Глиптотерий
| Глиптотерий Временной диапазон: ранний плиоцен — поздний плейстоцен ( бланкан — ранчолабрей ) ( энсенадан — луханиан ) ~ | |
|---|---|
 | |
| G. texanum , Национальный музей естественной истории. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Ремень |
| Семья: | Хламифориды |
| Подсемейство: | † Глиптодонтины |
| Род: | † Глиптотерий Осборн, 1903 год. |
| Типовой вид | |
| † Glyptotherium texanum Осборн, 1903 год. | |
| Другие виды | |
| |
 | |
| Распространение глиптотерия (оранжевый) по сравнению с глиптодоном (зеленый). | |
| Синонимы | |
Синонимы G. texanum Синонимы G. cylindricum | |
Glyptotherium (от греческого «рифленый или резной зверь») — род глиптодонтов (вымершая группа крупных травоядных броненосцев ) семейства Chlamyphoridae (семейство южноамериканских броненосцев), живших с раннего плиоцена , около 3,6 миллионов лет назад. назад, к позднему плейстоцену, около 15 000 лет назад. Он имел широкое распространение, обитая в США , Мексике , Гватемале , Коста-Рике , Гондурасе , Сальвадоре , Панаме , Венесуэле и Бразилии . Род был впервые описан в 1903 году американским палеонтологом Генри Фэйрфилдом Осборном , где типовым видом был G. texanum , на основе окаменелостей, обнаруженных в плиоценовых пластах Бланкан в Ллано-Эстакадо , штат Техас , США. С тех пор окаменелости глиптотериев были обнаружены во многих других ископаемых местах, от Флориды до Колумбии. Другой вид, G. cylindricum , был назван в 1912 году охотником за ископаемыми Барнумом Брауном на основе частичного скелета, который был обнаружен в плейстоценовых отложениях в Халиско, Мексика . Эти два вида различаются по нескольким аспектам, включая возраст. G. texanum относится к более древним слоям раннего плиоцена и раннего плейстоцена , тогда как G. cylindricum встречается исключительно в слоях позднего плейстоцена .
Глиптотерий представлял собой большого четвероногого (четвероногого) травоядного броненосца с бронированным панцирем (верхним панцирем), который состоял из сотен связанных между собой остеодерм (структур в дерме, состоящих из костей). Другие части доспехов покрывали хвост и крышу черепа , череп был высоким, с гипсодонтными (высококорончатыми) зубами. Что касается посткраниальной анатомии, характерны таз , сросшийся с панцирем, амальгаматный позвоночник, короткие конечности и маленькие пальцы для Glyptotherium и его родственников . Глиптотерий достигал 2 метров (6,56 футов) в длину и 400 килограммов (880 фунтов) веса, что делало его одним из крупнейших глиптодонтов, но не таким большим, как его близкий родственник Glyptodon или Doedicurus , самый крупный из известных глиптодонтов. Глиптотерий морфологически и филогенетически наиболее похож на глиптодон, однако они отличаются по ряду признаков. Глиптотерий в среднем меньше по размеру, у него более короткий панцирь, относительно длинный хвост и тонкая скуловая кость.
Глиптодонты впервые появились в эоцене , но значительно разнообразились в миоцене и плиоцене , в основном в сантакруцианских стоянках Аргентины . Однако в плейстоцене их разнообразие уменьшилось, хотя в этот период они достигли пика размеров. Глиптотерий считается примером североамериканской мегафауны , большая часть которой вымерла и, возможно, была уничтожена изменением климата или вмешательством человека. Глиптотерий в основном был травоядным , но также имел смешанную диету, состоящую из фруктов и других растений, обитавших на открытых лугах. Панцирь мог защитить животное от хищников, многие из которых сосуществовали с Glyptotherium за время его существования, в том числе «саблезубый кот» Smilodon , «костедробильная собака» Borophagus и «короткомордые медведи» ( Tremarctinae ).
История и филогения
[ редактировать ]Окаменелости, относящиеся к Glyptotherium, были обнаружены еще в 1870-х годах, когда инженеры-строители Дж. Н. Куатапаро и Сантьяго Рамирес собрали череп, почти полный панцирь и связанный с ним посткраниальный скелет глиптодонта из дренажного канала недалеко от Текишкиака , Мексика . Ранхолабрейский плейстоцена . возраст [1] [2] [3] Это была первая находка глиптодонта в Северной Америке. [2] [3] Куатапаро и Рамирес назвали окаменелости Glyptodon mexicanum в 1875 году, но окаменелости были утеряны . [1] [4] Другой вид Glyptodon был описан в 1889 году из Мексики, G. nathoristi, немецкими палеонтологами на основе останков панциря из плейстоценовых местонахождений в Эджутле , Оахака . [5] [6] Оба этих вида с тех пор стали синонимами G. cylindricum. [2] [3]
Первые окаменелости глиптотериев , описанные в Соединенных Штатах, были описаны в 1888 году палеонтологом Эдвардом Дринкером Коупом и состояли только из одной панцирной остеодермы, которая была собрана из нижнеплейстоценовых « слоев Equus » в округе Нуэсес , штат Техас . [7] [2] Коуп назвал свою остеодерму Glyptodon peltaliferus . [8] но Коуп не дал этому виду надлежащего описания, которое соответствовало бы правилам ICZN , что сделало его nomen nudum , и с тех пор он стал синонимом G. cylindricum . [2] [3] В следующем году Джозеф Лейди назвал Glyptodon floridanus на основе изолированных остеодерм панциря и частей хвостового панциря, хотя некоторые из них также были отнесены к G. peltaliferus . [9] которые были собраны из плейстоценовых отложений в округе ДеСото , Флорида . [10] [11] Этот вид теперь рассматривается как nomen vanum и считается младшим синонимом Glyptotherium cylindricum согласно обзору рода, проведенному американскими палеонтологами Дэвидом Джиллеттом и Клейтоном Рэем (1981). [12] [2]

Сам глиптотерий получил свое название в 1903 году, когда окаменелости, собранные экспедицией Американского музея естественной истории под руководством Дж. У. Гидли в пласты раннего плиоцена из формации Бланко в Ллано Эстакадо , штат Техас, были описаны Генри Фэйрфилдом Осборном как новый род и вид глиптодонтов. Глиптотерий Техасский . [7] Окаменелости были отложены в AMNH и состоят из панциря и связанных с ним посткраниальных элементов, одного из немногих G. texanum . известных скелетов [7] [2] Родовое название Glyptotherium происходит от греческих корней glyph, означающих «резной» или «рифленый», в честь его родственника Glyptodon , и therion , означающего «зверь», широко используемого суффикса для доисторических млекопитающих. Видовое название типового вида, G. texanum, произошло после открытия голотипа в Техасе. [7]
Еще одна находка была сделана в 1910 году, когда во время путешествия в Халиско , Мексика, охотник за окаменелостями Барнум Браун собрал полный спинной панцирь и несколько дополнительных окаменелостей, включая зубы, одной особи из плейстоценовых слоев этой области. [3] [13] Образец также был отправлен в Американский музей естественной истории, где Браун в 1912 году описал его как новый род и вид глиптодонта Brachyostracon cylindricum . [13] Видовое название cylindricum, означающее «цилиндрический», происходит от цилиндрической анатомии премоляров голотипа G. cylindricum. [13] Браун также рекомбинировал Glyptodon mexicanum в Brachyostracon mexicanum. [13] Brachyostracon теперь считается синонимом Glyptotherium на уровне рода, но G. cylindricum является действительным видом. [2] [14] В 1923 году Оливер Перри Хэй назвал новый вид глиптодонов , G. rivipacis , на основе окаменелостей, описанных Лейди из округа ДеСото , Флорида. [15] Этот вид теперь рассматривается как nomen nudum и синоним Glyptotherium cylindricum . [2] [16] Хэй также описал хорошо сохранившиеся окаменелости, в том числе элементы черепа и зубы, которые были собраны в слоях Ранчолабрея в Вулф-Сити и Синтоне, штат Техас, Коупа и которые были отнесены к Glyptodon peltaliferus . [14] Эти окаменелости с тех пор были отнесены к G. cylindricum. [2] [14]
Третий род североамериканских глиптодонтов, Boreostracon floridanum , был установлен Джорджем Гейлордом Симпсоном в 1929 году на основе нескольких изолированных экземпляров, обнаруженных AMNH в местонахождениях ранчолабрианской эпохи во Флориде, причем образец голотипа представляет собой заднюю часть панциря, обнаруженную в местности Семинол-Филд. в округе Пинеллас , штат Флорида. [12] [17] Симпсон отнес все окаменелости, ранее описанные из Флориды, к B. floridanum и считал, что все окаменелости глиптодонтов, обнаруженные в Северной Америке, не принадлежали глиптодонам . [17] Однако Симпсон решил не выделять новый род или вид для Glyptodon peltaliferus , но он все же считал, что они произошли от отдельной формы глиптодонта. [17] Boreostracon floridanum был синонимом Glyptotherium cylindricum. [2]
Исследования глиптодонтов Северной Америки уменьшились после исследований Гидли, Хэя, Симпсона и других, но некоторые палеонтологи по-прежнему ошибочно относили окаменелости с континента к глиптодонам . [14]
множество окаменелостей раннего плейстоцена собрал В 1927 году Университет Оклахомы в местности во Фредерике, штат Оклахома , включая несколько фрагментарных окаменелостей глиптодонтов, непарнокопытных , гомфотериев и верблюдовых . [18] Окаменелости глиптодонта первоначально были отнесены к глиптодонам в 1928 году. [19] [20] но не были должным образом описаны до 1953 года. [18] Они были описаны как новый род и вид Xenoglyptodon fredericensis в 1953 году на основании частичной нижней челюсти и нескольких зубов. [18] В настоящее время этот вид считается младшим синонимом Glyptotherium texanum . [2] [14] После того, как все эти роды были названы, серьезная переоценка не проводилась до 1981 года, когда Дэвид Джилетт и Клейтон Рэй опубликовали монографию о глиптодонтах Северной Америки. В своей монографии они объединили все ранее названные роды и виды в Glyptotherium, синонимизировали некоторые виды, а также исследовали экологию, анатомию и распространение рода. [14] Однако G. arizonae, G. floridanum и G. mexicanum были сохранены как действительные виды. [14] все они позже были синонимами G. texanum и G. cylindricum после открытия более полных скелетов, доказавших их синонимию. [2] [21] После более позднего рассмотрения первый вид был объявлен младшим синонимом G. texanum. [2] [21] и последние два синонима G. cylindricum . [2]
С 2000-х по 2020-е годы к этому роду были отнесены сотни дополнительных окаменелостей из Центральной Америки и Бразилии. [22] [23] К ним относятся окаменелости, ранее отнесенные к Glyptodon и Hoplophorus , поскольку многие окаменелости были поспешно отнесены палеонтологами 19-го века к обоим. [22] [24] Один из экземпляров, отнесенных к Glyptotherium , изолированной остеодерме спинного панциря, был собран Фридрихом фон Селлоу в карбонатных пещерах плейстоценового возраста в Лагоа-Санта , штат Минас-Жерайс , Бразилия, в начале 1800-х годов. [22] Позже он был описан как новый вид Hoplophorus , H. meyeri , в 1845 году датским палеонтологом Питером Вильгельмом Лундом . [25] [24] [22] Однако Лунд неправильно назвал таксон, сделав его nomen nudum . [22] Остеодерму отнесли к Glyptotherium в 2010 году, что сделало ее первым известным образцом Glyptotherium . [22] В 2022 году было описано множество окаменелостей Glyptotherium cylindricum , включая черепа, некоторые из сохранившихся патологий, вызванных человеком, которые были собраны в нескольких местах в Фальконе , северная Венесуэла, и относятся к позднему плейстоцену. [26]

