Нейронный дарвинизм
Эта статья написана как личное размышление, личное эссе или аргументативное эссе , в котором излагаются личные чувства редактора Википедии или представлен оригинальный аргумент по определенной теме. ( Июль 2021 г. ) |

Нейронный дарвинизм — биологический, а точнее дарвинистский и селекционистский подход к пониманию глобальной функции мозга , первоначально предложенный американским биологом, исследователем и лауреатом Нобелевской премии. [1] Джеральд Морис Эдельман (1 июля 1929 г. - 17 мая 2014 г.). Книга Эдельмана 1987 года «Нейронный дарвинизм». [2] познакомил публику с теорией выбора групп нейронов (TNGS), которая является основной теорией, лежащей в основе объяснения Эдельмана глобальной функции мозга.
Из-за названия книги TNGS чаще всего называют теорией нейронного дарвинизма , хотя корни TNGS уходят корнями в книгу Эдельмана и Маунткасла 1978 года « Внимательный мозг – корковая организация и групповая избирательная теория функций высшего мозга », в которой Коллега Эдельмана, американский нейрофизиолог и анатом Вернон Б. Маунткасл (15 июля 1918 г. - 11 января 2015 г.), описывает столбчатую структуру групп коры внутри неокортекса : [3] в то время как Эдельман развивает свои аргументы в пользу селективных процессов, действующих среди вырожденных первичных репертуаров групп нейронов. [4] На развитие нейронного дарвинизма глубокое влияние оказали работы Эдельмана в области иммунологии , эмбриологии и нейробиологии , а также его методологическая приверженность идее отбора как объединяющей основы биологических наук .
Введение в нейронный дарвинизм
[ редактировать ]Нейрондарвинизм на самом деле является нейронной частью натурфилософской и объяснительной структуры, которую Эдельман использует в большей части своей работы – соматических селективных систем. Нейрондарвинизм является основой для обширного набора биологических гипотез и теорий , разработанных Эдельманом и его командой, которые стремятся согласовать позвоночных и млекопитающих нейронов морфологию , факты биологии развития и эволюции , а также теорию естественного отбора. [5] в детальную модель нейронных и когнитивных функций в реальном времени , имеющую биологическую направленность. Он построен снизу вверх с использованием вариаций, встречающихся в природе . Это контрастирует с вычислительными и алгоритмическими подходами, которые рассматривают вариации как шум в системе логических схем с двухточечным соединением.
Книга « Нейронный дарвинизм – Теория отбора групп нейронов» (1987) является первой в трилогии книг, написанных Эдельманом, чтобы очертить масштабы и широту его идей о том, как биологическую теорию сознания и эволюции плана тела животных. можно разработать развивались по принципу «снизу вверх». В соответствии с принципами популяционной биологии и дарвиновской теорией естественного отбора – в отличие от нисходящих алгоритмических и вычислительных подходов, которые доминировали в зарождающейся когнитивной психологии того времени.
Два других тома — «Топобиология. Введение в молекулярную эмбриологию». [6] (1988) с их морфорегуляторной гипотезой развития строения тела животных и эволюционной диверсификации посредством дифференциальной экспрессии молекул клеточной поверхности во время развития; и «Вспомнившееся настоящее» – биологическая теория сознания [7] (1989) – новый биологический подход к пониманию роли и функции «сознания» и его связи с когнитивной и поведенческой физиологией.
Эдельман напишет еще четыре книги для широкой публики, объясняя свои идеи о том, как работает мозг и как сознание возникает в результате физической организации мозга и тела – « Яркий воздух, яркий огонь – о материи разума». [8] (1992), Вселенная сознания - как материя становится воображением [9] (2000) с Джулио Тонони, Шире, чем небо - феноменальный дар сознания [10] (2004) и «Вторая природа - наука о мозге и человеческие знания». [11] (2006).
