N-кадгерин играют ключевую роль в процессе мезенхимально-эпителиального перехода в развивающемся эмбрионе. Вероятно, этот процесс регулируется параксисом и MESP2. В свою очередь, MESP2 регулируется передачей сигналов Notch. Параксис регулируется процессами, в которых участвует цитоскелет .
Спецификация
[ редактировать ]
Гены Hox специфицируют сомиты в целом на основе их положения вдоль передне-задней оси посредством спецификации пресомитной мезодермы до того, как произойдет сомитогенез. После образования сомитов их идентичность в целом уже определена, о чем свидетельствует тот факт, что трансплантация сомитов из одного региона в совершенно другой регион приводит к образованию структур, обычно наблюдаемых в исходном регионе. Напротив, клетки внутри каждого сомита сохраняют пластичность (способность образовывать любые структуры) до относительно позднего периода сомитного развития. [4]
Производные
[ редактировать ]
У развивающегося эмбриона позвоночных сомиты расщепляются с образованием дерматомов, скелетных мышц (миотомов), сухожилий и хрящей (синдетомов). [9] и кости (склеротомы).
Поскольку склеротом дифференцируется раньше дерматома и миотома, термин «дермомиотом» относится к объединенному дерматому и миотому до их разделения. [10]
Дерматом
[ редактировать ]Дерматом — это дорсальная часть параксиального сомита мезодермы , из которой образуется кожа ( дерма ). У эмбриона человека он возникает на третьей неделе эмбриогенеза . [2] Он образуется, когда дермомиотом (оставшаяся часть сомита, оставшаяся при миграции склеротома) расщепляется с образованием дерматома и миотома. [2] В состав дерматомов входят кожа, жир и соединительная ткань шеи и туловища , хотя большая часть кожи происходит из латеральной пластинки мезодермы . [2]
Миотом
[ редактировать ]Миотом — это часть сомита, образующая мышцы животного. [2] Каждый миотом делится на эпаксиальную часть ( эпимер ) сзади и гипаксиальную часть ( гипомер ) спереди. [2] Миобласты гипаксиального отдела образуют мышцы грудной и передней брюшной стенок. Эпаксиальная мышечная масса теряет сегментарный характер и образует мышцы-разгибатели шеи и туловища млекопитающих.
У рыб, саламандр, червяг и рептилий мускулатура тела остается сегментированной, как у эмбриона, хотя часто становится складчатой и перекрывающейся, с эпаксиальными и гипаксиальными массами, разделенными на несколько отдельных мышечных групп. [ нужна ссылка ]
Склеротом
[ редактировать ]Склеротом ) (или пластинка кожи образует позвонки , реберный хрящ и часть затылочной кости; миотом образует мускулатуру спины, ребер и конечностей; синдетом образует сухожилия, а дерматом образует кожу на спине. Кроме того, сомиты определяют пути миграции клеток нервного гребня и аксонов спинномозговых нервов . Из своего первоначального местоположения внутри сомита клетки склеротома мигрируют медиально к хорде . Эти клетки встречаются с клетками склеротома с другой стороны, образуя тело позвонка. Нижняя половина одного склеротома сливается с верхней половиной соседнего, образуя тела каждого позвонка. [11] Из тела позвонка клетки склеротома движутся дорсально и окружают развивающийся спинной мозг , образуя дугу позвонка. Другие клетки перемещаются дистально к реберным отросткам грудных позвонков, образуя ребра. [11]
У членистоногих
[ редактировать ]В развитии ракообразных сомит представляет собой сегмент гипотетического примитивного строения тела ракообразных. У современных ракообразных некоторые из этих сомитов могут быть слиты. [ нужна ссылка ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Кушьери, Альфред. «Третья неделя жизни» . Мальтийский университет . Проверено 13 октября 2007 г.
- ^ Jump up to: а б с д и ж Ларсен, Уильям Дж. (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. стр. 53–86. ISBN 978-0-443-06583-5 .
- ^ «Метамер» . Словарь и тезаурус-Мерриам-Вебстер онлайн . Мерриам-Вебстер. 2012 . Проверено 11 декабря 2012 г.
- ^ Jump up to: а б с д Гилберт, Сан-Франциско (2010). Биология развития (9-е изд.). Sinauer Associates, Inc., стр. 413–415 . ISBN 978-0-87893-384-6 .
- ^ Бейкер, RE ; Шнелл, С.; Майни, ПК (2006). «Часы и механизм волнового фронта для формирования сомитов» . Биология развития . 293 (1): 116–126. дои : 10.1016/j.ydbio.2006.01.018 . ПМИД 16546158 .
- ^ Гольдбетер, А.; Пурке, О. (2008). «Моделирование часов сегментации как сети связанных колебаний в сигнальных путях Notch, Wnt и FGF» (PDF) . Журнал теоретической биологии . 252 (3): 574–585. Бибкод : 2008JThBi.252..574G . дои : 10.1016/j.jtbi.2008.01.006 . PMID 18308339 – через Свободный университет Брюсселя .
- ^ Вахи, Кану (2016). «Множество ролей передачи сигналов Notch во время сомитогенеза позвоночных». Семинары по клеточной биологии и биологии развития . 49 : 68–75. дои : 10.1016/j.semcdb.2014.11.010 . ПМИД 25483003 . S2CID 10822545 .
- ^ Гомес, К; и др. (2008). «Контроль количества сегментов у эмбрионов позвоночных». Природа . 454 (7202): 335–339. Бибкод : 2008Natur.454..335G . дои : 10.1038/nature07020 . ПМИД 18563087 . S2CID 4373389 .
- ^ Брент А.Е., Швейцер Р., Табин С.Дж. (апрель 2003 г.). «Сомитический отсек предшественников сухожилий» . Клетка . 113 (2): 235–48. дои : 10.1016/S0092-8674(03)00268-X . ПМИД 12705871 . S2CID 16291509 .
- ^ «Изображения эмбриона» . Медицинский факультет Университета Северной Каролины . Проверено 19 октября 2007 г.
- ^ Jump up to: а б Уокер, Уоррен Ф. младший (1987) Функциональная анатомия позвоночных, Сан-Франциско: Издательство Saunders College.
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|