Агробактерия тумефациенс
| Агробактерия тумефациенс | |
|---|---|
 | |
| Agrobacterium tumefaciens прикрепляется к клетке моркови | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Псевдомонадота |
| Сорт: | Альфапротеобактерии |
| Заказ: | Гифомикробиалы |
| Семья: | Ризобиевые |
| Род: | Агробактерия |
| Разновидность: | А. тумефациенс |
| Биномиальное имя | |
| Агробактерия тумефациенс (Смит и Таунсенд, 1907 г.) Конн, 1942 г. (утвержденные списки, 1980 г.) | |
| Тип штамма | |
| АТСС 4720 [1] [2] [а] | |
| Синонимы [7] [8] [2] | |
Гомотипические синонимы
Гетеротипические синонимы Agrobacterium radiobacter (Beijerinck and van Delden 1902) Conn 1942 (Утвержденные списки 1980 г.) НЕ являются синонимами. [2] Эти два раньше были синонимами [6] на основании необоснованного изменения типовых штаммов в утвержденных списках 1980 г., которые теперь отменены. [2] | |
Агробактерия тумефациенс [3] [2] является возбудителем корончатой желчнокаменной болезни (образования опухолей ) у более чем 140 видов эвдикотов . Это палочковидная грамотрицательная почвенная бактерия . [4] Симптомы вызваны вставкой небольшого сегмента ДНК (известного как Т-ДНК , что означает «транспортная ДНК», не путать с тРНК, которая переносит аминокислоты во время синтеза белка) из плазмиды в растительную клетку. [9] который включен в полуслучайное место в геном растения . Геномы растений можно сконструировать с помощью Agrobacterium для доставки последовательностей, содержащихся в бинарных векторах Т-ДНК .
Agrobacterium tumefaciens — альфапротеобактерия семейства Rhizobiaceae , в состав которого входят азотфиксирующие бобовых симбионты . В отличие от азотфиксирующих симбионтов, опухолеобразующие виды Agrobacterium являются патогенными и не приносят пользы растению. Большое разнообразие растений, поражаемых Agrobacterium, вызывает серьезную озабоченность в сельском хозяйстве. [10]
С экономической точки зрения A. tumefaciens является серьезным возбудителем грецких орехов , виноградной лозы , косточковых плодов , ореховых деревьев, сахарной свеклы , хрена и ревеня , а стойкий характер опухолей или галлов, вызванных этой болезнью, делает ее особенно вредной для многолетних культур. . [11]
Agrobacterium tumefaciens оптимально растет при температуре 28 ° C (82 ° F). Время удвоения может варьироваться от 2,5 до 4 часов в зависимости от среды, формата культуры и уровня аэрации. [12] При температуре выше 30 ° C (86 ° F) A. tumefaciens начинает испытывать тепловой шок, который может привести к ошибкам в делении клеток. [12]
Спряжение
[ редактировать ]Чтобы быть вирулентной , бактерия содержит индуцирующую опухоль плазмиду (Ti-плазмиду или pTi) длиной 200 т.п.н. , которая содержит Т-ДНК и все гены, необходимые для ее переноса в растительную клетку. [13] Многие штаммы A. tumefaciens не содержат pTi.
Поскольку плазмида Ti необходима для возникновения заболевания, происходят события предварительного проникновения в ризосферу, способствующие бактериальной конъюгации - обмену плазмидами между бактериями. В присутствии опинов N- A. tumefaciens вырабатывает диффузионный сигнал конъюгации, называемый ( 3-оксооктаноил)-L-гомосерин лактоном (3OC8HSL) или аутоиндуктором Agrobacterium . [14] Это активирует фактор транскрипции TraR, положительно регулирующий транскрипцию генов, необходимых для конъюгации. [15]
Способы заражения
[ редактировать ]Agrobacterium tumefaciens заражает растение через свою Ti-плазмиду. Плазмида Ti интегрирует сегмент своей ДНК, известный как Т-ДНК, в хромосомную ДНК клеток растения-хозяина. У A. tumefaciens есть жгутики , которые позволяют ему плавать через почву к фотоассимилятам , которые накапливаются в ризосфере вокруг корней. Некоторые штаммы могут хемотаксически перемещаться в сторону химических экссудатов растений, таких как ацетосирингон и сахара, что указывает на наличие раны в растении, через которую могут проникнуть бактерии. Фенольные соединения распознаются белком VirA , трансмембранным белком, кодируемым геном virA на плазмиде Ti. Сахара распознаются белком chvE, белком, кодируемым хромосомным геном, расположенным в периплазматическом пространстве. [16]