Нейронный дарвинизм – это исследование биологической мысли и философии , а также фундаментальной науки ; Эдельман хорошо разбирается в истории науки, естествознания и медицины , а также в робототехнике , кибернетике , вычислениях и искусственном интеллекте . Излагая аргументы в пользу нейронного дарвинизма, или, точнее, TNGS, Эдельман обрисовывает набор концепций для переосмысления проблемы организации и функционирования нервной системы , одновременно требуя строго научных критериев для построения фундамента истинно дарвиновское и, следовательно, биологическое объяснение нейронных функций, восприятия , познания и глобальной функции мозга, способное поддерживать первичное сознание и сознание более высокого порядка.
Популяционное мышление – соматические избирательные системы
[ редактировать ]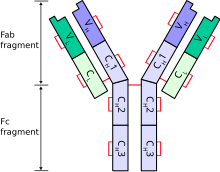
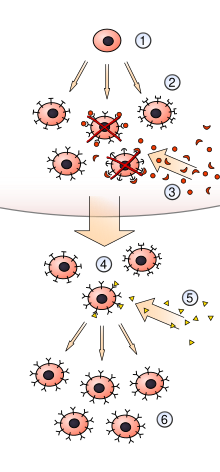
Эдельмана вдохновили успехи коллеги-лауреата Нобелевской премии. [12] Фрэнк Макфарлейн Бернет и его теория клональной селекции (CST) приобретенного антигенного иммунитета путем дифференциальной амплификации ранее существовавших вариаций в пределах конечного пула лимфоцитов иммунной системы . Популяция вариантов лимфоцитов в организме отражает различные популяции организмов в экологии. Существующее ранее разнообразие является двигателем адаптации в эволюции популяций.
«Из эволюционной и иммунологической теории ясно, что перед лицом неизвестного будущего фундаментальным требованием для успешной адаптации является уже существующее разнообразие». [13] - Джеральд М. Эдельман (1978)
Эдельман признает объяснительный диапазон использования Бернетом дарвиновских принципов при описании работы иммунной системы - и обобщает этот процесс на все клеточные популяции организма. Он также приходит к тому, чтобы рассматривать проблему как проблему распознавания и памяти с биологической точки зрения, где различие и сохранение «я» от «чужого» жизненно важно для целостности организма.
Нейрондарвинизм, как TNGS, представляет собой теорию отбора групп нейронов, которая переосмысливает фундаментальные концепции теоретического подхода Дарвина и Бернета. Нейронный дарвинизм описывает развитие и эволюцию мозга млекопитающих и его функционирование путем распространения дарвиновской парадигмы на тело и нервную систему.
Антитела и NCAM – новое понимание соматических селективных систем
[ редактировать ]Эдельман был медицинским исследователем, физико-химиком , иммунологом и начинающим нейробиологом, когда в 1972 году ему была присуждена Нобелевская премия по физиологии и медицине (совместно с Родни Портером из Великобритании). Часть премии Эдельмана была получена за его работу по выявлению химической структуры антител позвоночных путем расщепления ковалентных дисульфидных мостиков, соединяющих фрагменты компонентных цепей вместе, в результате чего была обнаружена пара двухдоменных легких цепей и четырехдоменных тяжелых цепей. Последующий анализ показал, что концевые домены обеих цепей являются вариабельными доменами, ответственными за распознавание антигена. [14]
Работа Портера и Эдельмана раскрыла молекулярные и генетические основы, лежащие в основе формирования разнообразия антител в иммунной системе. Их работа подтвердила более ранние идеи о ранее существовавшем разнообразии иммунной системы, выдвинутые датским иммунологом-новатором Нильсом К. Йерном (23 декабря 1911 г. - 7 октября 1994 г.); а также поддержка работы Фрэнка Макфарлейна Бернета, описывающей, как лимфоциты, способные связываться со специфическими чужеродными антигенами, дифференциально амплифицируются путем клонального размножения выбранных ранее существовавших вариантов после открытия антигена.
Эдельман черпал вдохновение из механо-химических аспектов взаимодействия антиген/антитело/лимфоцит в отношении распознавания себя-не-я; дегенеративная популяция лимфоцитов в их физиологическом контексте; и биотеоретические основы этой работы в дарвиновских терминах.