Филогения
[ редактировать ]Glyptotherium — это род подсемейства Glyptodontinae, вымершей клады крупных, полностью бронированных броненосцев, которые впервые появились в позднем эоцене (около 33,5 млн лет назад) и вымерли в период вымирания в позднем плейстоцене — начале голоцена . [27] Glyptodontinae классифицировали как отдельное семейство или даже надсемейство до 2016 года, когда древняя ДНК была извлечена из панциря 12 000-летнего экземпляра Doedicurus (крупного глиптодонта с булавой) и почти полный митохондриальный геном был реконструирован (76-кратное покрытие) . ). Сравнение с ДНК современных броненосцев показало, что глиптодонты отделились от толипеутиновых и хламифориновых броненосцев примерно 33,5 миллиона лет назад, в позднем эоцене . [27] [28] Это побудило их переместить из собственного семейства Glyptodontidae в нынешнее Chlamyphoridae , переименованное в Glyptodontinae. подсемейство [28] Следуя этим генетическим данным и летописи окаменелостей, глиптодонты быстро развили свою характерную анатомию и огромные размеры (также известные как гигантизм), возможно, из-за похолодания, более сухого климата, расширения саванн и размеров хищников, таких как Арктотерий и Смилодон . [27] Chylamyphoridae — группа отряда Cingulata, в которую входят все современные броненосцы в дополнение к другим ископаемым группам, таким как Pachyarmatheriidae и Pampatheridae . Cingulata сама входит в базальную группу млекопитающих Xenarthra, которая включает в себя ряд американских групп млекопитающих, таких как Vermilingua (муравьеды) и Folivora (ленивцы и наземные ленивцы ) в отряде Pilosa . Следующий филогенетический анализ был проведен Фредериком Дельсуком и его коллегами в 2016 году и представляет филогению Cingulata с использованием древней ДНК Doedicurus для определения положения ее и других глиптодонтов: [27] [29]

Внутренняя филогения Glyptodontinae запутана и постоянно меняется: многие виды и семейства созданы на основе фрагментарного или недиагностического материала, который не имеет всестороннего анализа. [30] [24] [2] Обычно его считают отдельным семейством, но анализ ДНК свел его к подсемейству с племенами, а не подсемействами. Одно племя, Glyptodontini (обычно обозначаемое как Glyptodontinae), представляет собой группу более молодых и крупных глиптодонтов, которые эволюционировали в среднем миоцене (около 13 млн лет назад) с Boreostemma , [31] но разделен на 2 рода: Glyptodon на юге и Glyptotherium на севере, [2] хотя глиптодон также жил на севере Южной Америки, в Колумбии . [2] [32] Glyptodontini часто считают более базальным по сравнению с большинством других глиптодонтов, таких Doedicurus , Hoplophorus и как Panochthus . Glyptodontini отличается от других групп, например, тем, что у него большие конические бугорчатые остеодермы отсутствуют или присутствуют только на каудальной (к хвостовой) выемке на заднем конце панциря, а также орнамент панциря на панцире, отличный от хвоста. [33] Родственным таксоном или ближайшим родственником другого таксона Glyptotherium является род Glyptodon , который появился в среднем плейстоцене в Аргентине. [33] [30] Glyptotherium почти идентичен Glyptodon во многих аспектах, настолько, что первые окаменелости Glyptotherium описанные были ошибочно идентифицированы как окаменелости Glyptodon . [14] [1]
Когда Glyptotherium был впервые описан Генри Фэйрфилдом Осборном в 1903 году, он поместил его на уровень семейства Glyptodontidae и, хотя он был похож на Glyptodon , Осборн заявил, что он был ближе по внешнему виду и классификации к Panochthus и Neosclerocalyptus (тогда Sclerocalyptus ). [7] Барнум Браун считал, что его род Brachyostracon и два его вида, B. cylindricum и B. mexicanum , принадлежали к отдельному семейству глиптодонтов, основываясь на удлинении черепа последнего вида и ширине панциря по сравнению с его длиной. [13] Однако Браун не придумал никакого нового названия для этого семейства, а вместо этого отнес этот род к Sclerocalyptidae вместе с южноамериканскими глиптодонтами, такими как Panochthus , Neosclerocalyptus и Plohophorus . [13] Джордж Гейлорд Симпсон классифицировал свой род Boreostracon как близкого родственника Glyptodon , но все же считал, что существует несколько североамериканских родов глиптодонтов. [17] Мид (1953) отнес ксеноглиптодон к глиптодонтам, близким к другим североамериканским родам, но не указал на его связь с южноамериканскими родами. [18] Ниже приведен филогенетический анализ Glyptodontinae, проведенный Cuadrelli et al ., 2020, с Glyptodontidae как семейством, а не подсемейством, который фокусируется на продвинутых глиптодонтах: [33]
| Хламифориды |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Описание
[ редактировать ]
Как и у его живого родственника, броненосца, у Glyptotherium был панцирь, покрывавший все его туловище, а меньший панцирь также покрывал крышу черепа, как у черепахи . Однако, в отличие от панциря (верхнего панциря) черепахи, панцирь Glyptotherium состоял из сотен небольших шестиугольных чешуек, при этом у Glyptotherium сохранилось до 1800 остеодерм (костных структур в дерме) или более у каждой особи. [34] Осевой скелет глиптодонтов демонстрирует обширное сращение позвоночного столба, а таз (тазовая кость) сращен с панцирем, что делает таз полностью неподвижным. [14] Глиптотерий был очень гравипортальным и имел короткие конечности, очень похожие на таковые у других глиптодонтов. Большие хвосты глиптодонтов, вероятно, служили противовесом остальной части тела, а хвостовой панцирь Glyptotherium заканчивался тупой трубкой, состоящей из 2-3 сросшихся трубок. [2] в отличие от южноамериканских глиптодонтов с булавчатым хвостом. [35] Пальцы Glyptotherium очень толстые и приспособлены к нагрузке, хотя у некоторых сохранились большие когтевые ножны, которые имели промежуточную морфологию между когтями и копытами. [14] В плейстоцене филогенетическое разнообразие глиптодонтов уменьшилось, но увеличилось в размерах. [26] [2] Вес и размеры глиптотериев различаются, но G. texanum был меньше и легче, чем более поздний вид, G. cylindricum. Один экземпляр G. cylindricum оценивался в 350-380 килограммов по сравнению с 457 кг его родственника Glyptodon reticulatus. [26] Самые большие оценки массы Glyptotherium основаны на взрослых экземплярах из Бразилии и Аризоны: по одной оценке экземпляр G. cylindricum составляет 710 кг (1570 фунтов), а особь G. texanum - 1165 кг (2568 фунтов). [36] Один экземпляр G. cylindricum , AMNH 15548, сохранил длину панциря 1690 сантиметров по сравнению с всего 1400 см у MSM P4464, экземпляра G. texanum . [2]

Glyptotherium texanum отличается от G. cylindricum конфигурацией внешней скульптуры симметричных шестиугольных остеодерм дорсального панциря. В остеодермах взрослого G. texanum центральные фигуры остеодерм плоские или слабовыпуклые и очень крупные, а у G. cylindricum — плоские или вогнутые и значительно мельче. [2] [37] У обоих видов центральные фигуры остеодерм увеличиваются по направлению к краям панциря. [2] [21] Хотя считалось, что G. texanum крупнее G. cylindricum , считается, что этот род был одинаково крупным на протяжении всего плиоцена и плейстоцена. [2]
Череп и нижняя челюсть
[ редактировать ]Зубной ряд глиптодонтов имеет все гипсодонтные (зубы с высокой коронкой, приспособленные для выпаса скота), коренные, щечные зубы, которые являются одними из наиболее известных гипсодонтов у наземных млекопитающих. [38] Черепа глиптодонтов имеют несколько уникальных особенностей; верхняя увеличены и небная кости по вертикали, чтобы освободить место для моляров, а черепная коробка брахицефальная , короткая и плоская. [39] Кожная броня не ограничивалась панцирем и хвостом, поскольку крыша черепа была защищена «головным щитом», сделанным из остеодерм. [26] [40] Некоторые палеонтологи предположили, что у Glyptotherium и некоторых глиптодонтов также был хоботок или большая морда, похожая на таковую у слонов и тапиров. [41] но немногие приняли эту гипотезу. [2] [26] известен только один полный череп Из Glyptotherium texanum , в то время как его родственники, такие как Glyptodon и Neosclerocalyptus , известны из многих черепов, что дает ограниченное представление о его анатомии. [42] [43] узкие Скулы Glyptotherium ; , тонкие, почти параллельные, близки к сагиттальной плоскости при взгляде спереди у глиптодона эта структура более широкая, прочная, расходящаяся, а не параллельная и более латерально расположенная. Glyptotherium и другие глиптодонты сохраняют большие носовые ходы и пазухи, которые, возможно, имели ноздри, приспособленные для дыхания в холодном засушливом климате Америки во время плейстоцена . [44] В свою очередь, подглазничные отверстия узкие и не видны при виде спереди у Glyptotherium , а у Glyptodon они широкие и хорошо заметны при взгляде спереди. При взгляде сбоку дорсо-вентральная высота между крышей черепа и небной плоскостью у Glyptodon уменьшается спереди, в отличие от Glyptotherium ; кончик носа находится в более низкой плоскости по отношению к скуловой дуге у Glyptodon , но у Glyptotherium находится выше плоскости скуловой дуги. У Glyptotherium окклюзионный латеральный профиль слегка изогнут, тогда как у Glyptodon он сильно изогнут . У Glyptodon 1-й коренной зуб (сокращенно mf1) явно трехлопастный (трехлопастный) как с лингвальной, так и с лабиальной стороны, почти такой же трехдольный, как mf2; напротив, у Glyptotherium наблюдается очень низкая трехдольчатость mf1, эллиптическая в поперечном сечении, mf2 слабо трехлопастная, а mf3 трехлопастная. У обоих родов от mf4 до mf8 полностью трехлопастные и серийно идентичные. [2]
Мандибулы Glyptotherium и Glyptodon очень похожи, но у Glyptotherium нижняя челюсть меньше примерно на 10%. Угол между окклюзионной плоскостью и передним краем восходящей ветви у Glyptotherium составляет около 60°, а у Glyptodon — 65° . Вентральный край горизонтальной ветви у Glyptodon более вогнутый , чем у Glyptotherium . значительно расширена в Область симфиза у Glyptotherium передне-заднем направлении по сравнению с Glyptodon . эллипсоидные Зубы mf1 у Glyptotherium , а mf2 субмолярные , тогда как у Glyptodon оба зуба трехлопастные. [2]
Панцирь и остеодермы
[ редактировать ]