Для индукции опухоли необходимо наличие как минимум 25 генов vir на плазмиде Ti. [17] Помимо своей роли восприятия, virA и chvE индуцируют другие гены vir . Белок VirA обладает аутокиназной активностью : он фосфорилирует себя по остатку гистидина. Затем белок VirA фосфорилирует белок VirG по его аспартатному остатку. Белок virG представляет собой цитоплазматический белок, продуцируемый плазмидным геном virG Ti. Это транскрипционный фактор , индуцирующий транскрипцию vir оперонов . Белок ChvE регулирует второй механизм активации генов vir . Повышает чувствительность белка VirA к фенольным соединениям. [16]
Прикрепление — это двухэтапный процесс. После первоначального слабого и обратимого прикрепления бактерии синтезируют целлюлозные фибриллы , которые прикрепляют их к поврежденной растительной клетке, к которой они были привлечены. В этом процессе участвуют четыре основных гена: chvA , chvB , pscA и att . Продукты первых трех генов, по-видимому, участвуют в собственно синтезе фибрилл целлюлозы. Эти фибриллы также связывают бактерии друг с другом, помогая формировать микроколонии . [ нужна ссылка ]
VirC, наиболее важный вирулентный белок, является необходимым шагом в рекомбинации незаконной реколонизации. Он выбирает участок ДНК растения-хозяина, который будет заменен, и разрезает эту цепь ДНК. [ нужна ссылка ]
После образования фибрилл целлюлозы вырабатывается кальций-зависимый внешней мембраны белок , называемый рикадгезин, который также помогает бактериям прикрепляться к клеточной стенке. Гомологи этого белка можно найти и у других ризобий. В настоящее время имеется несколько отчетов по стандартизации протокола Agrobacterium трансформации, опосредованной . Влияние различных параметров, таких как время заражения, ацетосирингон, DTT и цистеин, было изучено на соевых бобах ( Glycine max ). [18]
Возможные растительные соединения, которые инициируют агробактериями растительных клеток: заражение [19]
- Ацетосирингон и другие фенольные соединения
- альфа- гидроксиацетосирингон
- Катехин
- Феруловая кислота
- Галловая кислота
- п-гидроксибензойная кислота
- Протокатеховая кислота
- пирогалловая кислота
- Резорциловая кислота
- Синапиновая кислота
- Сиринговая кислота
- Ванилин
Формирование Т-пилуса
[ редактировать ]Для переноса Т-ДНК в клетку растительную A. tumefaciens использует механизм секреции типа IV, включающий выработку Т- пилюсов . При обнаружении ацетосирингона и других веществ событие сигнальной трансдукции активирует экспрессию 11 генов оперона VirB , которые отвечают за формирование Т-пилусов.
Пропилин образуется первым. Это полипептид из 121 аминокислоты, который требует обработки путем удаления 47 остатков с образованием субъединицы Т-пилюс. Считалось, что субъединица приобретает циркулярную форму за счет образования пептидной связи между двумя концами полипептида. Однако структура Т-пилуса с высоким разрешением не выявила циклизации пилина, при этом общая организация субъединиц пилина очень похожа на организацию других конъюгативных пилей, таких как F-пилус. [20]
Продукты других генов VirB используются для переноса субъединиц через плазматическую мембрану . Исследования двухгибридных дрожжей свидетельствуют о том, что VirB6, VirB7, VirB8, VirB9 и VirB10 могут кодировать компоненты транспортера. для Также потребуется АТФаза активного транспорта субъединиц.
Перенос Т-ДНК в растительную клетку
[ редактировать ]- Агробактериальная клетка
- Агробактериальная хромосома
- Ti-плазмида (а. Т-ДНК, б. гены vir, c. начало репликации, d. катаболизм опинов)
- Растительная клетка
- Растительные митохондрии
- Растительный хлоропласт
- Ядро растения
- Распознавание VirA
- VirA фосфорилирует VirG
- VirG вызывает транскрипцию генов Vir
- Гены Vir вырезают Т-ДНК и образуют нуклеопротеиновый комплекс («Т-комплекс»).
- Т-комплекс проникает в цитоплазму растения через Т-пилус.
- Т-ДНК проникает в ядро растения через ядерную пору.
