Хамелеон
| Хамелеоны Временной диапазон: ранний миоцен – настоящее время, среднего палеоцена Происхождение | |
|---|---|
 | |
| По часовой стрелке сверху слева: Chamaeleon , Calumma parsonii , Chamaeleo namaquensis , Trioceros jacksonii , Furcifer pardalis и Brookesia micra. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Заказ: | Чешуйчатый |
| Подотряд: | Игуания |
| Клэйд : | Акродонта |
| Семья: | Хамелеониды Рафинеск , 1815 г. |
| Роды | |
 | |
| Родной ареал Chamaeleonidae | |
Хамелеоны или хамелеоны ( семейство Chamaeleonidae ) — самобытная и узкоспециализированная клада Старого Света ящериц , насчитывающая 200 видов, описанных по состоянию на июнь 2015 года. [1] Представители этого семейства наиболее известны своей ярко выраженной цветовой гаммой и способностью изменять цвет камуфляжа . Большое количество видов этого семейства демонстрируют значительную вариабельность способности менять цвет. Для кого-то это скорее изменение яркости (оттенков коричневого); у других можно увидеть множество цветовых комбинаций (красный, желтый, зеленый, синий).
Хамелеонов также отличают зигодактильные ноги, цепкий хвост, сжатое с боков тело, головные шлемы, языки-снаряды, используемые для ловли добычи, покачивающаяся походка. [2] а у некоторых видов на лбу и морде имеются гребни или рога. Глаза хамелеонов независимо подвижны, и поэтому мозг хамелеона постоянно анализирует два отдельных, индивидуальных изображения окружающей среды. При охоте на добычу глаза скоординированно фокусируются вперед, обеспечивая животному стереоскопическое зрение .
Хамелеоны ведут дневной образ жизни и приспособлены для визуальной охоты на беспозвоночных, преимущественно насекомых, хотя крупные виды могут ловить и мелких позвоночных. Хамелеоны обычно ведут древесный образ жизни , но есть также много видов, живущих на земле. Древесные виды используют свой цепкий хвост как дополнительную точку опоры, когда они движутся или отдыхают на деревьях или кустах; из-за этого их хвост часто называют «пятой конечностью». В зависимости от вида они варьируются от тропических лесов до пустынь и от низин до высокогорья, при этом подавляющее большинство встречается в Африке (около половины видов обитает только на Мадагаскаре ), но есть один вид в южной Европе и несколько в южной части Европы. Азия вплоть до Индии и Шри-Ланки . Они были завезены на Гавайи и во Флориду . [1] [3]
Этимология

Английское слово хамелеон ( / kə ˈ n m iː l i ə / / kuh-MEEL-ee-un / k ə ˈ m i l . j ə n , kuh -MEEL-yuhn ) представляет собой упрощенное написание латинского chamaeleōn , [4] заимствование греческого χαμαιλέων ( хамаилён ), [5] соединение ) « на χαμαί » ( хамаи земле [6] и leōn ( leōn ) «лев». [7] [8] [9]
Классификация
В 1986 году семейство Chamaeleonidae было разделено на два подсемейства: Brookesiinae и Chamaeleoninae . [10] В рамках этой классификации к Brookesiinae вошли роды Brookesia и Rampholeon , а также позднее отколовшиеся от них роды ( Palleon и Rieppeleon ), а к Chamaeleoninae — роды Bradypodion , Calumma , Chamaeleo , Furcifer и Trioceros , а также роды, позже отделившиеся от них. от них ( Архай , Надзикамбия и Киньонгия ). Однако с тех пор обоснованность обозначения этого подсемейства стала предметом многочисленных споров. [11] хотя большинство филогенетических исследований подтверждают представление о том, что карликовые хамелеоны подсемейства Brookesiinae не являются монофилетической группой. [12] [13] [14] [15]
Хотя некоторые авторитеты ранее предпочитали использовать эту подсемейную классификацию на основании принципа отсутствия доказательств, [11] позже эти власти отказались от этого подсемейства, больше не признавая никаких подсемейств семейства Chamaeleonidae. [16]
Однако в 2015 году Гло переработал подсемейство, поместив только роды Brookesia и Palleon в подсемейство Brookesiinae , а все остальные роды были помещены в Chamaeleoninae . [1]
Изменение цвета
Некоторые виды хамелеонов способны менять окраску кожи . Различные виды хамелеонов могут варьировать свою окраску и рисунок , комбинируя розовый, синий, красный, оранжевый, зеленый, черный, коричневый, голубой, желтый, бирюзовый и фиолетовый цвета. [17] Кожа хамелеона имеет поверхностный слой, содержащий пигменты, а под этим слоем находятся клетки с очень маленькими (наноразмерными) гуанина кристаллами . Хамелеоны меняют цвет, «активно настраивая фотонный отклик решетки маленьких нанокристаллов гуанина в s-иридофоре». [18] Эта настройка с помощью неизвестного молекулярного механизма меняет длину волны света, отраженного от кристаллов, что меняет цвет кожи. Изменение цвета было воспроизведено ex vivo путем изменения осмолярности кусочков белой кожи. [18]

(а) Для двух самцов (m1 и m2) показано обратимое изменение цвета: во время возбуждения (белые стрелки) фон кожи смещается от исходного состояния (зеленый) к желтому/оранжевому, а также сдвигаются как вертикальные полосы, так и горизонтальные полосы в середине тела. от синего до белесого (м1). У некоторых животных (м2) синие вертикальные полосы покрыты красными пигментными клетками.
(б) Красные точки: временная эволюция диаграммы цветности CIE третьего мужчины с зеленой кожей на видео высокого разрешения; пунктирная белая линия: оптический отклик при численном моделировании с использованием гранецентрированной кубической (FCC) решетки кристаллов гуанина с параметром решетки, указанным черными стрелками.
(c) Окрашивание гематоксилином и эозином поперечного среза белой кожи, показывающее эпидермис (ep) и два толстых слоя иридофоров.
(г) ПЭМ-изображения нанокристаллов гуанина в S-иридофоре в возбужденном состоянии и трехмерная модель ГЦК-решетки (показаны в двух ориентациях).
(д) ПЭМ-изображение нанокристаллов гуанина в D-иридофоре.
