Боковой лемниск
| Боковой лемниск | |
|---|---|
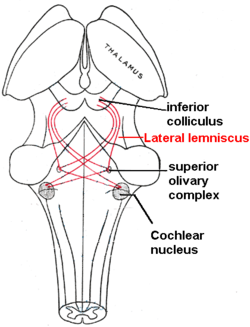 Латеральный лемниск выделен красным, так как он соединяет ядро улитки , верхнее оливковое ядро и нижний холмик . Вид сзади. | |
| Подробности | |
| Идентификаторы | |
| латинский | латеральная лемниска |
| Нейроимена | 609 |
| НейроЛекс ID | бирнлекс_976 |
| ТА98 | А14.1.05.317 А14.1.08.670 А14.1.06.204 |
| ТА2 | 5866 |
| ФМА | 72502 |
| Анатомические термины нейроанатомии | |
Латеральный лемниск — это тракт аксонов в стволе мозга , который передает информацию о звуке от ядра улитки к различным ядрам ствола мозга и, в конечном итоге, к контралатеральным нижним бугоркам среднего мозга . Внутри этих волокон расположены три отдельные, преимущественно тормозные, клеточные группы, которые поэтому называются ядрами латерального переплетения.
Соединения
[ редактировать ]На каждой из боковых лемнисков имеется по три небольших ядра:
- промежуточное ядро боковой петли (INLL)
- вентральное ядро латеральной петли (VNLL)
- дорсальное ядро латеральной петли (DNLL)
Волокна, выходящие из этих ядер ствола мозга, поднимаясь к нижнему холмику, снова присоединяются к латеральной петле. В этом смысле это не « лемниск » в истинном смысле этого слова (второй порядок, перекрещенные сенсорные аксоны), поскольку третьего (а из латеральной верхней оливы из некоторых из них исходит информация — четвертого) порядка. ядра ствола мозга.
Латеральная петля расположена в месте пересечения ядер улитки и ретикулярной формации моста (PRF). PRF спускается по ретикулоспинальному тракту, где иннервирует мотонейроны и спинальные интернейроны. Это главный слуховой тракт в стволе мозга, который соединяет верхний оливковый комплекс (SOC) с нижними холмиками (IC). Дорсальное ядро улитки (DCN) имеет вход от LL и выход к контралатеральному LL через ипсилатеральную и контралатеральную дорсальную акустическую полоску.
Две лемниски сообщаются через комиссуральные волокна Пробста.
Ядра латеральной петли
[ редактировать ]Функция комплекса ядер латеральной петли неизвестна; однако он имеет хорошее временное разрешение по сравнению с другими клетками, расположенными выше ядер улитки, и чувствителен как к временным, так и к амплитудным изменениям звука. Он также участвует в акустическом рефлексе испуга; наиболее вероятным регионом для этого является VNLL.
ДНЛЛ
[ редактировать ]Клетки DNLL лучше всего реагируют на двусторонние входные сигналы и имеют устойчивые ответы, настроенные по началу и сложности. Ядро преимущественно ГАМКергическое. [ 1 ] и проецируется билатерально к нижнему холмику и контралатерально к DNLL, при этом разные популяции клеток проецируются к каждому IC. [ 2 ]
У крыс DNLL имеет выраженную столбчатую организацию. Почти все нейроны окрашиваются на ГАМК, особенно в центральной части ядра, а оставшиеся ГАМК-отрицательные клетки перемежаются с положительными и часто окрашиваются на глицин. Видны две популяции ГАМК+ клеток: более крупные светлоокрашенные клетки, выступающие в сторону контралатерального IC, и более мелкие, более темные окрашенные клетки, выступающие ипсилатерально. ГАМКергические окончания аксонов образуют плотные группы, окруженные ГАМК-лемнискальными волокнами по всему ядру, и образуют синапсы как на соматах, так и в нейропиле. Глицинергические окончания аксонов, с другой стороны, более точно локализованы, при этом большинство нейронов-реципиентов расположены латерально в ядре. [ 3 ]
ИНЛЛ
[ редактировать ]INLL также имеет небольшую спонтанную активность и широкие кривые настройки. Временные реакции существенно отличаются от клеток VNLL.