- Т-ДНК достигает интеграции
Т-ДНК необходимо вырезать из кольцевой плазмиды. Обычно это осуществляется генами Vir внутри хелперной плазмиды. [21] Комплекс VirD1/D2 разрывает ДНК на левой и правой пограничных последовательностях. Белок VirD2 ковалентно присоединен к 5'-концу. VirD2 содержит мотив , который приводит к нацеливанию нуклеопротеинового комплекса на систему секреции типа IV (T4SS). Структура Т-пилуса показала, что центральный канал пилуса слишком узок, чтобы обеспечить перенос свернутого VirD2, что позволяет предположить, что VirD2 должен быть частично развернут во время процесса конъюгации. [20]
В цитоплазме клетки-реципиента комплекс Т-ДНК покрывается белками VirE2, которые экспортируются через T4SS независимо от комплекса Т-ДНК. Сигналы ядерной локализации , или NLS, расположенные на VirE2 и VirD2, распознаются белком импортина альфа, который затем связывается с импортином бета и комплексом ядерных пор для переноса Т-ДНК в ядро . VIP1 также, по-видимому, является важным белком в этом процессе, возможно, действуя как адаптер для доставки VirE2 к импортину. Оказавшись внутри ядра, VIP2 может нацелить Т-ДНК на области хроматина , которые активно транскрибируются, так что Т-ДНК может интегрироваться в геном хозяина.
Гены в Т-ДНК
[ редактировать ]Гормоны
[ редактировать ]Чтобы вызвать образование галла , Т-ДНК кодирует гены, отвечающие за выработку ауксина или индол-3-уксусной кислоты по пути IAM. Этот путь биосинтеза не используется во многих растениях для производства ауксина, поэтому это означает, что у растения нет молекулярных средств его регулирования, и ауксин будет производиться конститутивно. гены продукции цитокининов Также экспрессируются . Это стимулирует пролиферацию клеток и образование галлов.
Мнения
[ редактировать ]Т-ДНК содержит гены, кодирующие ферменты , которые заставляют растение создавать специализированные производные аминокислот , которые бактерии могут метаболизировать , называемые опины . [22] Опины — это класс химических веществ, которые служат источником азота для A. tumefaciens , но не для большинства других организмов. Конкретным типом опина, вырабатываемого растениями, инфицированными A. tumefaciens C58, является нопалин . [23]
нопалинового типа Две Ti-плазмиды , pTi-SAKURA и pTiC58, были полностью секвенированы. « A. fabrum » C58, первый полностью секвенированный патовар , был впервые выделен из галла кроны вишневого дерева. Геном одновременно секвенировали Goodner et al. [24] и Вуд и др. [25] в 2001 году. Геном A. tumefaciens C58 состоит из кольцевой хромосомы, двух плазмид и линейной хромосомы . Наличие ковалентно связанной кольцевой хромосомы характерно для бактерий, за некоторыми исключениями. Однако наличие как одной кольцевой хромосомы, так и одной линейной хромосомы уникально для группы этого рода. Две плазмиды — pTiC58, ответственная за процессы, участвующие в вирулентности , и pAtC58, [б] когда-то получивший название «загадочная» плазмида. [24] [25]
Было показано, что плазмида pAtC58 участвует в метаболизме опинов и конъюгирует с другими бактериями в отсутствие плазмиды pTiC58. [26] Если Ti-плазмиду удалить, роста опухоли, на основании которого классифицируют этот вид бактерий, не происходит.
Биотехнологическое использование
[ редактировать ]Конференция Асиломар в 1975 году установила широко распространенное мнение о том, что рекомбинантные методы недостаточно изучены и требуют строгого контроля. [27] [28] Возможности передачи ДНК Agrobacterium широко изучались в биотехнологии как средство внедрения чужеродных генов в растения. Вскоре после конференции Asilomar Марк Ван Монтегю и Джефф Шелл открыли механизм переноса генов между Agrobacterium и растениями, что привело к разработке методов превращения бактерии в эффективную систему доставки для генной инженерии растений. [29] Плазмидная Т-ДНК, перенесенная в растение, является идеальным средством генной инженерии. [30] Это делается путем клонирования желаемой последовательности гена в бинарные векторы Т-ДНК , которые будут использоваться для доставки интересующей последовательности в эукариотические клетки. Этот процесс был осуществлен с использованием гена люциферазы светлячка для получения светящихся растений. [31] Эта люминесценция оказалась полезным устройством при изучении функции хлоропластов растений и в качестве репортерного гена . [31] Также возможно трансформировать Arabidopsis thaliana , погружая цветы в бульон Agrobacterium : полученные семена будут трансгенными . В лабораторных условиях Т-ДНК также была перенесена в клетки человека, что продемонстрировало разнообразие применения вставки. [32]