К 1974 году Эдельман почувствовал, что иммунология прочно утвердилась на прочной теоретической основе в описательном смысле, была готова к количественным экспериментам и могла стать идеальной моделью для изучения процессов эволюционного отбора в течение наблюдаемого периода времени. [15]
Его исследования взаимодействий иммунной системы развили в нем понимание важности клеточной поверхности и встроенных в мембрану молекулярных механизмов взаимодействия с другими клетками и субстратами. Эдельман продолжил развивать свои идеи топобиологии вокруг этих механизмов, а также их генетической и эпигенетической регуляции в условиях окружающей среды.
Во время набега на молекулярную эмбриологию и нейробиологию в 1975 году Эдельман и его команда выделили первую молекулу адгезии нервных клеток (N-CAM), одну из многих молекул, которые удерживают нервную систему животных вместе. N-CAM оказалась важной молекулой, управляющей развитием и дифференцировкой групп нейронов в нервной системе и мозге во время эмбриогенеза . К изумлению Эдельмана, генетическое секвенирование показало, что N-CAM был предком антител позвоночных. [16] образуется в результате ряда событий дупликации всего генома у происхождения позвоночных [17] это дало начало целому суперсемейству генов иммуноглобулинов .
Эдельман предположил, что молекула N-CAM, которая используется для самораспознавания и сцепления между нейронами нервной системы, дала начало их эволюционным потомкам, антителам, которые развили распознавание себя-не-собственного посредством адгезии антигена у истоков позвоночных. Иммунная система, основанная на антителах. Если клональный отбор был способом работы иммунной системы, возможно, он был наследственным и более общим – и действовал в эмбрионе и нервной системе.
Разнообразие биологических систем – вырождение, сложность, устойчивость и способность к развитию.
[ редактировать ]


Вырождение и его связь с вариациями — ключевая концепция нейронного дарвинизма. Чем больше мы отклоняемся от идеальной формы, тем больше у нас возникает искушение описать отклонения как несовершенства. Эдельман, с другой стороны, открыто признает структурную и динамическую изменчивость нервной системы. Ему нравится противопоставлять различия между избыточностью в инженерной системе и вырождением в биологической системе. Он продолжает демонстрировать, как «шум» вычислительного и алгоритмического подхода на самом деле полезен для соматической селективной системы, предоставляя широкий и вырожденный набор потенциальных элементов распознавания. [18]
Аргумент Эдельмана заключается в том, что в инженерной системе
- возникла известная проблема
- логичное решение найдено
- изобретение создано для реализации решения проблемы
Чтобы обеспечить надежность решения, критически важные компоненты копируются в виде точных копий. Резервирование обеспечивает отказоустойчивое резервное копирование в случае катастрофического отказа важного компонента, но это тот же ответ на ту же проблему после замены.
Если проблема предсказуема и известна заранее, резервирование работает оптимально. Но биологические системы сталкиваются с открытой и непредсказуемой ареной пространственно-временных событий, о которых они ничего не знают. В этой области избыточность терпит неудачу — ответ может быть разработан не для той проблемы.
Вариации способствуют вырождению; дегенерация обеспечивает соматическим селективным системам более одного способа решения проблемы и склонность повторно использовать решение для других проблем. Это свойство вырождения делает систему более адаптивно устойчивой перед лицом непредвиденных обстоятельств: когда одно конкретное решение неожиданно терпит неудачу, существуют другие незатронутые пути, которые можно использовать для достижения той же цели. Вначале Эдельман тратит много времени на противопоставление вырождения и избыточности, восходящих и нисходящих процессов, а также селекционистских и инструктистских объяснений биологических явлений.
Отказ от вычислительных моделей, кодов и двухточечных соединений
[ редактировать ]Эдельман был хорошо осведомлен о более ранних дебатах в иммунологии между учеными, которые считали, что лимфоциты иммунной системы узнали или получили инструкции об антигене, а затем разработали ответ; и селекционисты, которые считали, что лимфоциты уже содержат ответ на антиген в существующей популяции, который дифференциально усиливается внутри популяции при контакте с антигеном. И он прекрасно понимал, что на стороне селекционера есть доказательства.