Глиптотерий и все другие глиптодонты имели большой дорсальный панцирь, покрывающий большую часть спины и состоящий из соединенных между собой остеодерм. Панцирь Glyptotherium был короче, чем у его родственника Glyptodon , но гораздо более удлинен, чем у Boreostemma . Верхняя точка панциря находилась в центре средней линии, а у глиптодона она была слегка смещена. Панцирь глиптотерия был сильно изогнутым, с выпуклой предподвздошной и вогнутой после подвздошной, что придавало ему седловидный нависание над хвостом. [2] [31] У Glyptodon высота верха и низа панциря составляет 60% его общей длины, тогда как у Glyptotherium он выше и составляет ок. 70%. Вентральные края панциря у Glyptotherium гораздо более прямоугольные и менее выпуклые, чем у Glyptodon. У Glyptotherium остеодермы в передне-латеральных областях панциря менее анкилозированы, чем у Glyptodon , что позволяет предположить, что передне-латеральные области панциря у первых имели большую гибкость. Остеодермы каудального отверстия более конические у Glyptodon и более округлые у Glyptotherium , хотя у последних анатомия остеодерм каудального отверстия варьируется в зависимости от пола. [21] а у глиптодона это зависит от возраста. [45] часто используются для дифференциации этих двух таксонов, Хотя Glyptotherium и Glyptodon они имеют очень схожую морфологию остеодермы, которая различается только в нескольких областях. Оба рода имеют очень толстые остеодермы по сравнению с таковыми у многих южноамериканских глиптодонтов, таких как Hoplophorus и Neosclerocalyptus , но Glyptotherium всегда сохраняет «розеточный» рисунок, где центральная фигура остеодермы окружена рядом периферийных фигур. Некоторые экземпляры глиптодонов сохраняют эти «розетки», но у других они отсутствуют. Центральные и радиальные борозды глубже и шире у Glyptodon (около 4–6 мм), чем у Glyptotherium (около 1–2,4 мм). [2] [21] Примечательно, что у остеодермов Glyptotherium сохраняются небольшие промежутки для волосяных фолликулов в бороздах, что указывает на то, что у Glyptotherium был «нечеткий» панцирь с выступающим мехом. Количество фолликулов варьируется в зависимости от возраста и площади панциря: у молодых особей фолликулов больше, чем у взрослых, а в боковой, каудальной и задней частях панциря известно меньше фолликулов. [46] [21]
Каудальные кольца
[ редактировать ]
Глиптотерий является глиптодонтом, то есть его хвостовой панцирь состоит из ряда хвостовых колец, заканчивающихся короткой хвостовой трубкой, в отличие от булавовидных концов у других глиптодонтов, но морфология у Glyptotherium, Glyptodon и Boreostemma различается . В целом, Boreostemma сохраняет хвостовой панцирь, более похожий на Glyptotherium, чем на Glyptodon . [31] длиннее, Хвостовой панцирь у Glyptotherium чем у Glyptodon , при этом у одного экземпляра G. texanum (UMMP 34 826) сохранился комплект хвостового панциря метровой длины. У Glyptotherium длина хвостового панциря составляет около 50% от общей длины спинного панциря, тогда как у Glyptodon это значение ниже и составляет около 30-40%. У глиптодона 8–9 полных хвостовых колец плюс одна хвостовая трубка, но у Glyptotherium сохраняется 1 неполное хвостовое кольцо в дополнение к 8–9 полным хвостовым кольцам и хвостовой трубке. У обоих родов каждое хвостовое кольцо состоит из двух-трех поперечных рядов анкилозированных остеодерм, причем самый дистальный ряд остеодерм имеет более или менее развитую коническую морфологию. У Glyptotherium у некоторых экземпляров (например, AMNH 95,737) небольшое количество конических остеодерм (обычно две). В этом отличие от Glyptodon , у которого большинство остеодерм дистального ряда (до 12) имеют четкую коническую морфологию. короче Конечная хвостовая трубка у глиптодона . У Glyptotherium конечная трубка состоит из 2–3 анкилозированных колец, тогда как у Глиптодон имеет всего два анкилозированных кольца. У Glyptotherium эта хвостовая трубка представляет собой ок. 20% общей длины хвостового панциря, тогда как у глиптодона эта структура составляет 13% общей длины. [2]
Палеобиология
[ редактировать ]Поза
[ редактировать ]Было сделано несколько интерпретаций позы глиптодонта. [47] первоначально британским палеонтологом Ричардом Оуэном в 1841 году с использованием сравнительной анатомии. [48] Оуэн предположил, что фаланги несут вес из-за своей короткой и широкой физиологии, в дополнение к данным посткраниального скелета. [48] Было также высказано предположение, что вертикальное положение глиптодонтов возможно , сначала Сенешалем (1865), который заявил, что хвост может быть равновесием для передней половины тела, а также методом поддержки ног. [49] [47] Позже были проведены линейные измерения, которые пролили свет на эту гипотезу и показали, что прямохождение возможно. [50] [51] Сочленение надколенника с бедренной костью позволяет предположить возможность вращения голени во время разгибания колена и, возможно, даже фиксации колена. [52]
Кормление и диета
[ редактировать ]
глиптотерий Традиционно считается, что питается влажными прибрежными травами . [53] Этот род в основном был травоядным, но также имел смешанную диету, состоящую из растений C3 и C4, основываясь на изотопном анализе образцов зубов, извлеченных из позднеплейстоценового местонахождения Cedral в Сан-Луис-Потоси , Мексика. [54] В данной местности сохранились растения С4 из семейств Poacea , Amaranthacea и Quenopodiacea . [55] это означает, что они были возможными источниками пищи для глиптотериев . [54] В частности, Седрал представлял собой территорию с горячими источниками и открытыми лугами рядом с ними, что позволяет предположить, что Glyptotherium питался на лугах возле источников воды, как и привычки питания современных капибар . [41] [54] Дополнительный изотопный анализ Glyptotherium и гигантского наземного ленивца Eremotherium показал, что они имеют одинаковые уровни изотопов с современными земноводными бегемотами , что указывает на то, что они были полуводными травоядными животными, питавшимися водными растениями. Дополнительные исследования образцов зубов из восточной Бразилии показывают, что Glyptotherium паслись во влажных, равнинных тропических и субтропических средах обитания вдоль рек или источников воды, что подтверждает гипотезу полуводных Glyptotherium . [36] [23] [56] Дополнительные изотопные данные из Бразилии позволяют предположить, что фрукты также были частью рациона Glyptotherium , хотя в целом их доля составляла лишь около 20%. [57] У глиптотериев и всех других глиптодонтов были гипсодонтные зубы, зубы с высокой коронкой и шероховатыми плоскими поверхностями, приспособленными для измельчения и измельчения, которые были приспособлены для разрушения песчанистого волокнистого материала, такого как трава. [58] [41] Эта диета Glyptotherium контрастирует с диетой их родственников Pampatheres , которые считались насекомоядными или травоядными. [54] [59] Как и большинство других ксенартранов, глиптодонты имели более низкие потребности в энергии, чем большинство других млекопитающих. Они могли выжить при более низких нормах потребления, чем другие травоядные животные с аналогичной массой. [60]
Способности копать
[ редактировать ]Многие современные виды броненосцев обладают способностями к рытью и имеют большие когти, приспособленные для соскребания земли, чтобы прокладывать норы или добывать пищу под землей. [61] [62] Кроме того, большую часть их рациона составляют насекомые и другие беспозвоночные, живущие под землей. [63] в отличие от травоядных Glyptotherium . [64] Поскольку глиптодонты принадлежат к семейству броненосцев, их ископаемые способности исследовались несколько раз. [47] Оуэн (1841 г.) выступал против этой идеи, хотя сопротивление исходили от Нодо (1856 г.) и Сенешаля (1865 г.), которые считали, что для этого рода земледелие возможно. [47] [48] Однако эволюция жесткого панциря в отличие от гибкого у современных броненосцев, а также слабо развитый дельтовидный гребень на плечевой кости (плечевая кость) предоставили доказательства против ископаемых гипотез. У локтя был большой диапазон движений, как и при копании поясной извилины, но это, скорее всего, связано с адаптацией к размеру. [65] [47]
Онтогенез
[ редактировать ]Молодые и взрослые особи Glyptotherium texanum, обнаруженные в местонахождениях Бланкан в Аризоне, сохраняют почти полную серию роста, одну из немногих, известных у глиптодонтов. [21] Зубы Glyptotherium сохраняют гипсодонтию, при которой зубы не перестают расти. [21] У молодых людей уже известен полностью артикулированный контакт остеодермы с остеодермой, даже в их возрасте. [21] Рост остеодерм продолжался от молодых особей до взрослых и прекращался, когда остеодермы становились анкилозированными (сросшимися). [17] [21] Боковой профиль панцирей молоди плавно выпуклый, переходящий в высокоарочный панцирь взрослых особей. Другое онтогенетическое изменение касается борозд и периферических фигур остеодермы; центральные фигуры являются самыми крупными относительно борозд у новорожденных и подростков, но у взрослых это соотношение становится ниже. Однако у G. cylindricum остеодермы растут намного быстрее, а борозды намного меньше. Остеодермы также относительно толще у молодых особей Glyptotherium по сравнению со взрослыми. [21]
Половой диморфизм
[ редактировать ]Особи Glyptotherium texanum, обнаруженные в местонахождениях Бланкан в Аризоне, сохраняют половой диморфизм между особями мужского и женского пола. Хвостовое отверстие, крупные конические остеодермы, защищающие основание хвоста, самцов и самок отличаются тем, что краевые остеодермы самцов гораздо более конические и выпуклые, чем у самок. Даже в панцирях новорожденных Glyptotherium краевые остеодермы имеют либо коническую, либо плоскую форму, что позволяет определить их пол. [21]
Остеодермы и защита
[ редактировать ]
Остеодермы Glyptotherium трабекулярного состоят из губчатого ядра между двумя компактными слоями. Каждая шестиугольная остеодерма соединяется с соседними остеодермами швами, образуя большой прочный панцирь, подобный панцирю современных животных. Однако панцири глиптодонтов, таких как Glyptotherium, были гораздо менее гибкими, чем у современных броненосцев. [66] Трабекулярное ядро состояло из опорных стоек средней толщины 0,25 мм, которые составляли центральную опору остеодермы. Механический анализ показал, что меньшие области нагрузки, представляющие собой более острые предметы, вызывают более высокие напряжения, чем те, которые вызываются большими тупыми предметами. Это можно понимать как то, что естественная структура эволюционировала, чтобы выдерживать тупые удары крупных предметов, таких как хвостовые булавы, а не как защита от острых предметов, таких как зубы. [37] Это является дополнительным доказательством теории о том, что глиптодонты использовали свои панцири для внутривидовых боев, используя хвостовые булавы, а также для защиты. [37]
Болезни и патологии