Механизм, с помощью которого Agrobacterium вводит материалы в клетку-хозяина, представляет собой систему секреции типа IV , которая очень похожа на механизмы, используемые патогенами для внедрения материалов (обычно белков ) в клетки человека путем секреции типа III. Он также использует тип передачи сигналов, консервативный у многих грамотрицательных бактерий, называемый ощущением кворума . [ нужна ссылка ] Это делает Agrobacterium важной темой медицинских исследований. также [ нужна ссылка ]
Естественная генетическая трансформация
[ редактировать ]Естественная генетическая трансформация бактерий представляет собой половой процесс , включающий перенос ДНК из одной клетки в другую через промежуточную среду и интеграцию донорной последовательности в геном реципиента путем гомологичной рекомбинации . A. tumefaciens может претерпевать естественную трансформацию в почве без какой-либо специальной физической или химической обработки. [33]
Цикл заболевания
[ редактировать ]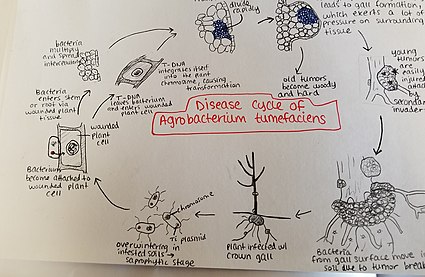
Agrobacterium tumefaciens зимует в зараженных почвах. Виды Agrobacterium ведут преимущественно сапрофитный образ жизни, поэтому даже виды, паразитирующие на растениях этого рода, обычно выживают в почве в течение длительных периодов времени, даже без присутствия растения-хозяина. [34] Однако при наличии растения-хозяина бактерии проникают в ткань растения через недавние раны или естественные отверстия корней или стеблей у земли. Эти раны могут быть вызваны культурными практиками, прививкой, насекомыми и т. д. Как только бактерии проникают в растение, они возникают межклеточно и стимулируют пролиферацию окружающих тканей за счет клеточной трансформации. Agrobacterium осуществляет этот контроль, встраивая плазмиду Т-ДНК в геном растения. Более подробную информацию о процессе внедрения плазмидной ДНК в геном хозяина см. выше. Чрезмерный разрастание растительной ткани приводит к образованию галлов на стебле и корнях. Эти опухоли оказывают значительное давление на окружающую растительную ткань, что приводит к ее раздавливанию и/или деформации. Разрушение сосудов приводит к уменьшению тока воды в ксилеме. Молодые опухоли мягкие и поэтому уязвимы для вторичной инвазии насекомыми и сапрофитными микроорганизмами. Эта вторичная инвазия вызывает разрушение слоев периферических клеток, а также изменение цвета опухоли из-за распада. Разрушение мягких тканей приводит к высвобождению Agrobacterium tumefaciens попадает в почву, позволяя возобновить процесс заболевания на новом растении-хозяине. [35]
Управление болезнями
[ редактировать ]Заболевание корончатого желчного пузыря, вызываемое Agrobacterium tumefaciens, можно контролировать с помощью различных методов. Лучший способ борьбы с этим заболеванием — принять профилактические меры, такие как стерилизация инструментов для обрезки, чтобы избежать заражения новых растений. Ценной практикой также является проведение обязательных проверок питомников и отказ от зараженных растений, а также отказ от посадки восприимчивых растений на зараженных полях. Для предотвращения заболеваний важно избегать повреждения кроны/корней растений во время выращивания. В методах садоводства, при которых несколько растений растут как одно, например, окулировка и прививка. [36] эти приемы приводят к повреждению растений. Раны являются основным местом проникновения бактерий в растение-хозяин. Поэтому целесообразно выполнять эти методы в то время года, когда агробактерии не активны. Борьба с насекомыми, грызущими корни, также полезна для снижения уровня инфекции, поскольку эти насекомые вызывают раны (так называемые бактериальные входные пути) в корнях растений. [35] Зараженный растительный материал рекомендуется сжигать, а не помещать в компостную кучу, поскольку бактерии способны жить в почве в течение многих лет. [37]