Теоретический подход Эдельмана в нейронном дарвинизме был задуман в противовес нисходящим алгоритмическим, вычислительным и инструктистским подходам к объяснению нейронных функций. Вместо этого Эдельман пытается обратить проблемы этой парадигмы в свою пользу; тем самым подчеркивая разницу между восходящими процессами, которые мы видим в биологии, и нисходящими процессами, которые мы видим в инженерных алгоритмах. Он рассматривает нейроны как живые организмы, работающие кооперативно и конкурентно в рамках своей локальной экологии , и отвергает модели, которые рассматривают мозг как компьютерные чипы или логические элементы в алгоритмически организованной машине .
Приверженность Эдельмана дарвиновским основам биологии, его новое понимание эволюционных взаимоотношений между двумя молекулами, с которыми он работал, а также его опыт работы в области иммунологии привели к тому, что он стал все более критичным и неудовлетворенным попытками описать работу нервной системы и мозга. в вычислительном или алгоритмическом плане.
Эдельман категорически отвергает вычислительные подходы к объяснению биологии как небиологической. Эдельман признает, что в нервной системе позвоночных существует сохранение филогенетической организации и структуры, но также указывает на изобилие локального естественного разнообразия, изменчивости и вырождения. Это изменение внутри нервной системы разрушительно для теорий, основанных на строгой двухточечной связи, вычислениях или логических схемах, основанных на кодах. Попытки понять этот шум создают трудности для нисходящих алгоритмических подходов и отрицают фундаментальные факты биологической природы проблемы.
Эдельман понял, что проблематичный и раздражающий шум парадигмы вычислительной схемологики можно по-новому интерпретировать с точки зрения популяционной биологии – где это изменение в сигнале или архитектуре на самом деле было двигателем изобретательности и устойчивости с точки зрения селекциониста.
Завершение программы Дарвина – проблемы эволюционной морфологии и морфологии развития
[ редактировать ]В «Топобиологии» Эдельман размышляет о поисках Дарвином связей между морфологией и эмбриологией в его теории естественного отбора . Он выделяет четыре нерешенные проблемы в развитии и эволюции морфологии, которые Дарвин считал важными: [19]
- Объяснение конечного числа строений тела, проявленных начиная с докембрия .
- Объяснение крупномасштабных морфологических изменений за относительно короткие периоды геологического времени.
- Понимание размеров тела и основы аллометрии .
- Как адаптивная приспособленность может объяснить отбор, приводящий к появлению сложных структур тела.
Позже, в книге «Яркий воздух, яркий огонь » Эдельман описывает то, что он называет программой Дарвина для получения полного понимания правил поведения и формы в эволюционной биологии. [20] Он выделяет четыре необходимых требования:
- Отчет о влиянии наследственности на поведение, а поведение — на наследственность.
- Отчет о том, как отбор влияет на поведение – и как поведение влияет на отбор.
- Отчет о том, как поведение обеспечивается и ограничивается морфологией.
- Отчет о том, как морфогенез происходит в развитии и эволюции.
Важно отметить, что эти требования напрямую не связаны с генами, а с точки зрения наследственности. Это понятно, если учесть, что сам Дарвин, похоже, не осознавал напрямую важность менделевской генетики . Ситуация изменилась к началу 1900-х годов: неодарвиновский синтез объединил популяционную биологию менделевской наследственности с дарвиновским естественным отбором. К 1940-м годам было показано, что эти теории взаимно согласуются и согласуются с палеонтологией и сравнительной морфологией. Теория стала известна как современный синтез на основании названия книги «Эволюция: современный синтез», вышедшей Джулиана Хаксли в 1942 году . [21]
Современный синтез по-настоящему начался с открытия структурной основы наследственности в форме ДНК. Современный синтез значительно ускорился и расширился с развитием геномных наук, молекулярной биологии, а также с развитием вычислительных методов и возможностей моделирования динамики популяций. Но биологам, занимающимся эволюционным развитием, не хватало чего-то очень важного... – и это было включение одной из основополагающих отраслей биологии, эмбриологии. Четкое понимание пути от зародыша к зиготе , эмбриону , потомству и взрослой особи было недостающим компонентом синтеза. Эдельман и его команда располагались во времени и пространстве, чтобы в полной мере извлечь выгоду из этих технических разработок и научных задач – по мере того, как его исследования продвигались все глубже и глубже в клеточные и молекулярные основы нейрофизиологических аспектов поведения и познания с дарвиновской точки зрения.