[ редактировать ]Частичный посткраниальный скелет Glyptotherium cylindricum , найденный на северо-востоке Бразилии и включающий частичные конечности, сохранил 3 различных типа артрита: один тип воспалительного артрита ( спондилоартропатия ), один тип кристаллического артрита ( болезнь отложения пирофосфата кальция ) и один пролиферативный артрит ( остеоартрит ). Спондилоартропатия была наиболее распространенной, с эрозией костей правой локтевой кости, правой и левой лучевых костей, левой бедренной кости, а также правой и левой большеберцово-малоберцовой кости и ассоциировалась со склерозом костей правой локтевой кости и лучевой кости. Болезнь отложения пирофосфата кальция присутствует на суставной поверхности левой надколенника, а остеоартрит представлен остеофитами на некоторых участках левой задней конечности. [67] На нескольких остеодермах, также обнаруженных в Бразилии, обнаружен эктопаразитизм, поражения и наросты, некоторые из этих инфекций, вероятно, были вызваны блохами. [68]
Палеоэкология
[ редактировать ]Глиптотерии в основном паслись на лесных лугах и древесных саваннах, хотя, судя по местонахождениям окаменелостей из Мексики, они, возможно, предпочитали луга рядом с источниками воды. [54] Из-за их широкого распространения Glyptotherium палеоэкология могла различаться в зависимости от региона и двух его видов. [23] [54]

В Северной Америке Glyptotherium известен от двух разных видов, обитавших в трех разных интервалах: Бланканском , Ирвингтонском и Ранчолабрейском , на территории нескольких штатов США и Мексики. [2] Однако Glyptotherium не был обнаружен в водоемах Тихого океана в Соединенных Штатах или к западу от реки Колорадо . [2] вместо этого они присутствуют в основном на Атлантической равнине и Внутренних равнинах . [69] Во времена Бланкана Glyptotherium texanum сосуществовал со многими эндемичными родами Северной Америки, поскольку Берингия еще не сформировалась. Из-за этого фауна Бланкана резко контрастировала с фауной последующего плейстоцена . В слоях бланканского возраста западного Техаса, Нью-Мексико и Аризоны сохранились хоботные гомфотеры Stegomastodon и Cuvieronius , непарнокопытные, представленные такими травоядными, как Nannipus и Equus . Известны и другие ксенартраны, такие как мегалонихидный наземный ленивец Megalonyx и милодонтид Paramylodon . Также были обнаружены окаменелости мелких млекопитающих, таких как насекомоядные Hesperoscalops и Sorex . Гигантский ископаемый суслик Paenemarmota также известен из Бланкана. [70] К хищникам относятся необычный родственник собак, сокрушающий кости, Борофаг и «бегущая гиена» Часмапортет . [71] кроме «саблезубой» кошки Smilodon gracilis . [72] Также были обнаружены некоторые изолированные окаменелости птиц, состоящие из стервятников, соколов и, возможно, врановых. [71] [73] [74] Окаменелости из Гватемалы были найдены на больших высотах, что показывает, что Glyptotherium легко адаптируется и может жить в различных средах. [75] Это подтверждается общей диетой Glyptotherium и открытиями ископаемых в тропических, субтропических, лесных и даже полуводных средах. В мексиканском штате Юкатан окаменелости Glyptotherium и наземного ленивца Paramylodon известны из богатых водой районов, таких как прибрежные леса и болота, а не из открытых лугов. [76]
В Бразильском межтропическом регионе (BIR) на востоке Бразилии Glyptotherium пасся в древесных саваннах, тропических лугах и других травянистых районах вблизи источников воды. Крупные мезотравоядные млекопитающие в БИР были широко распространены и разнообразны, включая коровоподобных токсодонтидов Toxodon Platensis и Piauhytherium , Xenorhinotherium литоптерна макраухениид и непарнокопытных, таких как Hippidion Principale и Equus neogaeus . Токсодонтиды также были крупными смешанными питателями и жили в лесных районах, в то время как непарнокопытные почти полностью были травоядными. В этом районе также присутствуют другие окаменелости ксенартранов из нескольких различных семейств, таких как гигантский наземный ленивец Eremotherium , сцелидотерииды Catonyx и и Valgipes , милодонтиды Glossotherium , Ocnotherium мегатериидный Mylodonopsis . более мелкие наземные ленивцы, такие как мегалонихиды Ahytherium и Australonyx и нотротериид Nothrotherium В этом районе также были обнаружены . Эремотерий был универсалом, а Нотротерий - специалистом по деревьям в лесах с низкой плотностью, и Вальгипес был промежуточным из двух видов, обитавших в древесных саваннах. другие глиптодонты и поясные животные, такие как пасущийся глиптодонт Panochthus и всеядные пампатеры Pampatherium и Holmesina На открытых лугах присутствовали и Notiomastodon Platensis . В БИР также был обнаружен вид хоботных, , который также присутствовал и пасся на открытых лугах смешанного типа. Среди хищников были некоторые из крупнейших известных хищников-млекопитающих, такие как гигантский кошачий популятор Smilodon и медведь Arctotherium wingei . [55] [56] Из BIR также известно несколько существующих таксонов, таких как гуанако , гигантские муравьеды , ошейниковые пекари и полосатые свиноносые скунсы . [77] В БИР также известны два типа современных млекопитающих, питающихся крабоедами: енот -крабоед и лисица-крабоед , что указывает на то, что крабы также присутствовали в этом регионе. [77] Окружающая среда БИР неясна, поскольку оба вида были травоядными, но появление в этом районе древесных ископаемых обезьян Protopithecus и Caipora вызывает путаницу в отношении палеосреды этого района. Считалось, что большая часть Бразилии была покрыта открытой тропической растительностью серрадо в позднем плейстоцене , но если Protopithecus и Caipora были древесными, их присутствие предполагает, что в позднем плейстоцене в этом регионе мог быть густой закрытый лес. [77] [78] Вполне возможно, что на протяжении всего изменения климата в позднем плейстоцене в этом регионе чередовались сухие открытые саванны и закрытые влажные леса. [79]
Великая американская развязка
[ редактировать ]
Южная Америка, континент, на котором произошли глиптодонты, была изолирована после распада суши Гондваны в конце мезозойской эры . [80] Этот период отделения от остальных континентов Земли привел к эпохе уникальной эволюции млекопитающих, с доминированием таких групп, как сумчатые , ксенартраны и некопытные, в отличие от фауны млекопитающих Северной Америки. Сумчатые, вероятно, попали в Южную Америку до ее отделения от остальной части Гондваны в позднем меловом или палеогеновом периоде , хотя происхождение отрядов млекопитающих, таких как Xenarthra и Notoungulata, оказавшихся на континенте, остается загадкой. [81] До образования Панамского перешейка было несколько перемещений внешних млекопитающих в Южную Америку , например, приматов и грызунов , которые, возможно, сплавились на континент из Африки, а также перемещение летучих мышей посредством полета. [82] [83] Что касается фауны Северной Америки, то современные группы, такие как псовые, кошачьи, урсиды , тапириды , антилокаприды и непарнокопытные, населяли этот регион в дополнение к вымершим семействам, таким как гомфотерии, амфициониды и маммутиды . [84] [80] Великий американский обмен не вступил в свою самую большую стадию до тех пор, пока Панамский перешеек не образовался 2,7 миллиона лет назад во время бланканского этапа плиоцена. [85] [86] [87] Это усилило перемещение глиптодонтов, наземных ленивцев, капибар, пампатеров, птиц-террористов и сумчатых в Северную Америку по центральноамериканскому маршруту, а также обратную миграцию копытных, хоботных, кошачьих, псовых и многих других групп мегафауны в Южную Америку. [88] Период после основания перешейка стал свидетелем исчезновения или истребления многих групп, в том числе южноамериканских птиц-террористов, токсодонтов, макраухениид , пампатеров, наземных ленивцев и глиптодонтов. [89] [90]
Сам Glyptotherium был частью этого обмена, развиваясь в Бланкане Мексики и США после образования перешейка и его иммиграции. [2] [88] Географическая изоляция Glyptotherium в сочетании с адаптацией к различным средам Северной Америки привела к тому, что Glyptotherium отделился от других глиптодонтин. [2]
Хотя G. texanum эволюционировал в Северной Америке в G. cylindricum в Ранчолабрии, он эмигрировал на юг, в Центральную и некоторые части северной части Южной Америки, примерно 15 000–20 000 лет назад. [26] [22] [91] Это также связано с экологической сегрегацией: горные барьеры в Колумбии, возможно, разделяют Glyptodon и Glyptotherium . [32] Впоследствии Glyptodon обитал в основном в Андах и прибрежных районах, при этом Glyptotherium был известен из лугов и слабо засаженных лесом низинных отложений вблизи водных акваторий, что послужило причиной его распространения в тропических регионах Венесуэлы и восточной Бразилии. [87] [23] [92] Возвращение группы в Южную Америку из Северной Америки также наблюдалось у родственного поясного семейства Pampatheriidae, [86] возможно, этому способствуют широкие прибрежные низменности, открытые во время ледниковых периодов на побережьях Карибского бассейна и Атлантического океана, что позволяет мигрировать между Флоридой, Мексикой, Центральной Америкой и/или северной частью Южной Америки. [87] [93]


Хищничество
[ редактировать ]Смилодон, возможно, время от времени охотился на Glyptotherium , судя по черепу одной особи Glyptotherium texanum, извлеченной из плейстоценовых отложений в Аризоне, с характерными эллиптическими следами проколов, которые лучше всего соответствуют таковым у кошки -махайродонта , что указывает на то, что хищник успешно прокусил череп через панцирь. головной щит. [94] Рассматриваемый глиптотерий был молодым человеком с еще развивающимся головным щитом, что делало его гораздо более уязвимым для нападения кошки. [95] Хотя первоначально Джордж Брандес предположил, что это возможно в 1900 году, [96] Клыки смилодона не могли пробить толстые остеодермы панциря глиптодонтов. [97]
Брандес выдвинул гипотезу, что эволюция толстого панциря глиптодонтов и длинных клыков махайродонтов была примером коэволюции. [96] но Биргер Болин в 1940 году утверждал, что клыки слишком хрупкие, чтобы нанести ущерб панцирю глиптодонтов. [97] [45] Однако эволюция дополнительных защитных структур могла произойти в ответ на прибытие смилодонов и арктотерий в Энсенадан . [45]
В 2017 году была описана правая локтевая нога взрослого Glyptotherium , собранная в слоях позднего плейстоцена в штате Риу-Гранди-ду-Норти, Бразилия, которая имела несколько следов грызения от нового ихновида Machichnus , M. fatimae , которые, возможно, были вызваны молодь вида псовых Protocyon troglodytes или взрослая особь Cerdocyon тыс . [98]
Отношения с людьми
[ редактировать ]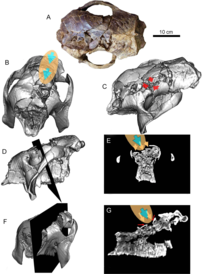