Для борьбы с этим заболеванием также используются методы биологического контроля. В 1970-е и 1980-е годы обычной практикой обработки проросших семян, сеянцев и подвоев было замачивание их в суспензии К84. K84 состоит из A. radiobacter, вида, родственного A. tumefaciens , но не патогенного. K84 производит бактериоцин (агроцин 84), который является антибиотиком, специфичным против родственных бактерий, включая A. tumefaciens . Этот метод, который оказался успешным в борьбе с заболеванием в коммерческих масштабах, имел риск передачи гена устойчивости K84 патогенным агробактериям. Таким образом, в 1990-е годы было создано использование генно-инженерного штамма К84, известного как К-1026. Этот штамм столь же успешен в борьбе с корончатым галлом, как и K84, без учета переноса генов устойчивости. [38]
Среда
[ редактировать ]
Хозяин, окружающая среда и патоген являются чрезвычайно важными понятиями в отношении патологии растений. Агробактерии имеют самый широкий круг хозяев среди всех патогенов растений. [39] поэтому основным фактором, который следует учитывать в случае корончатого галла, является окружающая среда. Существуют различные условия и факторы, которые создают благоприятную среду для A. tumefaciens при заражении различных хозяев. Бактерия не может проникнуть в растение-хозяин без точки входа, например раны. Факторы, приводящие к повреждению растений, включают агротехнику, прививку, травмы от замерзания, ростовые трещины, почвенных насекомых и других животных в окружающей среде, наносящих вред растению. Следовательно, в исключительно суровые зимы обычно увеличивается заболеваемость коронковым галлом из-за повреждений, связанных с погодными условиями. [40] Наряду с этим существуют способы опосредования заражения растения-хозяина. Например, нематоды могут действовать как переносчики агробактерий в корни растений. В частности, корневые паразитические нематоды повреждают растительную клетку, создавая рану для проникновения бактерий. [41] Наконец, температура является фактором при рассмотрении A. tumefaciens инфекции . Оптимальная температура для образования корончатого галла, вызванного этой бактерией, составляет 22 ° C (72 ° F) из-за термочувствительности переноса Т-ДНК. Образование опухолей значительно снижается в условиях более высоких температур. [42]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Веласкес Э., Флорес-Феликс Х.Д., Санчес-Хуанес Ф., Игуаль Х.М., Пейкс А. (2020). «Штамм АТСС 4720 Т является подлинным типовым штаммом Agrobacterium tumefaciens , который не является более поздним гетеротипическим синонимом Agrobacterium radiobacter » . Int J Syst Evol Microbiol . 70 (9): 5172–5176. doi : 10.1099/ijsem.0.004443 . PMID 32915125 .
- ^ Jump up to: а б с д и Арахал Д.Р., Булл К.Т., Буссе Х.Дж., Кристенсен Х., Чувочина М., Дедыш С.Н., Фурнье П.Е., Константинидис К.Т., Паркер К.Т., Росселло-Мора Р., Вентоза А., Гёкер М. (27 апреля 2023 г.). «Судебные заключения 123–127». Международный журнал систематической и эволюционной микробиологии . 72 (12). дои : 10.1099/ijsem.0.005708 . hdl : 10261/295959 . ПМИД 36748499 . [Примечание: Судебное заключение 127 определяет штамм ATCC 4720 как типовой штамм Agrobacterium tumefaciens.]
- ^ Jump up to: а б с «Браузер таксономии ( Agrobacterium tumefaciens )» . Национальный центр биотехнологической информации . Проверено 7 января 2024 г.
- ^ Jump up to: а б Смит Э.Ф., Таунсенд, штат Колорадо (апрель 1907 г.). «Растение-опухоль бактериального происхождения» . Наука . 25 (643): 671–3. Бибкод : 1907Sci....25..671S . дои : 10.1126/science.25.643.671 . ПМИД 17746161 .
- ^ Деламута-младший, Шерер А.Дж., Рибейро Р.А., Хунгрия М. (2020). «Генетическое разнообразие видов Agrobacterium, выделенных из клубеньков фасоли обыкновенной и сои в Бразилии, Мексике, Эквадоре и Мозамбике, и описание нового вида Agrobacterium fabacearum sp. nov» . Международный журнал систематической и эволюционной микробиологии . 70 (7): 4233–4244. дои : 10.1099/ijsem.0.004278 . ISSN 1466-5034 .
- ^ Тиндалл Б.Дж. и др. (Судебная комиссия) (2014). «Судебное заключение № 94: Agrobacterium radiobacter (Beijerinck and van Delden 1902) Conn 1942 имеет приоритет над Agrobacterium tumefaciens (Smith and Townsend 1907) Conn 1942, когда они рассматриваются как представители одного и того же вида на основе принципа приоритета и правила. 23а, примечание 1 применительно к соответствующим видовым эпитетам» . Int J Syst Evol Microbiol . 64 (Часть 10): 3590–3592. дои : 10.1099/ijs.0.069203-0 . ПМИД 25288664 .
- ^ Бьюкенен Р.Э. (1965). «Предложение об отказе от родового названия Polymonas Lieske 1928» . Международный бюллетень бактериологической номенклатуры и таксономии . 15 (1): 43–44. дои : 10.1099/00207713-15-1-43 .