Масштабные линейки, 20 мм (в); 200 нм (г, д). [18]
Изменение цвета у хамелеонов выполняет маскировочную функцию , но чаще всего — социальную передачу сигналов и реакцию на температуру и другие условия. Относительная важность этих функций варьируется в зависимости от обстоятельств, а также от вида. Изменение цвета сигнализирует другим хамелеонам о физиологическом состоянии и намерениях хамелеона. [19] [20] Поскольку хамелеоны являются экзотермическими животными , еще одной причиной, по которой они меняют цвет, является регулирование температуры своего тела: либо на более темный цвет, чтобы поглощать свет и тепло, чтобы повысить свою температуру, либо на более светлый цвет, чтобы отражать свет и тепло, тем самым либо стабилизируя, либо понижая свою температуру. температура тела. [21] [22] Хамелеоны склонны проявлять более яркие цвета, когда проявляют агрессию к другим хамелеонам. [23] и более темные цвета, когда они подчиняются или «сдаются». [24] Большинство родов хамелеонов (исключение составляют Chamaeleo , Rampholeon и Rieppeleon ) имеют синюю флуоресценцию с видоспецифичным рисунком в бугорках черепа , а у Brookesia она также имеется в бугорках на теле. Флуоресценция исходит от костей, которые покрыты очень тонкой кожей, и, возможно, играет сигнальную роль, особенно в затененных местах обитания. [25]
Некоторые виды, такие как карликовый хамелеон Смита и несколько других представителей рода Bradypodion , корректируют свою окраску для маскировки в зависимости от зрения конкретного вида хищников (например, птиц или змей), со стороны которых им угрожает опасность. [26] [27] В интродуцированной гавайской популяции хамелеона Джексона заметные изменения цвета, которые используются для общения между хамелеонами, увеличились, тогда как изменения цвета камуфляжа против хищников уменьшились по сравнению с местной популяцией в Кении, где хищников больше. [28]
В коже хамелеонов есть два наложенных друг на друга слоя, которые контролируют их цвет и терморегуляцию. Верхний слой содержит решетку нанокристаллов гуанина, и, возбуждая эту решетку, можно манипулировать расстоянием между нанокристаллами, что, в свою очередь, влияет на то, какие длины волн света отражаются, а какие поглощаются. Возбуждение решетки увеличивает расстояние между нанокристаллами, и кожа отражает более длинные волны света. Таким образом, в расслабленном состоянии кристаллы отражают синий и зеленый цвета, а в возбужденном состоянии отражаются более длинные волны, такие как желтый, оранжевый, зеленый и красный. [29]
Кожа хамелеона также содержит некоторые желтые пигменты, которые в сочетании с синим, отражаемым расслабленной кристаллической решеткой, дают характерный зеленый цвет, который свойствен многим хамелеонам в расслабленном состоянии. Цветовые палитры хамелеона развивались в результате эволюции и окружающей среды. Хамелеоны, живущие в лесу, имеют более четкую и красочную палитру по сравнению с хамелеонами, живущими в пустыне или саванне, у которых более простая, коричневая и обугленная палитра. [30]
Эволюция
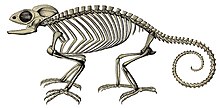
Самый старый описанный хамелеон — Anqingosaurus brevicephalus из среднего палеоцена (около 58,7–61,7 млн лет назад ) из Китая. [31] Другие окаменелости хамелеонов включают Chamaeleo caroliquarti из нижнего миоцена (около 13–23 млн лет назад) в Чехии и Германии и Chamaeleo intermedius из верхнего миоцена (около 5–13 млн лет назад) в Кении. [31]
Хамелеоны, вероятно, намного старше этого возраста и, возможно, имеют общего предка с игуанидами и агамидами, возраст которых превышает 100 млн лет назад (агамиды более близки). Поскольку окаменелости были найдены в Африке, Европе и Азии, хамелеоны, безусловно, когда-то были более распространены, чем сегодня.
Хотя почти половина всех видов хамелеонов сегодня обитает на Мадагаскаре, это не дает оснований предполагать, что хамелеоны могли произойти оттуда. [32] Фактически, недавно было показано, что хамелеоны, скорее всего, произошли из материковой Африки. [15] Похоже, существовало две отдельные океанические миграции с материка на Мадагаскар. Было высказано предположение, что разнообразие видов хамелеонов напрямую отражает увеличение открытых местообитаний (саванны, луга и пустоши), которое сопровождало период олигоцена. Монофилия семьи подтверждается несколькими исследованиями. [33]
Даза и др. (2016) описали небольшую (длиной рыла и отверстия 10,6 мм), вероятно, неонатальную ящерицу, сохранившуюся в меловом ( граница альба и сеномана ) янтаре из Мьянмы . Авторы отметили, что у ящерицы «короткий и широкий череп, большие глазницы, удлиненный и мощный язычный отросток, лобная часть с параллельными краями, зарождающийся префронтальный выступ, уменьшенные сошники, отсутствие ретроартикулярного отростка, низкое количество пресакральных позвонков (от 15 до 17) и чрезвычайно короткий, завитый хвост»; авторы считали эти признаки показателем принадлежности ящерицы к Chamaeleonidae. Проведенный авторами филогенетический анализ показал, что ящерица была стволовым -хамелеонидом. [34] Однако Мацумото и Эванс (2018) интерпретировали этот экземпляр как амфибию -альбанерпетонтид . [35] В 2020 году этому экземпляру было присвоено имя Якша перетти , и было отмечено, что он имеет несколько конвергентно сходных с хамелеоном особенностей, включая приспособления для баллистического питания. [36]

Хотя точная эволюционная история изменения цвета хамелеонов до сих пор неизвестна, есть один аспект эволюционной истории изменения цвета хамелеонов, который уже окончательно изучен: эффекты эффективности сигналов. Было показано, что эффективность сигнала, или насколько хорошо сигнал виден на его фоне, напрямую коррелирует со спектральными качествами дисплеев-хамелеонов. [37] Карликовые хамелеоны, изучаемые хамелеоны, обитают в самых разных средах обитания: от лесов до лугов и кустарников. Было продемонстрировано, что хамелеоны в более ярких областях, как правило, подают более яркие сигналы, но хамелеоны в более темных областях, как правило, подают сигналы, относительно более контрастные по отношению к своему фону. Это открытие предполагает, что эффективность сигнала (и, следовательно, среда обитания) повлияла на эволюцию передачи сигналов хамелеона. Стюарт-Фокс и др. обратите внимание, что имеет смысл считать, что отбор по крипсису не так важен, как отбор по эффективности сигнала, поскольку сигналы показаны лишь кратко; хамелеоны почти всегда имеют приглушенные загадочные цвета. [37]
Описание

Хамелеоны сильно различаются по размеру и строению тела: максимальная общая длина варьируется от 22 мм (0,87 дюйма) у самца Brookesia nana в мире (одна из самых маленьких рептилий ) до 68,5 см (27,0 дюйма) у самца Furcifer oustaleti . [38] [39] У многих есть украшения на голове или лице, такие как носовые выступы или роговидные выступы в случае Trioceros jacksonii , или большие гребни на макушке, как у Chamaeleo calyptratus . Многие виды имеют половой диморфизм , и самцы обычно гораздо более украшены, чем самки хамелеонов.