Первое сообщение о возможном потреблении человеком или взаимодействии с Glyptotherium , штат Техас, было описано несколько остеодерм, которые, возможно, потреблялись людьми или его окаменелостями появилось в 1958 году, когда на участке Кловис в Льюисвилле . [99] [100] Однако эта идея человеческого потребления имеет мало доказательств, подтверждающих ее. [26] В 2022 году было описано множество окаменелостей Glyptotherium cylindricum, включая черепа, которые были собраны в нескольких местах в Фальконе , северная Венесуэла, и относятся к позднему плейстоцену . [26] Эти открытия были примечательны не только тем, что в них сохранились черепа, но и в четырех из них были обнаружены переломы в лобно-теменной области, что характерно для всех черепов. Визуальные и компьютерные данные указывают на то, что они, вероятно, были вызваны механическим воздействием при прямой перкуссии, скорее всего, ударом каменным рубилом или дубинкой, в результате чего кости в этой области раскололись на мягкие внутренние ткани черепа. Несмотря на то, что черепа были целыми и не имели признаков тафономического искажения или перемещения, челюсти у них часто отсутствовали. Челюсти могли быть удалены, чтобы «охотники» могли получить доступ к жевательным мышцам и языку и съесть их. Гипотеза о сосуществовании первых людей-охотников-собирателей и глиптодонтов в Южной Америке была впервые выдвинута в 1881 году на основе находок окаменелостей в аргентинских пампасах . [101] и с тех пор было обнаружено множество находок окаменелостей позднего плейстоцена, которые свидетельствуют о хищничестве человека на глиптодонтах. В этот период люди охотились на большое количество ксенартранов, населявших пампасы, и имеются данные, показывающие, что небольшой (300–450 кг) глиптодонт Neosclerocalyptus , [102] на броненосца Евтата и гигантского (2 тонны) глиптодонта Дедикура , крупнейшего из известных глиптодонтов, охотились. [103] Единственным другим свидетельством нападения человека за пределами Пампаса был частичный панцирь, обнаруженный также в Венесуэле, который был выпотрошен людьми. Находки в местонахождениях в Фальконе показали первые признаки человеческой охоты на черепах глиптодонтов, но Glyptotherium также оказался более беззащитным, чем глиптодонты типа Doedicurus . [26] [104]
Распределение
[ редактировать ]Глиптотерий — единственный известный глиптодонт Северной Америки, известный из нескольких регионов континента в разные периоды. На бланканском этапе раннего плиоцена населял только центральную Мексику , Glyptotherium texanum основываясь на открытии единственной остеодермы Glyptotherium texanum из раннего плиоцена слоев Гуанахуато , центральная Мексика, датируемой ок. 3,6 миллиона лет назад. [2] [105] На бланкан -ирвингтонском этапе раннего плейстоцена Аризона окаменелости G. texanum известны на большей части территории Мексики, а также в штатах США: , Техас , Оклахома , Флорида и , возможно, Южная Каролина . [2] [105] В ранчолабрии позднего плейстоцена , G. cylindricum произошел от G. texanum и его окаменелости были обнаружены в северной Венесуэле , восточной Бразилии , Центральной Америке , Мексике и штатах США Техас, Луизиана , Флорида и Южная Каролина. [2] [105] [106] [53] Окаменелости Glyptotherium из раннего плиоцена не были обнаружены в Центральной Америке, но вполне вероятно, что G. texanum населял этот район во время Великого американского биотического обмена . Окаменелости глиптодонтов среднего-позднего ирвингтонского периода не известны в США, что создает «пробел глиптодонтов» в летописи окаменелостей США. [2] Однако Glyptotherium во время этой «паузы» в Центральной Америке регистрируется , что позволяет предположить возможное перемещение Glyptotherium в южные районы во время ледникового периода. [91]

Окаменелости глиптотериев были собраны в Центральной Америке, в Гватемале . [75] Коста-Рика , [107] Гондурас , [108] Сальвадор , [109] и Панама . [110] Окаменелости из Центральной Америки обычно изолированы и фрагментарны, большинство из них представляют собой остеодермы или отдельные молариформы. [2] [111] В 2023 году был описан связанный скелет G. cylindricum , включая элементы черепа и конечностей, из Гватемалы, наиболее полный экземпляр из этого региона. [112] Судя по их палеобиогеографии, наиболее вероятно, что первые популяции Glyptotherium возникли в Центральной Америке во время Великого американского биотического обмена в позднем плиоцене. [2] [21] Окаменелости глиптотериев из Центральной Америки иногда относят к неопределенному виду. [23] [22] но большинство из них относится к Glyptotherium cylindricum или его синонимам. [75] [107] Это определение также основано на возрасте окаменелостей, поскольку возраст окаменелостей G. texanum оценивается в диапазоне от плиоцена до раннего плейстоцена, тогда как возраст окаменелостей G. cylindricum ограничивается поздним плейстоценом. [2] [113]
Хотя обычно его считают исключительно североамериканским родом, [113] [22] [2] окаменелости Glyptotherium из северной части Южной Америки в таких регионах, как Бразилия и Венесуэла . были обнаружены [22] [26] Окаменелости из Южной Америки обычно представляют собой только остеодермы или хвостовые кольца и иногда неопределенны на видовом уровне, но, скорее всего, они принадлежат Glyptotherium cylindricum . [2] [22] [26]
Вымирание
[ редактировать ]Хронология вымирания мегафауны (например, Glyptotherium ) в позднем плейстоцене оспаривается. [69] В Соединенных Штатах последняя надежная прямая радиоуглеродная дата для глиптотерия - 23 230 ± 490 лет назад, из пещеры Лаубах № 3 , штат Техас. [114] [69] Glyptotherium группируется вместе с Eremotherium , Holmesina и Paramylodon как имеющие надежные окончательные даты до окончания последнего ледникового максимума Северной Америки. [115] Однако статистический анализ предполагает, что более позднее выживание до конечного плейстоцена Соединенных Штатов возможно, что основано на ошибках выборки, связанных с необычной фауной, и отсутствии надежных датировок по влажной атлантической равнине из-за плохой сохранности. [69]
В Южной Америке сгоревшие останки Glyptotherium были неточно датированы периодом от 16 375 ± 400 до 14 300 ± 500 лет назад по радиоуглероду в Муако, Венесуэла. [26] с помощью аналогичных методов датирование экземпляра Glyptotherium из Таймы-Таймы до 12 580 ± 60 радиоуглеродных лет назад, [116] [117] хотя минимальная дата всего комплекса (~ 15 780 кал. BP, 12 980 ± 85 радиоуглеродных BP) более поздняя. [26] Как и в случае с другой вымершей плейстоценовой мегафауной, потенциальные причины вымирания включают человеческую охоту и изменение климата, связанное с холодным периодом раннего дриаса . [69]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Куатапаро, Дж. Н., и Рамирес, С. (1875). Описание ископаемого млекопитающего неизвестного вида, принадлежащего к роду «Glyptodon»: найдено среди посттретичных слоев Текискиака, в районе Зумпанго . Ф. Диас де Леон.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак Сурита, Альфредо Эдуардо; Джилетт, Дэвид Д.; Куадрелли, Франциско; Карлини, Альфредо Армандо (01 июня 2018 г.). «Повесть о двух кладах: сравнительное исследование Glyptodon Owen и Glyptotherium Osborn (Xenarthra, Cingulata, Glyptodontidae)» . Геобиос . 51 (3): 247–258. Бибкод : 2018Geobi..51..247Z . дои : 10.1016/j.geobios.2018.04.004 . hdl : 11336/83593 . ISSN 0016-6995 . S2CID 134450624 .
- ^ Jump up to: а б с д и Джилетт и Рэй, 1981 , с. 3.
- ^ Джилетт и Рэй 1981 , с. 16.
- ^ Феликс, JP (1899). Вклад в геологию и палеонтологию Мексиканской Республики (Том 1). А. Феликс.
- ^ Мальдонадо, М. (1948). Ископаемые позвоночные четвертичного периода Мексики. Журнал Мексиканского общества естественной истории, 9 (1-2): 1-35.
- ^ Jump up to: а б с д и Осборн, Генри Фэрфилд (1903). Glyptotherium Texanum: новый глиптодонт из нижнего плейстоцена Техаса . приказ попечителей Американского музея естественной истории.
- ^ Коуп, ЭД (1889). edentata Северной Америки. Американский натуралист , 23 (272), 657–664.
- ^ Лейди, Дж. (1889). Описание останков позвоночных из Пис-Крик, Флорида. Труды Свободного научного института Вагнера в Филадельфии , 2 , 19–31.
- ^ Джилетт и Рэй 1981 , с. 6.
- ^ Лейди, Д. (1889). Ископаемые позвоночные из Флориды. Труды Академии естественных наук Филадельфии , 96-97.
- ^ Jump up to: а б Джилетт и Рэй, 1981 , с. 14.
- ^ Jump up to: а б с д и ж Браун, Барнум (1912). «Брахиостракон, новый род глиптодонтов из Мексики» . Бюллетень АМНХ . 31 (17): 167–190. hdl : 2246/1402 .
- ^ Jump up to: а б с д и ж г час я дж Джилетт, Д.Д.; Рэй, CE (1981), Глиптодонты Северной Америки (PDF).
- ^ Хэй, ОП (1923). Плейстоцен Северной Америки и ее позвоночные животные из штатов к востоку от реки Миссисипи и из канадских провинций к востоку от долготы 95> О (№ 22). Институт Карнеги в Вашингтоне.
- ^ Джилетт и Рэй 1981 , с. 6 и 14.
- ^ Jump up to: а б с д и Симпсон, Г.Г., и Холмс, WW (1929). Плейстоценовая фауна млекопитающих поля семинолов, округ Пинеллас, Флорида . Бюллетень АМНХ; т. 56, статья 8.
- ^ Jump up to: а б с д Мид, GE (1953). Фауна позвоночных раннего плейстоцена из Фредерика, Оклахома. Геологический журнал , 61 (5), 452–460.
- ^ Гулд, Китай (1928). Ископаемый глиптодон в гравийных пластах Фредерик. В трудах Академии наук Оклахомы (стр. 148–150).
- ^ Спир, Лесли (10 февраля 1928 г.). «О древности человека во Фредерике, Оклахома» . Наука . 67 (1728): 160–161. Бибкод : 1928Sci....67..160S . дои : 10.1126/science.67.1728.160 . ISSN 0036-8075 . ПМИД 17752885 .
- ^ Jump up to: а б с д и ж г час я дж к л м Джилетт, Дэвид Д.; Карранса-Кастаньеда, Оскар; Уайт, Ричард С.; Морган, Гэри С.; Трэшер, Ларри К.; МакКорд, Роберт; Маккалоу, Гэвин (01 июня 2016 г.). «Онтогенез и половой диморфизм Glyptotherium texanum (Xenarthra, Cingulata) из плиоцена и плейстоцена (бланканский и ирвингтонский NALMA) Аризоны, Нью-Мексико и Мексики» . Журнал эволюции млекопитающих . 23 (2): 133–154. дои : 10.1007/s10914-015-9309-6 . ISSN 1573-7055 . S2CID 15219339 .
- ^ Jump up to: а б с д и ж г час я дж к Оливейра, Э. В., Порпино К.Д. и Баретто А. (2010). О присутствии Glyptotherium в позднем плейстоцене Северо-Востока Бразилии и статусе « Glyptodon » и « Chlamydotherium ». Палеобиогеографическое значение. Новогодний сборник трактатов по геологии и палонтологии , 258 (3), 353.
- ^ Jump up to: а б с д и Лесса, Карлос Микаэль Бонфим; ГОМЕС, Вероника Сантос; Черкинский, Александр; Дантас, Марио Андре Триндаде (2021). «Изотопная пеоэкология (δ13C, δ18O) двух комплексов мегамлекопитающих позднего плейстоцена из межтропического региона Бразилии» . Журнал южноамериканских наук о Земле . 112 : 103576. Бибкод : 2021JSAES.11203576L . дои : 10.1016/j.jsames.2021.103576 . ISSN 0895-9811 . S2CID 244181217 .
- ^ Jump up to: а б с Порпино, Клеберсон де О.; Ферникола, Хуан К.; Бергквист, Лилиан П. (18 мая 2010 г.). «Возвращаясь к внутритропическому бразильскому виду Hoplophorus euphractus (Cingulata, Glyptodontoidea) и филогенетическому сходству Hoplophorus» . Журнал палеонтологии позвоночных . 30 (3): 911–927. Бибкод : 2010JVPal..30..911P . дои : 10.1080/02724631003765735 . hdl : 11336/69074 . ISSN 0272-4634 . S2CID 83888299 .
- ^ Лунд, PW (1845): Conspectum-dasypodum. – Естественные науки и математические диссертации Датского королевского общества наук, 11: lxxxii-lxxxvi.