Эдельман по-новому интерпретирует цели «программы Дарвина» с точки зрения современного понимания генов, молекулярной биологии и других наук, которые были недоступны Дарвину. Одна из его целей — согласовать взаимоотношения между генами в популяции (геноме), которые лежат в зародышевой линии (сперматозоид, яйцеклетка и оплодотворенная яйцеклетка); и особи в популяции, у которых развиваются дегенеративные фенотипы (сома) по мере превращения из эмбриона во взрослую особь, которая в конечном итоге будет производить потомство, если приспособится. Отбор действует на фенотипы (сома), но эволюция происходит внутри генома вида (зародыш).
Эдельман следует за работой весьма влиятельного американского генетика и биолога-эволюциониста Ричарда Левонтина (29 марта 1929 г. – 4 июля 2021 г.), черпая особое вдохновение из его книги 1974 года « Генетическая основа эволюционных изменений» . [22] Эдельман, как и Левонтин, ищет полного описания преобразований (Т), которые уводят нас от: [23]
- Геном-зародыш (зиготы) – вклады отцовских и материнских генов рекомбинируются в оплодотворенной яйцеклетке вместе с материнским запасом белков, мРНК и других компонентов развития, но вновь сформированный диплоидный генетический комплемент особей не контролирует зигота еще; его необходимо активировать или внедрить в зиготы, продолжая метаболизм и физиологию, унаследованные от матери. Вскоре после рекомбинации зигота проходит трансформацию (Т1) до такой степени, что генетический контроль над зиготой передается индивидууму.
- Фенотип-сома (эмбрион) – эмбрион, который трансформируется (Т2) по правилам, регулирующим взаимоотношения генов, клеточного поведения и эпигенетических случайностей природы, в
- Фенотип-сома (взрослый) – взрослый человек, который производит потомство (Т3) с другой особью для объединения новой генетической рекомбинации путем введения каждой гаметы в виде
- Геном-зародыш (гаметы) – сперматозоид и яйцеклетка, содержащие гаплоидный генетический вклад каждого родителя, который трансформируется (Т4)...
- Геном-зародыш (зигота) — диплоидный набор генов в оплодотворенной яйцеклетке, которая вскоре станет новой индивидуальной зиготой.
Исследование Левонтином этих трансформаций между геномным и фенотипическим пространствами проводилось с точки зрения ключевого давления отбора, которое формирует организм в геологических эволюционных временных масштабах; но подход Эдельмана более механический и действует здесь и сейчас – он фокусируется на генетически ограниченной механохимии процессов отбора, которые управляют эпигенетическим поведением со стороны клеток внутри эмбриона и взрослого человека на протяжении всего времени развития.
Механохимия, мезенхима и эпителий – CAM и SAM в морфорегуляторном пространстве-времени
[ редактировать ]
Выделение Эдельманом NCAM привело его к теоретизированию о роли молекул клеточной адгезии (CAM) и молекул адгезии субстрата (SAM) в формировании плана тела животных как в реальном времени, так и в течение эволюционного времени. Топобиология в первую очередь посвящена этому вопросу, который лежит в основе понимания нейронного дарвинизма и формирования первичного репертуара TNGS.
В своей гипотезе о регуляторе Эдельман выдвигает гипотезу о роли молекул клеточной поверхности в эмбриогенезе и о том, как изменение экспрессии этих молекул во времени и месте внутри эмбриона может управлять развитием паттерна. [24] Позже он расширит эту гипотезу до гипотезы морфорегуляции. [25] Он описывает популяции эмбриональных клеток как организованные в виде мезенхимы или эпетейлии.