У крыс эта структура сильно гипертрофирована, образуя выраженную выпуклость на поверхности ствола мозга. Окрашивание GAD, GABA и глицина выявляет несколько отдельных областей, которые не заметны в стандартных цитоархитектурных препаратах. Небольшое количество нейронов, окрашенных ГАМК, организовано в небольшие группы, обычно в центре ядра, тогда как нейроны, окрашенные глицином, более распространены и широко рассредоточены, с региональными концентрациями в дорсолатеральной и вентролатеральной частях ядра. Большинство ГАМК+ клеток также являются глико+. [ 1 ] [неработающая сноска]
ВНЛЛ
[ редактировать ]Звук в контралатеральном ухе вызывает самые сильные реакции в VNLL, который занимается некоторой временной обработкой. VNLL также может быть важен для декодирования IC амплитудно-модулированных звуков.
Клетки VNLL имеют небольшую спонтанную активность, широкие и умеренно сложные кривые настройки; они имеют как фазические, так и тонические реакции и участвуют во временной обработке.
У крысы VNLL состоит из двух подразделений: вентральной (столбчатой) и дорсальной (нестолбчатой) областей. Столбчатая область содержит множество глицин-положительных (0 ГАМК+) нейронов, тогда как дорсальная область содержит кластеры ГАМК+-нейронов, смешанных с гли+-клетками, причем некоторые клетки содержат и то, и другое. [ 1 ]
Вводы и выводы в ядра
[ редактировать ]В таблице ниже показано, что каждое из ядер имеет сложное расположение ипсилатеральных и контралатеральных афферентных входов и выходов: [ нужна ссылка ]
| Ядро | Вход | Выход | ||
|---|---|---|---|---|
| Контралатеральный | Ипсилатеральный | Контралатеральный | Ипсилатеральный | |
| ВНЛЛ | Передние и задние вентральные ядра улитки | Медиальное ядро трапециевидного тела | Нижний холмик ДНЛЛ | |
| ИНЛЛ | Переднее и заднее вентральное ядро улитки | Медиальное ядро трапециевидного тела | Медиальное коленчатое тело Нижний холмик | |
| ДНЛЛ | Передняя Вентральная Улитковое ядро (и двустороннее) |
Медиальное верхнее оливковое ядро Латеральное верхнее оливковое ядро (и двустороннее) |
ДНЛЛ Нижний холмик Ретикулярная формация среднего мозга Улучшенный оливковый комплекс |
Нижний холмик Медиальное коленчатое тело Ретикулярная формация среднего мозга Улучшенный оливковый комплекс |
Дополнительные изображения
[ редактировать ]-
 Рассечение ствола головного мозга. Боковой вид.
Рассечение ствола головного мозга. Боковой вид. -
 Глубокое рассечение ствола головного мозга. Боковой вид.
Глубокое рассечение ствола головного мозга. Боковой вид. -
 Глубокое рассечение ствола головного мозга. Боковой вид.
Глубокое рассечение ствола головного мозга. Боковой вид. -
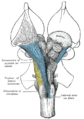 Глубокое рассечение ствола головного мозга. Вентральный вид.
Глубокое рассечение ствола головного мозга. Вентральный вид. -
 Рассечение ствола головного мозга. Дорсальный вид.
Рассечение ствола головного мозга. Дорсальный вид. -
 Корональный срез среднего мозга.
Корональный срез среднего мозга. -
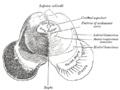 Поперечный срез среднего мозга на уровне нижних холмов.
Поперечный срез среднего мозга на уровне нижних холмов. -
 Схема, показывающая ход волокон лемниска; медиальная петля синего цвета, латеральная - красного.
Схема, показывающая ход волокон лемниска; медиальная петля синего цвета, латеральная - красного.








Ссылки
[ редактировать ]![]() Эта статья включает общедоступный текст со страницы 805 20-го издания «Анатомии Грея» (1918 г.).
Эта статья включает общедоступный текст со страницы 805 20-го издания «Анатомии Грея» (1918 г.).
- ^ Перейти обратно: а б с Адамс, Дж. К. и Э. Мугнаини (1984). «Дорсальное ядро боковой петли: ядро ГАМКергических проекционных нейронов». Brain Res Bull 13 (4): 585-90.
- ^ Баджо, В.М., М.А. Мерчан и др. (1993). «Морфология нейронов и эфферентные проекции дорсального ядра латеральной петли у крысы». J Comp Neurol 334(2): 241-62.
- ^ Винер, Дж. А., Д. Т. Ларю и др. (1995). «ГАМК и глицин в центральной слуховой системе усатой летучей мыши: структурные субстраты тормозной нейронной организации». J Comp Neurol 355(3): 317-53.