- ^ Савада Х., Иеки Х., Ояизу Х., Мацумото С. (1993). «Предложение об отказе от Agrobacterium tumefaciens и пересмотренных описаниях рода Agrobacterium и Agrobacterium radiobacter и Agrobacterium rhizogenes » . Int J Syst Bacteriol . 43 (4): 694–702. дои : 10.1099/00207713-43-4-694 . ПМИД 8240952 .
- ^ Чилтон, доктор медицинских наук, Драммонд М.Х., Мерло DJ, Скиаки Д., Монтойя А.Л., Гордон М.П., Нестер Э.В. (июнь 1977 г.). «Стабильное включение плазмидной ДНК в клетки высших растений: молекулярная основа онкогенеза корончатого галла». Клетка . 11 (2): 263–271. дои : 10.1016/0092-8674(77)90043-5 . ISSN 0092-8674 . ПМИД 890735 . S2CID 7533482 .
- ^ Мур Л.В., Чилтон В.С., Кэнфилд М.Л. (январь 1997 г.). «Разнообразие опинов и бактерий, катаболизирующих опины, выделенных из естественных опухолей корончатого желчного пузыря» . Прикладная и экологическая микробиология . 63 (1): 201–7. Бибкод : 1997ApEnM..63..201M . дои : 10.1128/АЕМ.63.1.201-207.1997 . ПМК 1389099 . ПМИД 16535484 .
- ^ «Коронные галлы» . Ботанический сад Миссури . Проверено 2 декабря 2019 г.
- ^ Jump up to: а б Мортон Э.Р., Фукуа С. (февраль 2012 г.). «Лабораторное обеспечение агробактерий » . Современные протоколы в микробиологии . Глава 1: Unit3D.1. дои : 10.1002/9780471729259.mc03d01s24 . ISBN 978-0471729259 . ПМК 3350319 . ПМИД 22307549 .
- ^ Гордон Дж. Э., Кристи П. Дж. (декабрь 2014 г.). « Плазмиды Agrobacterium Ti» . Микробиологический спектр . 2 (6). doi : 10.1128/microbiolspec.PLAS-0010-2013 . ПМК 4292801 . ПМИД 25593788 .
- ^ Огер П., Фарранд С.К. (2002). «Два опина контролируют супружеский перенос плазмиды Agrobacterium путем регулирования экспрессии отдельных копий гена активатора восприятия кворума traR» . Журнал бактериологии . 184 (4): 1121–1131. дои : 10.1128/jb.184.4.1121-1131.2002 . ISSN 0021-9193 . ПМК 134798 . ПМИД 11807073 .
- ^ Чжан Х.Б., Ван Л.Х., Чжан Л.Х. (2002). «Генетический контроль оборота сигнала кворума у Agrobacterium tumefaciens» . Труды Национальной академии наук . 99 (7): 4638–4643. Бибкод : 2002PNAS...99.4638Z . дои : 10.1073/pnas.022056699 . ISSN 0027-8424 . ПМК 123700 . ПМИД 11930013 .
- ^ Jump up to: а б Гельвин С.Б. (март 2003 г.). « Трансформация растений, опосредованная агробактериями : биология, лежащая в основе инструмента «генной игры» . Обзоры микробиологии и молекулярной биологии . 67 (1): 16–37, оглавление. дои : 10.1128/ммбр.67.1.16-37.2003 . ПМК 150518 . ПМИД 12626681 .
- ^ Винанс СК (1992). «Двусторонняя химическая передача сигналов во взаимодействиях агробактерий и растений» . Микробиологические обзоры . 56 (1): 12–31. дои : 10.1128/г.56.1.12-31.1992 . ISSN 0146-0749 . ПМЦ 372852 . ПМИД 1579105 .
- ^ Барате П.Л., Кумар Р.Р., Вагмаре С.Г., Павар К.Р., Табе Р.Х. (2018). «Влияние различных параметров на трансформацию, опосредованную Agrobacterium, в Glycine max » . Международный журнал перспективных биологических исследований . 8 (1): 99–105.
- ^ Патент США 6483013 «Способ трансформации хлопка, опосредованной агробактериями», опубликован 19 ноября 2002 г., передан компании Bayer BioScience NV (BE).
- ^ Jump up to: а б Бельтран Л.К., Цвиркайте-Крупович В., Миллер Дж., Ван Ф., Кройцбергер М.А., Патковски Дж.Б., Коста Т.Р., Схоутен С., Левенталь И., Контичелло В.П., Эгельман Э.Х., Крупович М. (07.02.2023). «Аппарат для импорта ДНК архей гомологичен бактериальному аппарату конъюгации» . Природные коммуникации . 14 (1): 666. Бибкод : 2023NatCo..14..666B . дои : 10.1038/s41467-023-36349-8 . ISSN 2041-1723 . ПМЦ 9905601 . ПМИД 36750723 .