Типичные размеры видов хамелеонов, которые обычно содержатся в неволе или в качестве домашних животных:
| Научное название | Общее имя | Длина (мужской) | Длина (женский) | Цвет | Продолжительность жизни (лет) |
|---|---|---|---|---|---|
| Хамелео калиптрат | Завуалированный хамелеон | 35–60 см | 25–33 см | Зеленый и светлые цвета | около 5 |
| Триоцерос Джексонии | Хамелеон Джексона | 23–33 см | 25–33 см | Зеленый и светлые цвета | 5–10 |
| Острый край пантеры | Пантера-хамелеон | 38–53 см | 23–33 см | Более темные цвета | около 5 (2–3 для рожавших самок) |
| Риппелеон бревикаудатус | Бородатый карликовый хамелеон | 5–8 см | 5–8 см | Коричневый, бежевый, зеленый | около 3–5 |
| Спектр рамполеона | Спектральный карликовый хамелеон | 8–10 см | 5–10 см | Тан и серый | 3–5 |
| Рамполеон временной | Усамбарский карликовый хамелеон без косточек | 6–10 см | 5–9 см | Серый и коричневый | 5–11 |
Ноги хамелеонов хорошо приспособлены к древесному передвижению , а такие виды, как Chamaeleo namaquensis , которые вторично переняли наземный образ жизни, сохранили ту же морфологию стоп с небольшими изменениями. На каждой стопе пять отдельных пальцев сгруппированы в два пучка. Пальцы в каждом пучке связаны в уплощенную группу по два или по три, что придает каждой ступне вид щипцов . На передних лапах наружная, латеральная группа содержит два пальца, а внутренняя, медиальная , группа — три. На задних лапах расположение обратное: медиальная группа содержит два пальца, а латеральная группа — три. Эти специализированные ноги позволяют хамелеонам крепко цепляться за узкие или грубые ветки. Кроме того, каждый палец снабжен острым когтем, позволяющим захватывать такие поверхности, как кора, при лазании. Ступни хамелеонов принято называть дидактильными или зигодактильными , хотя ни один из этих терминов не является полностью удовлетворительным, поскольку оба используются для описания разных стоп, таких как зигодактильные лапы попугаев или дидактильные лапы ленивцев или страусов, ни одна из которых не имеет существенного значения. как ноги хамелеона. Хотя термин «зигодактиль» достаточно хорошо описывает анатомию стопы хамелеона, структура его стопы не похожа на структуру стопы попугаев, к которым этот термин был впервые применен. Что касается дидактилии, у хамелеонов на каждой ноге явно по пять пальцев, а не по два.
У некоторых хамелеонов имеется гребень из небольших шипов, идущий вдоль позвоночника от проксимальной части хвоста до шеи; как протяженность, так и размер шипов различаются у разных видов и особей. Эти шипы помогают разбить четкий контур хамелеона, что помогает ему слиться с фоном.
Чувства
У хамелеонов самые отличительные глаза среди всех рептилий. Верхнее и нижнее веко соединены, имеется только маленькое отверстие, достаточно большое, чтобы зрачок мог видеть. Каждый глаз может поворачиваться и фокусироваться независимо, что позволяет хамелеону наблюдать два разных объекта одновременно. Это дает им полный обзор на 360 градусов вокруг своего тела. Местонахождение добычи осуществляется с использованием монокулярного восприятия глубины , а не стереопсиса . [40] Хамелеоны имеют самое большое увеличение (на размер) среди всех позвоночных. [41] с наибольшей плотностью колбочек в сетчатке . [42]
Как и у змей , у хамелеонов нет ни наружного, ни среднего уха , поэтому нет ни ушного отверстия, ни барабанной перепонки. Однако хамелеоны не глухие: они могут улавливать звуковые частоты в диапазоне 200–600 Гц. [43]
Хамелеоны могут видеть как в видимом, так и в ультрафиолетовом свете . [44] Хамелеоны, подвергшиеся воздействию ультрафиолета, демонстрируют повышенное социальное поведение и уровень активности, более склонны греться, питаться и размножаться, поскольку это положительно влияет на шишковидную железу .
Кормление
Все хамелеоны в основном насекомоядные , которые питаются баллистически высовывая изо рта свои длинные языки , чтобы поймать добычу, находящуюся на некотором расстоянии. [45] Обычно считается, что языки хамелеонов в полтора-два раза длиннее их тела (их длина, исключая хвост), недавно было обнаружено, что у более мелких хамелеонов (как более мелких видов, так и более мелких особей того же вида) Языковые аппараты пропорционально большего размера, чем их более крупные аналоги. [46] Таким образом, более мелкие хамелеоны способны выдвигать язык на большее расстояние, чем более крупные хамелеоны, которые являются предметом большинства исследований и оценок длины языка, и могут выдвигать язык более чем в два раза больше длины тела. [47]
Аппарат языка состоит из сильно модифицированных подъязычных костей , мышц языка и коллагеновых элементов . [48] [49] [46] [50] Подъязычная кость имеет удлиненный параллельный выступ, называемый энтоглоссальным отростком, над которым расположена трубчатая мышца – ускорительная мышца. [46] [50] [48] [49] Мышца-ускоритель сокращается вокруг челюстно-язычного отростка и отвечает за создание работы по усилению проекции языка как непосредственно, так и за счет нагрузки коллагеновых элементов, расположенных между челюстно-язычным отростком и мышцей-ускорителем. [45] [46] [48] [49] Мышца, втягивающая язык, подъязычная мышца, соединяет подъязычную и ускоряющую мышцы и отвечает за втягивание языка обратно в рот после его выбрасывания. [45] [46] [50] [48]
Выбрасывание языка происходит с чрезвычайно высокой скоростью, достигая добычи всего за 0,07 секунды. [48] [49] [51] был запущен с ускорением, превышающим 41 g . [51] Мощность , с которой запускается язычок, как известно, превышает 3000 Вт кг. −1 , превышает то, которое способны производить мышцы, что указывает на наличие эластичного усилителя мощности для усиления выступания языка. [49] Таким образом, отдача упругих элементов в аппарате языка ответственна за большую часть общей эффективности выдвижения языка.
Одним из последствий включения механизма упругой отдачи в механизм выдвижения языка является относительная термическая нечувствительность выдвижения языка по отношению к втягиванию языка, которое приводится в действие только за счет сокращения мышц и очень чувствительно к температуре. [51] [52] В то время как другие экзотермные животные становятся вялыми по мере снижения температуры их тела из-за снижения скорости сокращения их мышц, хамелеоны способны с высокой производительностью высовывать язык даже при низких температурах тела. [51] [52] Однако термическая чувствительность втягивания языка у хамелеонов не является проблемой, поскольку хамелеоны обладают очень эффективным механизмом удержания своей добычи после того, как язык вошел с ней в контакт, включая поверхностные явления, такие как мокрое прилипание и блокирование, а также всасывание. [53] Таким образом, термическая нечувствительность выступа языка позволяет хамелеонам эффективно питаться холодным утром, прежде чем они смогут поведенчески повышать температуру своего тела посредством терморегуляции , когда другие симпатрические виды ящериц все еще неактивны, что, вероятно, временно расширяет их тепловую нишу . в результате [51]
- Использование языка при кормлении
 Строение языка с чашеобразным концом.