- ^ Jump up to: а б с д и ж г час я дж к л м КАРЛИНИ, Альфредо А.; Каррильо-Брисеньо, Хорхе Д.; Хаймес, Артуро; Агилера, Оранжель; Зурита, Альфредо Э.; ИРИАрте, Хосе; Санчес-Вильягра, Марсело Р. (16 июня 2022 г.). «Поврежденные остатки глиптодонтидов из стоянок позднего плейстоцена на северо-западе Венесуэлы: свидетельства охоты человека?» . Швейцарский журнал палеонтологии . 141 (1): 11. Бибкод : 2022SwJP..141...11C . дои : 10.1186/s13358-022-00253-3 . ISSN 1664-2384 . S2CID 249681104 .
- ^ Jump up to: а б с д Митчелл, К.Дж.; Сканферла, А.; Сойбельзон, Э.; Бонини, Р.; Очоа, Дж.; Купер, А. (2016). «Древняя ДНК вымершего южноамериканского гигантского глиптодонта Doedicurus sp. (Xenarthra: Glyptodontidae) показывает, что глиптодонты произошли от эоценовых броненосцев» . Молекулярная экология . 25 (14): 3499–3508. Бибкод : 2016MolEc..25.3499M . дои : 10.1111/mec.13695 . hdl : 11336/48521 . ПМИД 27158910 . S2CID 3720645 .
- ^ Jump up to: а б Дельсюк, Ф.; Гибб, GC; Куч, М.; Билле, Г.; Отье, Л.; Саутон, Дж.; Руйяр, Ж.-М.; Ферникола, JC; Вискайно, Сан-Франциско; Макфи, RDE; Пойнар, Х.Н. (2016). «Филогенетическое сходство вымерших глиптодонтов» . Текущая биология 26 (4): Р155–Р156. Бибкод : 2016CBio...26.R155D . дои : 10.1016/j.cub.2016.01.039 . hdl : 11336/49579 . ПМИД 26906483 .
- ^ Дельсюк, Ф.; Гибб, GC; Куч, М.; Билле, Г.; Отье, Л.; Саутон, Дж.; Руйяр, Ж.-М.; Ферникола, JC; Вискайно, Сан-Франциско; Макфи, RDE; Пойнар, HN (22 февраля 2016 г.). «Филогенетическое сходство вымерших глиптодонтов» (PDF) . Текущая биология 26 (4): Р155–Р156. Бибкод : 2016CBio...26.R155D . дои : 10.1016/j.cub.2016.01.039 . ПМИД 26906483 .
- ^ Jump up to: а б Куадрелли, Фрэнсис; Зурита, Альфред Э.; Булл, Пол; Миньо-Бойлини, Анхель Р.; Родригес-Буало, Джеймс; Переа, Даниэль; Коин Суарес, Габриэль Э. (03 сентября 2018 г.). «Позднеплейстоценовые Glyptodontinae (Mammalia, Xenarthra, Glyptodontidae) из южной части Южной Америки: комплексный обзор » Журнал палеонтологии позвоночных . 38 (5): e1525390. Бибкод : 2018JVPal..38E5390C . дои : 10.1080/02724634.2018.1525390 . ISSN 0272-4634 . S2CID 92335544 .
- ^ Jump up to: а б с Зурита, Альфредо Э.; Гонсалес Руис, Лауреано Р.; Гомес-Круз, Арли Дж.; Аренас-Москера, Хосе Э. (01 мая 2013 г.). «Наиболее полные известные неогеновые Glyptodontidae (Mammalia, Xenarthra, Cingulata) из северной части Северной Америки: таксономические, палеобиогеографические и филогенетические значения» . Журнал палеонтологии позвоночных . 33 (3): 696–708. Бибкод : 2013JVPal..33..696Z . дои : 10.1080/02724634.2013.726677 . hdl : 11336/2971 . ISSN 0272-4634 . S2CID 86099989 .
- ^ Jump up to: а б Зурита, Альфредо Э.; Миньо-Бойлини, Анхель Р.; Франция, Аналия; Аренас-Москера, Хосе Э. (31 декабря 2012 г.). «Плейстоценовые Glyptodontidae Grey, 1869 (Xenarthra: Cingulata) из Колумбии и некоторые соображения о южноамериканских Glyptodontinae» (PDF) . Бразильский журнал палеонтологии . 15 (3): 273–280. дои : 10.4072/rbp.2012.3.04 .
- ^ Jump up to: а б с Куадрелли, Франциско; Зурита, Альфредо Э.; Турино, Пол; Миньо-Бойлини, Анхель Р.; Переа, Даниэль; Луна, Карлос А.; Джилетт, Дэвид Д.; Медина, Омар (16 сентября 2020 г.). «Новый вид глиптодонтин (Mammalia, Xenarthra, Glyptodontidae) из четвертичного периода Восточных Кордильер, Боливия: филогения и палеобиогеография» . Журнал систематической палеонтологии . 18 (18): 1543–1566. Бибкод : 2020JSPal..18.1543C . дои : 10.1080/14772019.2020.1784300 . ISSN 1477-2019 . S2CID 221064742 .
- ^ Джилетт и Рэй 1981 , с. 2.
- ^ Джилетт и Рэй 1981 , с. 1.
- ^ Jump up to: а б Дантас, Марио Андре Триндаде; Черкинский, Александр; Лесса, Карлос Микаэль Бонфим; САНТОС, Лучано Вилабойм; Коссуоль, Марио Альберто; ОМЕНА, Эрика Кавальканте; СИЛЬВА, Хорхе Луис Лопес да; Сиал, Альсидес Нобрега; Бочеренс, Эрве (29 ноября 2018 г.). «Интегративная изотопная палеоэкология (δ13C, δ18O) сообщества позвоночных позднего плейстоцена из Сержипи, северо-восток Бразилии». п. 482752. bioRxiv 10.1101/482752 . S2CID 91321429
- ^ Jump up to: а б с дю Плесси, Антон; Броекховен, Крис; Ядройцев Игорь; Ядройцава, Инна; ле Ру, Стефан Герхард (01 июня 2018 г.). «Анализ защитного дизайна природы: бронежилет глиптодонта» . Журнал механического поведения биомедицинских материалов . 82 : 218–223. дои : 10.1016/j.jmbbm.2018.03.037 . ISSN 1751-6161 . ПМИД 29621689 . S2CID 4608467 .
- ^ Джилетт и Рэй 1981 , с. 200.
- ^ Джилетт и Рэй 1981 , с. 58.
- ^ Джилетт и Рэй 1981 , с. 39.
- ^ Jump up to: а б с Джилетт и Рэй, 1981 , с. 202.
- ^ Зурита, А.Э., Мине-Бойлини, А. Р., Сойбельзон Э., Карлини А.А. и Уоллс Риверс Ф. (2009). Разнообразие Glyptodontidae (Xenarthra, Cingulata) в долине Тарья (Боливия): систематические, биостратиграфические и палеобиогеографические аспекты отдельного комплекса. Новогодний сборник геологических и палеонтологических исследований , 251 (2), 225.
- ^ Зурита, А.Е.; Скарано, AC; Карлини, А.А.; Сциллато-Яне, Дж.Дж.; Сойбельзон, Э. (4 апреля 2011 г.). «Neosclerocalyptus spp. (Cingulata: Glyptodontidae: Hoplophorini): морфология черепа и палеоокружение в меняющемся четвертичном периоде» . Журнал естественной истории . 45 (15–16): 893–914. Бибкод : 2011JNatH..45..893Z . дои : 10.1080/00222933.2010.536917 . ISSN 0022-2933 . S2CID 85146482 .
- ^ Ферникола, Хуан Карлос; Толедо, Нестор; Барго, М. Сусана; Вискайно, Серджио Ф. (22 сентября 2012 г.). «Неоморфное окостенение носовых хрящей и строение системы околоносовых пазух глиптодонта Neosclerocalyptus Paula Couto 1957 (Mammalia, Xenarthra)» . Электронная палеонтология . 15 (3): 1–22. дои : 10.26879/333 . ISSN 1094-8074 .
- ^ Jump up to: а б с Сурита, Альфредо Эдуардо; Сойбельзон, Леопольдо Гектор; Сойбельзон, Эстебан; Гаспарини, Херман Мариано; Ашен, Маркос Мартин; Арзани, Эктор (1 января 2010 г.). «Дополнительные защитные структуры у Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae)» . Анналы палеонтологии . 96 (1): 1–11. Бибкод : 2010AnPal..96....1Z . дои : 10.1016/j.annpal.2010.01.001 . ISSN 0753-3969 .
- ^ Джилетт и Рэй 1981 , с. 169, 172 и 173.
- ^ Jump up to: а б с д и Амсон, Эли; Ньякатура, Джон А. (01 декабря 2018 г.). «Посткраниальная костно-мышечная система ксенартранов: результаты более чем двухвековых исследований и направления на будущее» . Журнал эволюции млекопитающих . 25 (4): 459–484. дои : 10.1007/s10914-017-9408-7 . ISSN 1573-7055 . S2CID 254693335 .
- ^ Jump up to: а б с Оуэн, Ричард (1841). «VI. — Описание зуба и части скелета Glyptodon clavipes, большого четвероногого отряда неполнозубых, к которому принадлежит мозаичный костный панцирь, описанный и изображенный г-ном Клифтом в бывшем томе «Трудов геологического общества». Общества с рассмотрением вопроса, обладал ли Мегатерий аналогичным кожным доспехом» . Труды Лондонского геологического общества . 6 (1): 81–106. дои : 10.1144/transgslb.6.1.81 . ISSN 2042-5295 .
- ^ Сенешаль, DL (1865). Обратите внимание на панцирь или дермато-скелет и зубную систему Glyptodon clavipes, а также на биологические особенности этого животного, установленные на основе изучения его ископаемых останков . Balitout, Questroy et Cie.
- ^ Фари, РА, и Вискайно, Сан-Франциско (1997). Аллометрия костей современных и вымерших броненосцев (Xenarthra, Dasypoda). Zeitschrift Fur Saugetierkunde , 62 , 65-70.
- ^ Фаринья, Ричард А.; Вискайно, Серджио Ф.; Бланко, Р. Эрнесто (21 апреля 1997 г.). «Шкалирование показателя спортивных способностей ископаемых и современных наземных четвероногих» . Журнал теоретической биологии . 185 (4): 441–446. Бибкод : 1997JThBi.185..441F . дои : 10.1006/jtbi.1996.0323 . ISSN 0022-5193 .
- ^ Шокей, Би Джей (2001). Специализированные коленные суставы у некоторых вымерших эндемичных травоядных животных Южной Америки. Acta Palaeontologica Polonica , 46 (2).
- ^ Jump up to: а б Рассел, Дейл А.; Рич, Фредрик Дж.; Шнайдер, Винсент; Линч-Штиглиц, Жан (май 2009 г.). «Теплый термальный анклав в позднем плейстоцене на юго-востоке США» . Биологические обзоры . 84 (2): 173–202. дои : 10.1111/j.1469-185X.2008.00069.x . ISSN 1464-7931 . ПМИД 19391200 . S2CID 9609391 .
- ^ Jump up to: а б с д и ж Перес-Креспо, Вирджиния, Арройо-Кабралес, Дж., Альва-Вальдивия, Л.М., Моралес-Пуэнте, П., и Сьенфуэгос-Альварадо, Э. (2012). Определения диеты и среды обитания мексиканских глиптодонтов из Седрала (Сан-Луис-Потоси, Мексика) на основе анализа стабильных изотопов. Геологический журнал , 149 (1), 153-157.
- ^ Jump up to: а б Кили, Дж. Э., и Рандел, П. В. (2003). Эволюция механизмов концентрации углерода CAM и C4. Международный журнал наук о растениях , 164 (S3), S55-S77.