- ^ Кремер Т. «Руководство по бинарным векторам Т-ДНК в трансформации растений» . ГолдБио . Проверено 9 января 2024 г.
- ^ Зупан Дж., Мут Т.Р., Дрейпер О., Замбриски П. (июль 2000 г.). «Перенос ДНК из Agrobacterium tumefaciens в растения: праздник фундаментальных открытий». Заводской журнал . 23 (1): 11–28. дои : 10.1046/j.1365-313x.2000.00808.x . ПМИД 10929098 .
- ^ Эскобар М.А., Чивероло Э.Л., Полито В.С., Пинни К.А., Дандекар А.М. (январь 2003 г.). «Характеристика трансгенных растений с молчанием онкогенов: значение для биологии Agrobacterium и посттранскрипционного подавления генов» . Молекулярная патология растений . 4 (1): 57–65. дои : 10.1046/j.1364-3703.2003.00148.x . ISSN 1464-6722 . ПМИД 20569363 .
- ^ Jump up to: а б Гуднер Б., Хинкль Г., Гаттунг С., Миллер Н., Бланшар М., Куролло Б. и др. (декабрь 2001 г.). «Последовательность генома патогена растений и биотехнологического агента Agrobacterium tumefaciens C58». Наука . 294 (5550): 2323–2328. Бибкод : 2001Sci...294.2323G . дои : 10.1126/science.1066803 . ISSN 0036-8075 . ПМИД 11743194 . S2CID 86255214 .
- ^ Jump up to: а б Вуд Д.В., Сетубал Дж.К., Каул Р., Монкс Д.Э., Китадзима Дж.П., Окура В.К. и др. (декабрь 2001 г.). «Геном природного генного инженера Agrobacterium tumefaciens C58». Наука . 294 (5550): 2317–23. Бибкод : 2001Sci...294.2317W . CiteSeerX 10.1.1.7.9501 . дои : 10.1126/science.1066804 . ISSN 0036-8075 . ПМИД 11743193 . S2CID 2761564 .
- ^ Водекен-Дрансарт В., Пети А., Чилтон В.С., Дессау И. (1998). «Загадочная плазмида Agrobacterium tumefaciens коинтегрируется с плазмидой Ti и участвует в деградации опина» . Молекулярные растительно-микробные взаимодействия . 11 (7): 583–591. дои : 10.1094/mpmi.1998.11.7.583 .
- ^ Берг П., Балтимор Д., Бреннер С., Роблин Р.О., Сингер М.Ф. (6 июня 1975 г.). «Асиломарская конференция по рекомбинантным молекулам ДНК» . Наука . 188 (4192): 991–994. Бибкод : 1975Sci...188..991B . дои : 10.1126/science.1056638 . ISSN 0036-8075 . ПМИД 1056638 .
- ^ Национальный исследовательский совет, Совет по сельскому хозяйству и природным ресурсам, Комитет по генетически модифицированным растениям, защищенным от вредителей (2000). Генетически модифицированные растения, защищенные от вредителей: наука и регулирование . Вашингтон, округ Колумбия : Издательство национальных академий . стр. XXIII, 263. doi : 10.17226/9795 . ISBN 978-0-309-06930-4 . OCLC 894124744 . ПМИД 25032472 .
- ^ Шелл Дж., Ван Монтегю М. (1977). «Ти-плазмида Agrobacterium Tumefaciens, природный вектор для введения генов NIF в растения?». В Hollaender A, Burris RH, Day PR, Hardy RW (ред.). Генная инженерия для фиксации азота . Фундаментальные науки о жизни. Том. 9. стр. 159–79. дои : 10.1007/978-1-4684-0880-5_12 . ISBN 978-1-4684-0882-9 . ПМИД 336023 .
- ^ Замбриски П., Йоос Х., Дженетелло С., Лиманс Дж., Монтегю М.В., Шелл Дж. (1983). «Ти-плазмидный вектор для введения ДНК в растительные клетки без изменения их нормальной регенерационной способности» . Журнал ЭМБО . 2 (12): 2143–50. дои : 10.1002/j.1460-2075.1983.tb01715.x . ПМК 555426 . ПМИД 16453482 .
- ^ Jump up to: а б Рут М (1988). «Светящаяся в темноте биотехнология» . Бионаука . 38 (11): 745–747. дои : 10.2307/1310781 . JSTOR 1310781 .