Строение языка с чашеобразным концом. Язык начинает забастовку
Язык начинает забастовку Захват добычи
Захват добычи Поднесение добычи ко рту
Поднесение добычи ко рту




Кости
У некоторых видов хамелеонов есть кости, которые светятся в ультрафиолетовом свете , также известном как биогенная флуоресценция . [25] Около 31 различных видов хамелеонов Calumma , обитающих на Мадагаскаре , продемонстрировали эту флуоресценцию при компьютерной томографии . [54] Кости излучали ярко-голубое свечение и могли светиться даже сквозь четыре слоя кожи хамелеона. [54] Было обнаружено, что лицо имело другое сияние, проявляющееся в виде точек, также известных как бугорки на костях лица. [25] Свечение возникает благодаря белкам , пигментам , хитину и другим материалам, из которых состоит скелет хамелеона. [25] возможно, хамелеоны получили вторичную сигнальную систему, которая не влияет на их способность менять цвет и, возможно, возникла в результате полового отбора . [25]
Распространение и среда обитания

Хамелеоны в основном обитают на материковой части Африки к югу от Сахары и на острове Мадагаскар, хотя несколько видов обитают в Северной Африке , Южной Европе (Португалия, Испания, Италия, Греция, Кипр и Мальта), на Ближнем Востоке , на юго-востоке Пакистана , Индия , Шри-Ланка и несколько небольших островов в западной части Индийского океана . Завезенные , неместные популяции встречаются на Гавайях и во Флориде. [1]
Хамелеоны встречаются только в тропических и субтропических регионах и населяют все виды равнинных и горных лесов, лесных массивов, кустарников, саванн , а иногда и пустынь , но каждый вид, как правило, ограничен только одним из нескольких различных типов среды обитания. Типичные хамелеоны из подсемейства Chamaeleoninae ведут древесный образ жизни , обычно живут на деревьях или кустах, хотя некоторые (особенно хамелеон Намаква ) частично или в значительной степени ведут наземный образ жизни . Род Brookesia , который включает большинство видов подсемейства Brookesiinae, обитает низко в растительности или на земле среди опавших листьев . Многие виды хамелеонов имеют небольшое распространение и считаются находящимися под угрозой исчезновения. Сокращение численности хамелеонов в основном связано с утратой среды обитания. [55]
Воспроизведение
Большинство хамелеонов яйцекладущие , но все виды Bradypodion и многие Trioceros виды являются яйцеживородящими (хотя некоторые биологи предпочитают избегать термина яйцеживородящие из-за несоответствия с его использованием в некоторых группах животных, вместо этого просто используя живородящие ). [56]
Яйцекладущие виды откладывают яйца через три-шесть недель после совокупления . Самка выкапывает яму глубиной 10–30 см (4–12 дюймов), в зависимости от вида, и откладывает яйца. Размеры кладки сильно различаются в зависимости от вида. Мелкие виды Brookesia могут откладывать только два-четыре яйца, в то время как крупные покрытые хамелеоны ( Chamaeleo calyptratus ), как известно, откладывают кладки из 20–200 (покрытые хамелеоны) и 10–40 (пантеровые хамелеоны) яиц. Размеры кладки также могут сильно различаться у одного и того же вида. Яйца обычно вылупляются через четыре-12 месяцев, опять же в зависимости от вида. Яйца хамелеона Парсона ( Calumma parsoni ) обычно вылупляются через 400–660 дней. [57]
Хамелеоны откладывают яйца с гибкой скорлупой, на которые во время инкубации влияют характеристики окружающей среды. Масса яйца является наиболее важной для дифференциации выживших хамелеонов во время инкубации. Увеличение яичной массы будет зависеть от температуры и водного потенциала. [58] Чтобы понять динамику водного потенциала в яйцах хамелеонов, необходимо учитывать давление, оказываемое на яичную скорлупу, поскольку давление яичной скорлупы играет важную роль в водном отношении яиц в течение всего инкубационного периода. [59]
Яйцеживородящие виды, такие как хамелеон Джексона ( Trioceros jacksonii ), имеют период беременности от пяти до семи месяцев. Каждый молодой хамелеон рождается внутри липкой прозрачной оболочки желточного мешка. Мать прижимает каждое яйцо к ветке, где оно и прилипает. Мембрана разрывается, и только что вылупившийся хамелеон освобождается и уползает, чтобы охотиться и прятаться от хищников. За одну беременность у самки может родиться до 30 живых детенышей. [60]
Диета
Хамелеоны обычно едят насекомых , но более крупные виды, такие как обыкновенный хамелеон, могут также питаться другими ящерицами и молодыми птицами . [61] : 5 Разнообразие диет можно увидеть на следующих примерах:
- Скрытый хамелеон Chamaeleo calyptratus из Аравии насекомоядный, но питается листьями, когда другие источники воды недоступны. Его можно поддерживать на диете из сверчков . [62] Они могут съесть до 15–50 крупных сверчков в день.
- Хамелеон Джексона ( Trioceros jacksonii ) из Кении и северной Танзании поедает самых разных мелких животных, включая муравьев, бабочек, гусениц, улиток, червей, ящериц, гекконов, амфибий и других хамелеонов, а также растительный материал, такой как листья, нежные побеги и ягоды. Его можно поддерживать на смешанной диете, включающей капусту, листья одуванчика, салат, бананы, помидоры, яблоки, сверчков и свиристелей. [60]
- Обыкновенный хамелеон Европы, Северной Африки и Ближнего Востока Chamaeleo chamaeleon питается преимущественно осами и богомолами ; такие членистоногие составляют более трех четвертей его рациона. [61] : 5 Некоторые эксперты советуют не кормить обыкновенного хамелеона исключительно сверчками; они должны составлять не более половины рациона, а остальная часть должна составлять смесь восковых червей , дождевых червей , кузнечиков , мух и растительных материалов, таких как зеленые листья, овес и фрукты. [61] : 5–6
- Некоторые хамелеоны, такие как хамелеон, пантеровый мадагаскарский регулируют уровень витамина D3 , плохим источником которого является их рацион насекомых, подвергая себя воздействию солнечного света , поскольку его УФ- компонент увеличивает внутреннее производство. [63]
Адаптации против хищников
На хамелеонов охотятся множество других животных. Птицы и змеи — важнейшие хищники взрослых хамелеонов. Беспозвоночные, особенно муравьи, оказывают сильное хищническое давление на яйца и молодь хамелеонов. [64] Хамелеоны вряд ли смогут убежать от хищников и полагаются на крипсис как на свою основную защиту. [65] Хамелеоны могут менять как свой цвет, так и узор (в разной степени), чтобы походить на свое окружение, или нарушать очертания тела и оставаться скрытыми от взгляда потенциального врага. Только в случае обнаружения хамелеоны активно защищаются. Они принимают защитную позу, представляют нападающему сплюснутое с боков тело, чтобы казаться крупнее, предупреждают открытым ртом и, при необходимости, используют ноги и челюсти для отпора. [66] Вокализация иногда включается в демонстрацию угроз. [64]
- (видео) Хамелеон в Малави
 Хамелеон найден в Майсуре , Южная Индия.