- ^ Jump up to: а б ОМЕНА, Эрика Кавальканте; СИЛЬВА, Хорхе Луис Лопес да; Сиал, Альсидес Нобрега; Черкинский, Александр; Дантас, Марио Андре Триндаде (3 октября 2021 г.). «Мезо-мегатравоядные животные позднего плейстоцена из межтропического региона Бразилии: изотопная диета (δ13C), дифференциация никеля, гильдии и реконструкция палеосреды (δ13C, δ18O)» . Историческая биология . 33 (10): 2299–2304. Бибкод : 2021HBio...33.2299O . дои : 10.1080/08912963.2020.1789977 . ISSN 0891-2963 . S2CID 225543776 .
- ^ Дантас, МАТ, Черкинский, А., Лесса, КМБ, Сэйнтс, Л.В., Козцуоль, М.А., Омена, Э. К., ... и Бочеренс, Х. (2020). Изотопная палеоэкология (δ13C, δ18O) сообщества позвоночных позднего плейстоцена из Межтропического региона Бразилии. Бразильский журнал палеонтологии , 23 (2), 138–152.
- ^ Филлип Э. Джардин, Кристин М. Дженис, Сарда Сэни, Майкл Дж. Бентон. «Песчаник, а не трава: согласованные модели раннего происхождения гипсодонтии у копытных Великих равнин и глайров». Палеогеография, Палеоклиматология, Палеоэкология. Декабрь 2012 г.: 365–366, 1–10.
- ^ Де Юлиис, Херардо; Барго, Мария С.; Вискайно, Серхио Ф. (19 января 2001 г.). «Изменения в морфологии черепа и жевании у ископаемых гигантских броненосцев Pampatherium spp. и родственных родов (Mammalia: Xenarthra: Pampatheriidae) с комментариями к их систематике и распространению» . Журнал палеонтологии позвоночных . 20 (4): 743–754. doi : 10.1671/0272-4634(2000)020[0743:VISMAM]2.0.CO;2 . ISSN 0272-4634 . S2CID 86263866 .
- ^ Вискайно, Серджио Ф.; Кассини, Гильермо Х.; Ферникола, Хуан К.; Барго, М. Сусана (2011). «Оценка среды обитания и привычек питания с помощью экоморфологических особенностей глиптодонтов (млекопитающих, ксенартры)». Амегинян: 305–319. дои : 10.5710/AMGH.v48i3(364) . S2CID 85793531 . Проверено 29 октября 2015 г.
- ^ Вискайно, С.Ф., Фаринья, Р.А., и Маццетта, Г.В. (1999). Ульнарные размеры и фоссориальность у броненосцев. Акта Териологика ,
- ^ Картер, Т.С. и Энкарнасао, CD (1983). Характеристики и использование нор четырьмя видами броненосцев в Бразилии. Журнал маммологии , 64 (1), 103-108.
- ^ Вискайно, Серджио Ф.; Фаринья, Ричард А. (29 марта 2007 г.). «Питание и передвижение броненосца Peltephilus: новый взгляд» . Летайя . 30 (1): 79–86. дои : 10.1111/j.1502-3931.1997.tb00446.x .
- ^ Сааринен, Юха; Карме, Алексис (15 июня 2017 г.). «Износ зубов и рацион современных и ископаемых ксенартранов (Mammalia, Xenarthra) – применение нового подхода к мезоизносу» . Палеогеография, Палеоклиматология, Палеоэкология . 476 : 42–54. Бибкод : 2017PPP...476...42S . дои : 10.1016/j.palaeo.2017.03.027 . ISSN 0031-0182 .
- ^ Вискайно, Серхио Ф.; Бланко, Р. Эрнесто; Бендер, Дж. Бенджамин; Милн, Ник (2011). «Пропорции и функции конечностей глиптодонтов: Конечности глиптодонтов» . Летайя . 44 (1): 93–101. Бибкод : 2011Лета..44...93В . дои : 10.1111/j.1502-3931.2010.00228.x .
- ^ Чен, Ирен Х.; Кианг, Джеймс Х.; Корреа, Виктор; Лопес, Мария И.; Чен, По-Ю; МакКиттрик, Джоанна; Мейерс, Марк А. (1 июля 2011 г.). «Броня броненосца: механические испытания и оценка микроструктуры» . Журнал механического поведения биомедицинских материалов . Спецвыпуск по природным материалам / Доклады Третьей международной конференции по механике биоматериалов и тканей. 4 (5): 713–722. дои : 10.1016/j.jmbbm.2010.12.013 . ISSN 1751-6161 . ПМИД 21565719 .
- ^ Барбоза, Фернандо Энрике де Соуза; Порпино, Клеберсон де Оливейра; Фрагозо, Ана Бернадет Лима; Оливейра, Эдисон Висенте (13 февраля 2014 г.). «Артрит у глиптодонта (Mammalia, Xenarthra, Cingulata)» . ПЛОС ОДИН . 9 (2): e88646. Бибкод : 2014PLoSO...988646B . дои : 10.1371/journal.pone.0088646 . ISSN 1932-6203 . ПМЦ 3923812 . ПМИД 24551126 .
- ^ ЛИМА, Фабио Кунья Гимарайнш де; Порпино, Клеберсон де Оливейра (18 октября 2018 г.). «Эктопаразитизм и инфекции в экзоскелетах ископаемых крупных поясных извилины» . ПЛОС ОДИН . 13 (10): e0205656. Бибкод : 2018PLoSO..1305656D . дои : 10.1371/journal.pone.0205656 . ISSN 1932-6203 . ПМК 6193641 . ПМИД 30335796 .
- ^ Jump up to: а б с д и Фейт, Дж. Тайлер; Суровелл, Тодд А. (08 декабря 2009 г.). «Синхронное вымирание млекопитающих плейстоцена Северной Америки» . Труды Национальной академии наук . 106 (49): 20641–20645. Бибкод : 2009PNAS..10620641F . дои : 10.1073/pnas.0908153106 . ISSN 0027-8424 . ПМК 2791611 . ПМИД 19934040 .
- ^ Репеннинг, Чарльз А. (1962). « Гигантский суслик Paenemarmota ». Журнал палеонтологии . 36 (93): 540–556. JSTOR 1301086.
- ^ Jump up to: а б Далквест, WW (1975). Окаменелости позвоночных из местной фауны Бланко в Техасе.
- ^ Мартин, Л.Д., Шульц, CB, и Шульц, MR (1988). Саблезубые кошки из плио-плейстоцена Небраски.
- ^ Федучча, Дж. Алан; Форд, Норман Л. (1 октября 1970 г.). «Некоторые хищные птицы из верхнего плиоцена Канзаса» . Аук . 87 (4): 795–797. дои : 10.2307/4083714 . ISSN 1938-4254 . JSTOR 4083714 .
- ^ Маршалл, Уильям Х. (1 января 1937 г.). «Двухгребневый баклан, гнездящийся в заповеднике Медвежья река в штате Юта» . Кондор . 39 (1): 36. дои : 10.2307/1363487 . ISSN 1938-5129 . JSTOR 1363487 .
- ^ Jump up to: а б с Давила, С. Лорена; Стиннесбек, Сара Р.; Гонсалес, Сильвия; Линдауэр, Сюзанна; Эскамилла, Хуан; Стиннесбек, Вольфганг (01 сентября 2019 г.). «Позднеплейстоценовая (ранчолабрейская) фауна Гватемалы: пересмотр и интерпретация» . Четвертичные научные обзоры . 219 : 277–296. Бибкод : 2019QSRv..219..277D . doi : 10.1016/j.quascirev.2019.07.011 . ISSN 0277-3791 . S2CID 201306819 .
- ^ Стиннесбек, СР (2020). Мексиканские ископаемые наземные ленивцы - тематическое исследование оборота мегафауны позднего плейстоцена в Мексиканском коридоре.
- ^ Jump up to: а б с Картель, Кастор; Хартвиг, WC (1996). «Новый вымерший примат среди плейстоценовой мегафауны Баии, Бразилия». Труды Национальной академии наук. 93 (13): 6405–6409.
- ^ Айзенберг, Джон Ф.; Редфорд, Кент Х. (1989). Млекопитающие Неотропов, Том 3: Эквадор, Боливия, Бразилия. Издательство Чикагского университета. п. 247. ISBN 978-0-226-19542-1 .
- ^ Халенар, Лорен Б. (декабрь 2011 г.). «Реконструкция локомоторного репертуара Protopithecus brasiliensis». Анатомическая запись. 294 (12): 2048–2063.
- ^ Jump up to: а б Дэвид Уэбб, С. (23 августа 2006 г.). «Великий американский биотический обмен: закономерности и процессы1» . Анналы ботанического сада Миссури . 93 (2): 245–257. doi : 10.3417/0026-6493(2006)93[245:TGABIP]2.0.CO;2 . ISSN 0026-6493 . S2CID 198152030 .
- ^ Чарриер, Джон Дж. Флинн, Андре Р. Висс и Рейнальдо (2007). «Пропавшие млекопитающие Южной Америки» . Научный американец . 296 (5): 68–75. Бибкод : 2007SciAm.296e..68F . doi : 10.1038/scientificamerican0507-68 . ПМИД 17500416 . Проверено 9 мая 2023 г.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Блох, Дж.И., Вудрафф, Э.Д., Вуд, А.Р., Ринкон, А.Ф., Харрингтон, А.Р., Морган, Г.С., ... и Макфадден, Б.Дж. (2016). Первая ископаемая обезьяна Северной Америки и тропический биотический обмен раннего миоцена. Природа , 533 (7602), 243-246.
- ^ Крофт, Д.А. (2016). Рогатые броненосцы и рафтинговые обезьяны: удивительные ископаемые млекопитающие Южной Америки . Издательство Университета Индианы.
- ^ Морган, GS (2005). Великий американский биотический обмен во Флориде. Бюллетень Музея естественной истории Флориды , 45 (4), 271–311.
- ^ Макдональд, Х.Г. (2005). Палекология вымерших ксенартранов и Великий американский биотический обмен. Бюллетень Музея естественной истории Флориды , 45 (4), 319–340.
- ^ Jump up to: а б Сциллато-Ян, Дж.Дж.; Карлини, А.А.; Тонни, EP; Норьега, Дж.И. (1 октября 2005 г.). «Палеобиогеография позднеплейстоценовых пампатеров Южной Америки » Журнал южноамериканских наук о Земле . Четвертичная палеонтология и биостратиграфия юга Южной Африки. 20 (1): 131–138. Бибкод : 2005JSAES..20..131S . дои : 10.1016/j.jsames.2005.06.012 . hdl : 11336/80762 . ISSN 0895-9811 .
- ^ Jump up to: а б с Карлини, Альфредо А.; Зурита, Альфредо Э.; Агилера, Оранжель А. (2008). «Североамериканские глиптодонтины (Xenarthra, Mammalia) в верхнем плейстоцене северной части Южной Америки» . Палеонтологический журнал . 82 (2): 125–138. Бибкод : 2008PalZ...82..125C . дои : 10.1007/BF02988404 . eISSN 1867-6812 . ЛПВП : 11336/36844 . ISSN 0031-0220 . S2CID 128528271 .
- ^ Jump up to: а б Сьоне А.Л., Гаспарини Г.М., Сойбельзон Э., Сойбельзон Л.Х. и Тонни Э.П. (2015). Великий американский биотический обмен: взгляд из Южной Америки (стр. 97). Дордрехт: Спрингер.