- ^ Куник Т., Цфира Т., Капульник Ю., Гафни Ю., Дингуолл С., Цитовский В. (февраль 2001 г.). «Генетическая трансформация клеток HeLa агробактериями » . Труды Национальной академии наук Соединенных Штатов Америки . 98 (4): 1871–6. Бибкод : 2001PNAS...98.1871K . дои : 10.1073/pnas.041327598 . ЧВК 29349 . ПМИД 11172043 .
- ^ Деманеш С., Кей Э., Гурбьер Ф., Симоне П. (июнь 2001 г.). «Естественная трансформация Pseudomonas fluorescens и Agrobacterium tumefaciens в почве» . Прикладная и экологическая микробиология . 67 (6): 2617–21. Бибкод : 2001ApEnM..67.2617D . дои : 10.1128/АЕМ.67.6.2617-2621.2001 . ПМК 92915 . ПМИД 11375171 .
- ^ Шрот М.Н., Вайнхольд А.Р., Маккейн А.Х. (март 1971 г.). «Биология и борьба с Agrobacterium tumefaciens » . Хилгардия . 40 (15): 537–552. дои : 10.3733/hilg.v40n15p537 .
- ^ Jump up to: а б Агриос Г.Н. (2005). Патология растений (5-е изд.). Амстердам : Elsevier Academic Press . дои : 10.1016/C2009-0-02037-6 . ISBN 9780120445653 . OCLC 55488155 .
- ^ Бильдербек Т., Бир Р.Э., Ранни Т.Г. (30 июня 2014 г.). «Прививка и окулировка саженцев» . Публикации по расширению штата Северная Каролина . Проверено 12 декабря 2017 г.
- ^ Кеттер Р., Грабовски М. «Короночная желчь» . Расширение Университета Миннесоты . Архивировано из оригинала 16 октября 2017 года . Проверено 15 октября 2017 г.
- ^ Райдер М.Х., Джонс Д.А. (1 октября 1991 г.). «Биологический контроль корончатого галла с использованием штаммов агробактерий К84 и К1026». Функциональная биология растений . 18 (5): 571–579. дои : 10.1071/pp9910571 .
- ^ Эллис Массачусетс (15 апреля 2016 г.). «Бактериальный коронковый галл плодовых культур» . Огайолин . Расширение Университета штата Огайо . Проверено 20 октября 2017 г.
- ^ «Коронная желчь – растущая обеспокоенность виноградников» . Расширение штата Пенсильвания . 19 октября 2017 года. Архивировано из оригинала 20 октября 2017 года . Проверено 20 октября 2017 г.
- ^ Карими М., Ван Монтегю М., Гейсен Г. (ноябрь 2000 г.). «Нематоды как переносчики внедрения агробактерий в корни растений» . Молекулярная патология растений . 1 (6): 383–7. дои : 10.1046/j.1364-3703.2000.00043.x . ПМИД 20572986 . S2CID 35932276 .
- ^ Диллен В., Де Клерек Дж., Капила Дж., Ван Монтегю З.М., Ангенон Дж. (1 декабря 1997 г.). «Влияние температуры на Agrobacterium tumefaciens перенос генов растениям, опосредованный » . Заводской журнал . 12 (6): 1459–1463. дои : 10.1046/j.1365-313x.1997.12061459.x .
Дальнейшее чтение
[ редактировать ]- Дикинсон М (2003). Молекулярная патология растений . Научные издательства БИОС .
- Лай Э.М., Кадо С.И. (август 2000 г.). «Т-пилус Agrobacterium tumefaciens ». Тенденции в микробиологии . 8 (8): 361–9. дои : 10.1016/s0966-842x(00)01802-3 . ПМИД 10920395 .
- Уорд Д.В., Зупан-младший, Замбриски ПК (январь 2002 г.). « Агробактерия VirE2 получает обработку VIP1 при импорте ядерных растений». Тенденции в науке о растениях . 7 (1): 1–3. дои : 10.1016/s1360-1385(01)02175-6 . ПМИД 11804814 .
- Вебстер Дж., Томсон Дж. (1988). «Генетический анализ штамма Agrobacterium tumefaciens , продуцирующего агроцин, активный против возбудителя биотипа 3». Молекулярная и общая генетика . 214 (1): 142–147. дои : 10.1007/BF00340192 . S2CID 180063 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Agrobacterium tumefaciens, на Викискладе?
СМИ, связанные с Agrobacterium tumefaciens, на Викискладе? - Agrobacterium fabrum Страница генома C58 — секвенирование Cereon Genomics/Университета Ричмонда
| Базы данных органов управления : Национальные |
|---|