Хамелеон найден в Майсуре , Южная Индия. Этот обыкновенный хамелеон ( Chamaeleo chamaeleon ) почернел.
Этот обыкновенный хамелеон ( Chamaeleo chamaeleon ) почернел. Хамелеон с откидной шеей Chamaeleo dilepis , на которого напал бумсланг , когда он переходил дорогу в Намибии, принял угрожающую защитную позу.
Хамелеон с откидной шеей Chamaeleo dilepis , на которого напал бумсланг , когда он переходил дорогу в Намибии, принял угрожающую защитную позу. Хамелеон Намаква , демонстрирующий угрозу , в национальном парке Намиб-Науклуфт , почернел и открыл пасть, когда была предпринята попытка сдвинуть его с оживленной дороги.
Хамелеон Намаква , демонстрирующий угрозу , в национальном парке Намиб-Науклуфт , почернел и открыл пасть, когда была предпринята попытка сдвинуть его с оживленной дороги.




Паразиты
хамелеонах паразитируют На нематоды , в том числе острицы ( Filarioidea ). Острицы могут передаваться через укусы насекомых, таких как клещи и комары . Другие круглые черви передаются через пищу, зараженную яйцами круглых червей; личинки проникают через стенку кишечника в кровоток. [67]
Хамелеоны подвержены воздействию нескольких простейших паразитов, таких как Plasmodium , вызывающий малярию , Trypanosoma , вызывающий сонную болезнь , и Leishmania , вызывающий лейшманиоз . [68]
Хамелеоны подвержены паразитизму кокцидий , [68] включая виды родов Choleoeimeria , Eimeria и Isospora . [69]
Как домашние животные
Хамелеоны — популярные домашние животные-рептилии, в основном импортированные из африканских стран, таких как Мадагаскар, Танзания и Того. [70] Наиболее распространены в промысле сенегальский хамелеон ( Chamaeleo senegalensis ), йеменский или вуалированный хамелеон ( Chamaeleo calyptratus ), пантеровый хамелеон ( Furcifer pardalis ) и хамелеон Джексона ( Trioceros jacksonii ). [70] Другие хамелеоны, встречающиеся в неволе (хотя и нерегулярно), включают такие виды, как ковровый хамелеон ( Furcifer Lateralis ), хамелеон Меллера ( Trioceros melleri ), хамелеон Парсона ( Calumma parsonii ), а также несколько видов карликовых и листовидных хамелеонов, в основном из родов Brookesia , Rampholeon или Rieppeleon . Это одни из самых чувствительных рептилий, которые только можно иметь, требующие особого внимания и заботы.
США являются основным импортером хамелеонов с начала 1980-х годов, на их долю приходится 69% экспорта африканских рептилий. [70] Однако произошло значительное снижение численности из-за ужесточения правил защиты видов от вывоза из дикой природы, а также из-за того, что многие из них стали инвазивными в таких местах, как Флорида. [70] Однако они остались популярными, что может быть связано с разведением в неволе в США, которое возросло до такой степени, что США могут удовлетворить свои потребности, а теперь даже стали крупным экспортером. [70] В США они настолько популярны, что, несмотря на то, что во Флориде из-за торговли домашними животными обитает шесть инвазивных видов хамелеонов, любители рептилий в этих районах ищут хамелеонов, чтобы держать их в качестве домашних животных или разводить и продавать, причем некоторые из них продаются по цене до тысячи долларов. . [3]
Исторические понимания

Аристотель (4 век до н. э.) описывает хамелеонов в своей «Истории животных» . [71] Плиний Старший (I век нашей эры) также обсуждает хамелеонов в своей «Естественной истории» . [72] отмечая их способность менять цвет для маскировки.
Хамелеон был показан в Конрада Гесснера «Истории животных» (1563 г.), скопированной с «De aquatilibus» (1553 г.) Пьера Белона . [73]
В шекспировском «Гамлете » одноименный принц говорит: «Я верю, превосходное блюдо хамелеона. Я ем воздух, набитый обещаниями». Это относится к елизаветинской вере в то, что хамелеоны живут только в воздухе.
Ссылки
- ^ Jump up to: а б с д Гло, Ф. (2015). «Таксономический контрольный список хамелеонов (Squamata: Chamaeleonidae)» . Зоология позвоночных . 65 (2): 167–246. дои : 10.3897/vz.65.e31518 .
- ^ Эдмондс, Патрисия (сентябрь 2015 г.). «Настоящие цвета». Нэшнл Географик : 98.
- ^ Jump up to: а б Дейли, Наташа (2017). «Внутри секретного мира ловцов хамелеонов Флориды» . Нэшнл Географик . Архивировано из оригинала 9 ноября 2020 года.
- ^ хамелеон . Чарльтон Т. Льюис и Чарльз Шорт. Латинский словарь по проекту «Персей» .
- ^ χαμαιλέων . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей» .
- ^ χαμαί у Лидделла и Скотта .
- ^ лев у Лидделла и Скотта .
- ^ «Хамелеон» . Словарь.com .
- ^ Харпер, Дуглас. «хамелеон» . Интернет-словарь этимологии .
- ^ Клавер, К.; Бёме, В. (1986). «Филогения и классификация Chamaeleonidae (Sauria) с особым упором на морфологию гемипениса» . Боннер Зоологическая монография . 22 : 1–64.
- ^ Jump up to: а б Тилбери, Колин (2010). Хамелеоны Африки, Атлас, включающий хамелеонов Европы, Ближнего Востока и Азии . Франкфурт: Издание Chimaira. ISBN 978-3899734515 .
- ^ Таунсенд, Т.; Ларсон, А. (2002). «Молекулярная филогенетика и митохондриальная геномная эволюция Chamaeleonidae (Reptilia, Squamata)». Молекулярная филогенетика и эволюция . 23 (1): 22–36. дои : 10.1006/mpev.2001.1076 . ПМИД 12182400 .