- ^ Вудберн, Миссури (2010). Великий американский биотический обмен: расселение, тектоника, климат, уровень моря и загоны для содержания. Журнал эволюции млекопитающих , 17 , 245–264.
- ^ Лунделиус, Эрнест Л.; Брайант, Вон М.; Мандель, Рольф; Тис, Кеннет Дж.; Томс, Олстон (1 января 2013 г.). «Первое появление токсодонта (Mammalia, Notoungulata) в США» . Журнал палеонтологии позвоночных . 33 (1): 229–232. Бибкод : 2013JVPal..33..229L . дои : 10.1080/02724634.2012.711405 . hdl : 1808/13587 . ISSN 0272-4634 . S2CID 53601518 .
- ^ Jump up to: а б Зурита, Альфред Э.; Карлини, Альфред А.; Джилетт, Дэвид; Санчес, Родольфо (01 марта 2011 г.). «Позднеплиоценовые Glyptodontinae (Xenarthra, Cingulata, Glyptodontidae) Южной и Северной Америки: морфология и палеобиогеографические последствия в GABI» . Журнал южноамериканских наук о Земле . 31 (2): 178–185. Бибкод : 2011JSAES..31..178Z . дои : 10.1016/j.jsames.2011.02.001 . hdl : 11336/41714 . ISSN 0895-9811 .
- ^ Пухос Ф. и Салас Р. (2004). Систематическая переоценка и палеогеографический обзор ископаемой Ксенартры из Перу. Bulletin de l'Institut français d'études andines , (33 (2)), 331-377.
- ^ Распространение, Джордж; Коронато, Андреа М.; Салем, Моника (1 октября 2005 г.). «Хронология позднекайнозойских патагонских оледенений и их корреляция с биостратиграфическими подразделениями Пампейской области (Аргентина)» . Журнал южноамериканских наук о Земле . Четвертичная палеонтология и биостратиграфия юга Южной Африки. 20 (1): 81–103. Бибкод : 2005JSAES..20...81R . дои : 10.1016/j.jsames.2005.07.004 . hdl : 11336/150967 . ISSN 0895-9811 .
- ^ Энтони, Морис (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. стр. 100-1 203–204. ISBN 978-0-253-01042-1 .
- ^ Джилетт, Д.Д. (весна 2010 г.). «Глиптодонты в Аризоне» . Геология Аризоны . Геологическая служба Аризоны . Проверено 17 августа 2018 г.
- ^ Jump up to: а б Брандес, Г. (1900): О причине вымирания делювиальных млекопитающих. Коррблатт Д. немецкий. Гес ф. Антрополь. 31 год, Мюнхен, 1901 год.
- ^ Jump up to: а б Болин, Б. (1940). 8. Пищевые привычки махайродонтов, особенно смилодона .
- ^ Араужо-Жуниор, Эрминиу Исмаэль де; БАРБОЗА, Фернандо Энрике де Соуза; СИЛЬВА, Лукас Энрике Медейрос да (15 февраля 2017 г.). «Перекрывающаяся палеоихнология, палеоэкология и тафономия: анализ следов зубов в мегафаунистическом комплексе позднего плейстоцена-раннего голоцена Бразилии и описание нового ихнотаксона в твердом субстрате» . Палеогеография, Палеоклиматология, Палеоэкология . 468 : 122–128. Бибкод : 2017PPP...468..122A . дои : 10.1016/j.palaeo.2016.12.007 . ISSN 0031-0182 .
- ^ Крук, Уилсон В.; Харрис, РК (1958). «Лагерь плейстоцена недалеко от Льюисвилля, штат Техас» . Американская древность . 23 (3): 233–246. дои : 10.2307/276304 . ISSN 0002-7316 . JSTOR 276304 . S2CID 161227132 .
- ^ Вагеспак, Николь М.; Суровелл, Тодд А. (2003). «Стратегии охоты на Кловиса, или Как заработать на обильных ресурсах» . Американская древность . 68 (2): 333–352. дои : 10.2307/3557083 . ISSN 0002-7316 . JSTOR 3557083 . S2CID 164047864 .
- ^ Фогт, К. (1881). Скелет человека, связанный с глиптодонтидами. Бюллетень Парижского общества антропологии, 3 (4), 693–699.
- ^ Киньонес, София И.; О королях, Мартин; Зурита, Альфред Э.; Куадрелли, Фрэнсис; Миньо-Бойлини, Анхель Р.; Пуаре, Даниэль Г. (01 ноября 2020 г.). «Неосклерокалиптус Паула Коуто (Xenarthra, Glyptodontidae) в позднем плиоцене-раннем плейстоцене Пампейского региона (Аргентина): его вклад в понимание эволюционной истории плейстоценовых глиптодонтов» . Журнал южноамериканских наук о Земле . 103 : 102701. Бибкод : 2020JSAES.10302701Q . doi : 10.1016/j.jsames.2020.102701 . ISSN 0895-9811 . S2CID 225024450 .
- ^ Политис, Густаво Г.; Мессинео, Пабло Г.; Стаффорд, Томас В.; Линдси, Эмили Л. (2019). «Кампо-Лаборде: место убийства и разделки гигантских наземных ленивцев позднего плейстоцена в пампасах» . Достижения науки . 5 (3): eaau4546. Бибкод : 2019SciA....5.4546P . дои : 10.1126/sciadv.aau4546 . ISSN 2375-2548 . ПМК 6402857 . ПМИД 30854426 .
- ^ Пратес, Лучано; Перес, С. Иван (12 апреля 2021 г.). «Вымирание мегафауны Южной Америки в позднем плейстоцене, связанное с ростом численности рыбьего хвоста и численности населения» . Природные коммуникации . 12 (1): 2175. Бибкод : 2021NatCo..12.2175P . дои : 10.1038/s41467-021-22506-4 . ISSN 2041-1723 . ПМК 8041891 . ПМИД 33846353 .
- ^ Jump up to: а б с Олбрайт, Л.; Сандерс, Альберт; Уимс, Роберт; Чичимурри, Дэвид; Найт, Джеймс (31 октября 2019 г.). «Биостратиграфия кайнозойских позвоночных Южной Каролины, США, и дополнения к фауне» . Бюллетень Музея естественной истории Флориды . 57 (2): 77–236. дои : 10.58782/flmnh.qqgg4577 . ISSN 2373-9991 .
- ^ Сандерс, AE (2002). Дополнения к фауне млекопитающих плейстоцена Южной Каролины, Северной Каролины и Джорджии . Американское философское общество.
- ^ Jump up to: а б Валерио, Алабама, и Лаурито, Калифорния (2011). Летопись окаменелостей Glyptotherium floridanum (Xenarthra, Cingulata, Glyptodontidae) в четвертичном периоде Коста-Рики, Центральная Америка. Геологический журнал Центральной Америки , (45), 141–145.
- ^ Джексон, Д.Р., и Фернандес, Э. (2005). Небольшая мегафауна плейстоценовых млекопитающих из южного Гондураса. Бюллетень Музея естественной истории Флориды , 45 (4), 261–269.
- ^ Сиснерос, JC (2005). Новая фауна позвоночных плейстоцена из Сальвадора. Бразильский журнал палеонтологии , 8 (3), 239–255.
- ^ Лукас, Спенсер Г. (2014). «Млекопитающие позднего плейстоцена из Эль-Атильо, Панама» . Геологический журнал Центральной Америки (50): 139–151. ISSN 0256-7024 .
- ^ Валерио, Ана Л.; Лаурито, Сезар А. (2011). «Летопись окаменелостей Glyptotherium Floridanum (Xenarthra, Cingulata, Glyptodontidae) в четвертичном периоде Коста-Рики, Центральная Америка» . Геологический журнал Центральной Америки (на испанском языке) (45): 141–145. ISSN 0256-7024 .
- ^ Куадрелли, Франциско; Эскамилла, Джон; Сурита, Альфредо; Джилетт, Дэвид Д.; Давила, Лорена С. (22 августа 2023 г.). «Glyptotherium cylindricum (Cingulata, Glyptodontidae) из позднего плейстоцена Гватемалы: наиболее полная запись Glyptodontinae из Центральной Америки» . Алчеринга: Австралазийский журнал палеонтологии . 47 (3): 336–347. Бибкод : 2023Alch...47..336C . дои : 10.1080/03115518.2023.2242440 . ISSN 0311-5518 . S2CID 261137459 .
- ^ Jump up to: а б Рамирес-Крус, Гонсало А.; Монтеллано-Баллестерос, Марисоль (2014). «Две новые находки глиптодонтов (Mammalia: Cingulata) из позднего плейстоцена Тамаулипаса и Тласкалы, Мексика: значение для таксономии рода Glyptotherium» . Юго-западный натуралист . 59 (4): 522–530. дои : 10.1894/JKF-45.1 . ISSN 0038-4909 . S2CID 85750160 .
- ^ Хейнс, Гэри (8 февраля 2013 г.). «Вымирание в позднеледниковых ландшафтах Северной Америки» . Четвертичный интернационал . Заселение последних новых миров: первая колонизация Сахула и Америки. 285 : 89–98. Бибкод : 2013QuInt.285...89H . дои : 10.1016/j.quaint.2010.07.026 . ISSN 1040-6182 .
- ^ Стюарт, Энтони Джон (май 2015 г.). «Позднечетвертичное вымирание мегафауны на континентах: краткий обзор» . Геологический журнал . 50 (3): 338–363. Бибкод : 2015GeolJ..50..338S . дои : 10.1002/gj.2633 . ISSN 0072-1050 . S2CID 128868400 .
- ^ Карлини, А.А.; Зурита, AE (2006). «Glyptotherium Osborn (Mammalia, Xenarthra, Cingulata) в позднем плейстоцене Венесуэлы». 9-й Аргентинский конгресс палеонтологии и биостратиграфии, Кордова : 98.
- ^ ФАРИЯ, Фабио Энрике Кортес; Киношита, Анжела; Карвалью, Исмар де Соуза; Араужо-Жуниор, Эрминиу Исмаэль де; Пегорин, Присцилла; Мария Дж. Фигейредо, Ана; Баффа, Освальдо (01 августа 2020 г.). «Находка ERR окаменелостей мегафауны позднечетвертичного периода из Жоао Дурадо, Баия, Бразилия» . Журнал южноамериканских наук о Земле . 101 : 102586. Бибкод : 2020JSAES.10102586F . doi : 10.1016/j.jsames.2020.102586 . ISSN 0895-9811 . S2CID 216519652 .
Роды броненосцев и их вымершие союзники |
|---|



- Доисторические поясные мышцы
- Доисторические плацентарные роды
- Плиоценовые млекопитающие Северной Америки
- Плейстоценовые ксенартраны
- Плейстоценовые млекопитающие Северной Америки
- Первые выступления Занклина
- Голоценовые вымирания
- Бланкан
- Ирвингтонский
- Лужанский
- Ранчолабриан
- Плейстоцен Бразилия
- Голоценовая Бразилия
- Окаменелости Бразилии
- Плейстоцен Коста-Рика
- Окаменелости Коста-Рики
- Плейстоцен Сальвадор
- Окаменелости Сальвадора
- Плейстоценовая Мексика
- Окаменелости Мексики
- Плейстоценовая Панама
- Окаменелости Панамы
- Неоген США
- Плейстоцен США
- Окаменелости Соединенных Штатов
- Плейстоценовые млекопитающие Южной Америки
- Плейстоценовая Венесуэла
- Окаменелости Венесуэлы
- Ископаемые таксоны описаны в 1903 году.
- Таксоны, названные Генри Фэйрфилдом Осборном