- ^ Раксуорси, CJ; Форстнер, MRJ; Нуссбаум, РА (2002). «Излучение хамелеона в результате распространения в океане» (PDF) . Природа . 415 (6873): 784–787. Бибкод : 2002Natur.415..784R . дои : 10.1038/415784a . hdl : 2027.42/62614 . ПМИД 11845207 . S2CID 4422153 .
- ^ Таунсенд, ТМ; Толли, Калифорния; Гло, Ф.; и др. (2011). «На восток от Африки: распространение хамелеонов на Сейшельские острова, опосредованное палеотечением» . Биологические письма . 7 (2): 225–228. дои : 10.1098/rsbl.2010.0701 . ПМК 3061160 . ПМИД 20826471 .
- ^ Jump up to: а б Толли, Калифорния; Таунсенд, ТМ; Венсес, М. (2013). «Крупномасштабная филогения хамелеонов предполагает африканское происхождение и эоценовую диверсификацию» . Труды Королевского общества Б. 280 (1759): 20130184. doi : 10.1098/rspb.2013.0184 . ПМЦ 3619509 . ПМИД 23536596 .
- ^ Тилбери, Колин (2014). «Обзор систематики Chamaeleonidae». В Толли, Кристал А.; Херрел, Энтони (ред.). Биология хамелеонов . Беркли: Издательство Калифорнийского университета. стр. 151–174. ISBN 9780520276055 .
- ^ Шэрон Кац Купер. «Хамелеоны» . Национальный географический исследователь. Архивировано из оригинала 20 августа 2008 года.
- ^ Jump up to: а б с Тейсье, Жереми; Саенко, Сюзанна В.; ван дер Марель, Дирк; Милинкович, Мишель К. (10 марта 2015 г.). «Фотонные кристаллы вызывают активное изменение цвета у хамелеонов» . Природные коммуникации . 6 (1): 1–7. Бибкод : 2015NatCo...6.6368T . дои : 10.1038/ncomms7368 . ISSN 2041-1723 . ПМЦ 4366488 . ПМИД 25757068 .
- ^ Стюарт-Фокс, Д.; Муссалли, А. (2008). «Отбор социальных сигналов стимулирует эволюцию изменения цвета хамелеона» . ПЛОС Биология . 6 (1): е25. doi : 10.1371/journal.pbio.0060025 . ПМК 2214820 . ПМИД 18232740 .
- ^ Харрис, Том (18 мая 2001 г.). «Как работает камуфляж для животных» . Как все работает . Проверено 13 ноября 2006 г.
- ^ Уолтон, Б. Майкл; Беннетт, Альберт Ф. (1993). «Изменение цвета кенийских хамелеонов в зависимости от температуры». Физиологическая зоология . 66 (2). Издательство Чикагского университета: 270–287. дои : 10.1086/physzool.66.2.30163690 . ISSN 0031-935X . S2CID 80673490 .
- ^ Кук, Мария. «Адаптации хамелеонов» . Наука . Проверено 15 июня 2020 г.
- ^ Лигон, Рассел А.; МакГроу, Кевин Дж. (2013). «Во время соревнований хамелеоны общаются с помощью сложных изменений цвета: разные участки тела передают разную информацию» . Письма по биологии . 9 (6): 20130892. doi : 10.1098/rsbl.2013.0892 . ПМЦ 3871380 . ПМИД 24335271 .
- ^ Лигон, Рассел А. (2014). «Побежденные хамелеоны динамически темнеют во время диадических споров, чтобы уменьшить опасность со стороны доминантов». Поведенческая экология и социобиология . 68 (6): 1007–1017. дои : 10.1007/s00265-014-1713-z . S2CID 18606633 .
- ^ Jump up to: а б с д и Претцель, Дэвид; Хесс, Мартин; Шерц, Марк Д.; и др. (15 января 2018 г.). «Распространенная флуоресценция костей у хамелеонов» . Научные отчеты . 8 (1): 698. Бибкод : 2018НатСР...8..698П . дои : 10.1038/s41598-017-19070-7 . ISSN 2045-2322 . ПМК 5768862 . ПМИД 29335580 .
- ^ Янг, Эмма (2008) Хамелеоны настраивают камуфляж под зрение хищника . Новый учёный
- ^ Стюарт-Фокс, Д.; Муссалли, А. (2009). «Камуфляж, коммуникация и терморегуляция: уроки организмов, меняющих цвет» . Philos Trans R Soc Lond B Biol Sci . 364 (1516): 463–470. дои : 10.1098/rstb.2008.0254 . ПМК 2674084 . ПМИД 19000973 .
- ^ Уайтинг, MJ; Голландия, бакалавр наук; Кио, Дж.С.; Ноубл, DWA; Рэнкин, К.Дж.; Стюарт-Фокс, Д. (2022). «Агрессивные хамелеоны, выпущенные от хищников, демонстрируют более заметные цвета» . Достижения науки . 8 (19): eabn2415. Бибкод : 2022SciA....8N2415W . дои : 10.1126/sciadv.abn2415 . ПМК 9094656 . ПМИД 35544573 .
- ^ Патрисия Эдмондс (2015). «Красочный язык хамелеонов» . Национальное географическое общество. Архивировано из оригинала 11 марта 2016 года.
- ^ Стюарт-Фокс, Деви; Муссалли, Аднан (29 января 2008 г.). «Отбор социальных сигналов стимулирует эволюцию изменения цвета хамелеона» . ПЛОС Биология . 6 (1): е25. doi : 10.1371/journal.pbio.0060025 . ISSN 1545-7885 . ПМК 2214820 . ПМИД 18232740 .
- ^ Jump up to: а б Майзано, Джесси (27 августа 2003 г.). «Chamaeleo calyptratus, Скрытый Хамелеон» . Дигиморф . Техасский университет в Остине . Проверено 10 января 2012 г.
- ^ Толли, Кристал; Бургер, Мариус (2007). Хамелеоны Южной Африки . Струйк. стр. 26–28. ISBN 978-1-77007-375-3 .
- ^ Болет А., Эванс С.Э. (16 ноября 2013 г.). «Ископаемая история хамелеонов» . В Толли К.А., Херрел А. (ред.). Биология хамелеонов . Университет Калифорнии Пресс. ISBN 9780520276055 . Проверено 1 ноября 2017 г. - через Google Книги.
- ^ Даса, Хуан Д.; Стэнли, Эдвард Л.; Вагнер, Филипп; и др. (2016). «Окаменелости янтаря среднего мела проливают свет на прошлое разнообразие тропических ящериц» . Достижения науки . 2 (3): e1501080. Бибкод : 2016SciA....2E1080D . дои : 10.1126/sciadv.1501080 . ПМЦ 4783129 . ПМИД 26973870 .
- ^ Рёко Мацумото; Сьюзан Э. Эванс (2018). «Первая запись амфибий-альбанерпетонтид (Amphibia: Albanerpetontidae) из Восточной Азии» . ПЛОС ОДИН . 13 (1): e0189767. Бибкод : 2018PLoSO..1389767M . дои : 10.1371/journal.pone.0189767 . ПМК 5752013 . ПМИД 29298317 .
- ^ Даса, Хуан Д.; Стэнли, Эдвард Л.; Болет, Арнау; и др. (06.11.2020). «Загадочные земноводные в янтаре середины мелового периода были баллистическими кормушками, похожими на хамелеонов» . Наука . 370 (6517): 687–691. Бибкод : 2020Sci...370..687D . дои : 10.1126/science.abb6005 . ISSN 0036-8075 . ПМИД 33154135 . S2CID 226254862 .
- ^ Jump up to: а б Стюарт-Фокс, Д.; Муссалли, Аднан; Уайтинг, Мартин Дж. (2007). «Естественный отбор социальных сигналов: эффективность сигналов и эволюция окраски дисплея хамелеона» . Американский натуралист . 170 (6): 916–930. дои : 10.1086/522835 . ПМИД 18171173 . S2CID 21716855 .
- ^ Гло, Фрэнк; Кёлер, Йорн; Хавличек, Оливер; Рацоавина, Фаномезана М.; Ракотоарисон, Андолалао; Шерц, Марк Д. и Венсес, Мигель (28 января 2021 г.). «Чрезвычайная миниатюризация нового амниотного позвоночного и понимание эволюции размера гениталий хамелеонов» . Научные отчеты . 11 (1): 2522. doi : 10.1038/s41598-020-80955-1 . ПМЦ 7844282 . ПМИД 33510189 .
- ^ Гло, Фрэнк; Венсес, Мигель (1994). Полевой справочник по амфибиям и рептилиям Мадагаскара (2-е изд.). Кёльн: Verlags GbR. п. 253. ИСБН 978-3-929449-01-3 .
- ^ Отт, М.; Шеффель, Ф.; Кирмсе, В. (1998). «Бинокулярное зрение и аккомодация хамелеонов, ловящих добычу». Журнал сравнительной физиологии А. 182 (3): 319–330. дои : 10.1007/s003590050182 . S2CID 19988312 .
- ^ Отт, Матиас; Шеффель, Франк (1995). «Линза с отрицательной оптической силой в хамелеоне». Природа . 373 (6516): 692–694. Бибкод : 1995Natur.373..692O . дои : 10.1038/373692a0 . ПМИД 7854450 . S2CID 4262985 .
- ^ Стюарт-Фокс, Деви (2014). «Поведение хамелеона и изменение цвета». В Толли, Кристал А.; Херрел, Энтони (ред.). Биология хамелеонов . Беркли: Издательство Калифорнийского университета. стр. 115–130. ISBN 9780520276055 .
- ^ Ле Берр и Бартлетт, стр. 31.
- ^ «Новости Хамелеона» . Хамелеонньюс.com . Август 2004 г. Архивировано из оригинала 22 января 2008 г. Проверено 1 ноября 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Jump up to: а б с Хайэм, штат Техас; Андерсон, CV (2014), «Функции и адаптация хамелеонов», Толли, Калифорния; Херрел, А. (ред.), Биология хамелеонов , Беркли, Калифорния: University of California Press, стр. 63–83, ISBN. 9780520276055
- ^ Jump up to: а б с д и Андерсон, резюме; Шеридан, Т.; Дебан, С.М. (2012). «Масштабирование баллистического языкового аппарата у хамелеонов». Журнал морфологии . 273 (11): 1214–1226. дои : 10.1002/Jmor.20053 . ПМИД 22730103 . S2CID 21033176 .
- ^ Андерсон, Кристофер В. (2009) Rhamholeon spinosus Видео о кормлении . chamaeleonidae.com
- ^ Jump up to: а б с д и Херрел, А.; Мейерс, Джей-Джей; Нисикава, КЦ; Де Ври, Ф. (2001). «Морфология и гистохимия гиолингвального аппарата хамелеонов». Журнал морфологии . 249 (2): 154–170. дои : 10.1002/jmor.1047 . ПМИД 11466743 . S2CID 3246256 .
- ^ Jump up to: а б с д и де Гроот, Дж. Х.; ван Леувен, JL (2004). «Доказательства эластичного механизма проекции языка хамелеона» . Труды Лондонского королевского общества Б. 271 (1540): 761–770. дои : 10.1098/rspb.2003.2637 . ПМК 1691657 . ПМИД 15209111 .
- ^ Jump up to: а б с Андерсон, резюме; Хайэм, Т.Э. (2014), «Анатомия хамелеона», Толли, Калифорния; Херрел, А. (ред.), Биология хамелеонов , Беркли, Калифорния: University of California Press, стр. 7–55, ISBN. 9780520276055
- ^ Jump up to: а б с д и Андерсон, резюме; Дебан, С.М. (2010). «Баллистическая проекция языка у хамелеонов сохраняет высокую эффективность при низкой температуре» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (12): 5495–5499. Бибкод : 2010PNAS..107.5495A . дои : 10.1073/pnas.0910778107 . ПМЦ 2851764 . ПМИД 20212130 .
- ^ Jump up to: а б Андерсон, резюме; Дебан, С.М. (2012). «Термическое воздействие на двигательный контроль и in vitro динамику мышц баллистического языка у хамелеонов» . Журнал экспериментальной биологии . 215 (24): 4345–4357. дои : 10.1242/jeb.078881 . ПМИД 23125336 .
- ^ Херрел, А.; Мейерс, Джей-Джей; Аэртс, П.; Нисикава, К.К. (2000). «Механика захвата добычи у хамелеонов» (PDF) . Журнал экспериментальной биологии . 203 (Часть 21): 3255–3263. дои : 10.1242/jeb.203.21.3255 . ПМИД 11023845 . Архивировано из оригинала (PDF) 20 июня 2010 г. Проверено 16 ноября 2014 г.
- ^ Jump up to: а б Элейна Захос (18 января 2018 г.). «Кости хамелеона светятся в темноте даже сквозь кожу» . Нэшнл Географик . Архивировано из оригинала 18 января 2018 года . Проверено 3 августа 2018 г.
- ^ «Утрата и фрагментация среды обитания сокращают популяцию хамелеонов в Танзании» . Физика.орг . Проверено 1 ноября 2017 г.
- ^ Хьюз, Д.Ф.; Блэкберн, генеральный директор (2020). «Эволюционное происхождение живорождения у Chamaeleonidae» . Журнал зоологической систематики и эволюционных исследований . 58 (1): 284–302. дои : 10.1111/jzs.12328 .
- ^ Лаубе, Александра; Негр, Торстен; Огюстен, Андреас (2020). «781 день в яйце: длительное время инкубации у Calumma parsonii parsonii (Cuvier, 1824), что привело к появлению здоровой молоди и выявило косвенные доказательства задержки спермы у этого вида» . Герпетологические заметки . 13 : 425–428.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Диас-Паниагуа С, Куадрадо М (2003), «Влияние условий инкубации на успех вылупления, развитие эмбрионов и фенотип вылупления яиц обыкновенного хамелеона ( Chamaeleo chamaeleon )», Amphibia-Reptilia , 24 (4): 429–440, doi : 10.1163/156853803322763891
- ^ Эндрюс (2008), «Влияние температуры инкубации на рост и продуктивность скрытого хамелеона (Chamaeleo calyptratus», Журнал экспериментальной зоологии. Часть A, Экологическая генетика и физиология , 309 (8), Журнал экспериментальной зооли: 435–446, Bibcode : 2008JEZA..309..435A , doi : 10.1002/jez.470 , PMID 18512704
- ^ Jump up to: а б «Африканский тропический лес» . Хамелеон Джексона . Зоопарк Торонто. Архивировано из оригинала 11 ноября 2011 года . Проверено 9 января 2012 г.
- ^ Jump up to: а б с Девер, Дженнифер (5 декабря 2007 г.). «Обыкновенный Хамелеон» (PDF) . usfca.edu. Архивировано из оригинала (PDF) 3 февраля 2015 года . Проверено 9 января 2012 г.
- ^ «Рептилии и амфибии: завуалированный хамелеон» . Смитсоновский национальный зоологический парк. Архивировано из оригинала 17 декабря 2011 г. Проверено 9 января 2012 г.
- ^ Карстен, КБ; Фергюсон Г.В.; Чен ТЦ; Холик М.Ф. (2009). «Пантеровые хамелеоны, Furcifer pardalis, поведенчески регулируют оптимальное воздействие ультрафиолета в зависимости от уровня витамина D3 в рационе». Физиол. Биохим. Зоол . 82 (3): 218–25. дои : 10.1086/597525 . ПМИД 19335229 . S2CID 205990383 .
- ^ Jump up to: а б Стюарт-Фокс Д. (2014). «Поведение хамелеона и изменение цвета». В Толли К.А., Херрел А. (ред.). Биология хамелеонов . Беркли: Издательство Калифорнийского университета. стр. 115–130. ISBN 9780520276055 .
- ^ Миси Г.Дж., Раселиманана А., Херрел А. (2014). «Экология и история жизни хамелеонов». В Толли К.А., Херрел А. (ред.). Биология хамелеонов . Беркли: Издательство Калифорнийского университета. стр. 85–114. ISBN 9780520276055 .
- ^ Берг, Филипп; Берг, Джессика; Берг, Райнер (2020). «Взаимодействие хищника и жертвы между бумслангом типа Dispholidus и хамелеоном с откидной шеей Chamaeleo dilepis». Африканский журнал экологии . 58 (4): 855–859. дои : 10.1111/aje.12782 . S2CID 225209615 .
- ^ Ле Берр и Бартлетт, стр. 110.
- ^ Jump up to: а б Ле Берр и Бартлетт, стр. 109.
- ^ Слобода, Михал; Модры, Давид (2006). «Новые виды Choleoeimeria (Apicomplexa: Eimeriidae) от скрытого хамелеона Chamaeleo calyptratus (Sauria: Chamaeleonidae) с таксономической ревизией эймерийных кокцидий от хамелеонов» . Фолиа Паразитологическая . 53 (2): 91–97. дои : 10.14411/fp.2006.012 . ПМИД 16898122 .
- ^ Jump up to: а б с д и Карпентер, Ангус И.; Маркус Роуклифф, Дж.; Уоткинсон, Эндрю Р. (2004). «Динамика мировой торговли хамелеонами» . Биологическая консервация . 120 (2): 291–301. Бибкод : 2004BCons.120..291C . doi : 10.1016/j.biocon.2004.03.002 . ISSN 0006-3207 .
- ^ Аристотель, История животных, Книга II, Часть 11 [1] .
- ^ Плиний Старший, Естественная история 8.51.
- ^ Рабинович, Одед (2013). «Хамелеоны между наукой и литературой: наблюдение, письмо и ранняя Парижская академия наук в литературной сфере». История науки . 15 (1): 47. Бибкод : 2013HisSc..51...33R . дои : 10.1177/007327531305100102 . S2CID 140879009 .
Общая библиография
- Ле Берр, Франсуа; Бартлетт, Ричард Д. (2009). Справочник Хамелеона . Образовательная серия Бэррона. 3-е издание. ISBN 0764141422 .
Дальнейшее чтение
- «Ученые обнаружили, что мадагаскарского хамелеона в последний раз видели 100 лет назад» . Ассошиэйтед Пресс . 30 октября 2020 г.
- Андерсон, резюме; Дебан, С.М. (2010). «Баллистическая проекция языка у хамелеонов сохраняет высокую эффективность при низкой температуре» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (12): 5495–5499. Бибкод : 2010PNAS..107.5495A . дои : 10.1073/pnas.0910778107 . ПМЦ 2851764 . ПМИД 20212130 .
- Андерсон, резюме; Дебан, С.М. (2012). «Термическое воздействие на двигательный контроль и in vitro динамику мышц баллистического языка у хамелеонов» . Журнал экспериментальной биологии . 215 (24): 4345–4357. дои : 10.1242/jeb.078881 . ПМИД 23125336 .
- Андерсон, резюме; Шеридан, Т.; Дебан, С.М. (2012). «Масштабирование баллистического языкового аппарата у хамелеонов». Журнал морфологии . 273 (11): 1214–1226. дои : 10.1002/Jmor.20053 . ПМИД 22730103 . S2CID 21033176 .
- Дэвисон, Линда Дж. Хамелеоны: их уход и разведение. Издательство Hancock House, 1997.
- де Гроот, Дж. Х.; ван Леувен, JL (2004). «Доказательства эластичного механизма проекции языка хамелеона» . Труды Лондонского королевского общества Б. 271 (1540): 761–770. дои : 10.1098/rspb.2003.2637 . ПМК 1691657 . ПМИД 15209111 .
- де Восьоли, Филипп. Необходимый уход за хамелеонами. Передовые системы вивария, 2004.
- Херрел, А.; Мейерс, Джей-Джей; Нисикава, КЦ; Де Ври, Ф. (2001). «Морфология и гистохимия гиолингвального аппарата хамелеонов». Журнал морфологии . 249 (2): 154–170. дои : 10.1002/jmor.1047 . ПМИД 11466743 . S2CID 3246256 .
Внешние ссылки
 СМИ, связанные с Chamaeleonidae, на Викискладе?
СМИ, связанные с Chamaeleonidae, на Викискладе?  Данные, относящиеся к Chamaeleonidae , в Wikispecies
Данные, относящиеся к Chamaeleonidae , в Wikispecies
| Базы данных органов управления : Национальные |
|---|