Эдельман характеризует два типа населения следующим образом:
- Эпителий – популяция клеток, организованных в когерентные ткани с хорошо устоявшимися паттернами САМ; а также стабильный характер адгезии субстрата между клетками и внеклеточным матриксом.
- Мезенхима - популяция слабо связанных и мигрирующих клеток, которые втянули (или локализовали) свои молекулы CAM и SAM так, что они могут следовать гомофильным и гетерофильным градиентам внутри других популяций клеток эмбриона.
Он рассматривает цикл, управляемый CAM и SAM, в котором популяции клеток трансформируются взад и вперед между мезенхимой и эпителием посредством эпителиально-мезенхимальных трансформаций. [26] по мере того, как развитие эмбриона переходит в стадию плода. Экспрессия CAM и SAM находится под генетическим контролем, но распределение этих молекул на клеточной мембране и внеклеточном матриксе исторически зависит от эпигенетических событий, служащих одной из основных основ для создания уже существующего разнообразия в нервной системе и другие ткани.
Генетический вопрос развития
[ редактировать ]Необходимо рассмотреть множество вопросов развития, но Эдельман способен кратко изложить проблему таким образом, чтобы показать ему четкий путь объяснения вперед. Вопрос генетики развития определяет проблему – и теоретический подход для него.
«Как одномерный генетический код определяет трехмерное животное?» [27] - Джеральд М. Эдельман, из словаря топобиологии.
К 1984 году Эдельман будет готов ответить на этот вопрос и объединить его со своими более ранними идеями о дегенерации и соматическом отборе в нервной системе. Эдельман вернулся к этому вопросу в «Топобиологии» и объединил его с эволюционным подходом в поисках всеобъемлющей теории формирования и эволюции плана тела.
Гипотеза регулятора
[ редактировать ]В 1984 году Эдельман опубликовал свою гипотезу регулятора действия CAM и SAM в развитии и эволюции строения тела животных.
Эдельман повторил эту гипотезу в своей книге «Нейронный дарвинизм» в поддержку механизмов вырожденного формирования групп нейронов в первичном репертуаре. Гипотеза регулятора в первую очередь касалась действия CAM. Позже он расширил эту гипотезу в «Топобиологии» , включив в нее гораздо более разнообразный и всеобъемлющий набор морфорегуляторных молекул.
Эволюционный вопрос
[ редактировать ]Эдельман понял, что для того, чтобы по-настоящему завершить программу Дарвина, ему необходимо связать вопрос развития с более крупными проблемами эволюционной биологии.
«Как ответ на вопрос генетики развития (см.) согласуется с относительно быстрыми изменениями формы, происходящими за относительно короткий эволюционный период?» [28] - Джеральд М. Эдельман, из словаря топобиологии.
Гипотеза морфорегулятора
[ редактировать ]Вскоре после публикации своей гипотезы о регуляторе Эдельман расширил свое видение формирования паттернов во время эмбриогенеза и попытался связать его с более широкой эволюционной структурой. Его первая и главная цель — ответить на вопрос генетики развития, за которым следует вопрос эволюции, ясным, последовательным и последовательным образом.
TNGS – теория отбора групп нейронов
[ редактировать ]Мотивацией Эдельмана к разработке теории отбора групп нейронов (TNGS) было разрешение «ряда очевидных противоречий в наших знаниях о развитии, анатомии и физиологических функциях центральной нервной системы». [29] Актуальной проблемой для Эдельмана было объяснение категоризации восприятия без ссылки на центрального наблюдающего гомункула или «предположение, что мир заранее организован информационным образом». [29]
Освободиться от требований, требований и противоречий модели обработки информации; Эдельман предполагает, что перцептивная категоризация осуществляется путем отбора групп нейронов, организованных в различные сети, реакции которых по-разному усиливаются в сочетании с гедонистической обратной связью в течение опыта, изнутри огромной популяции групп нейронов, сталкивающихся с хаотичным набором сенсорных сигналов. вклад различной степени значимости и значимости для организма.
Эдельман категорически отвергает идею гомункула , описывая его как «близкого родственника электрика и нейронного декодера», артефакты централизованной наблюдателем нисходящей логики проектирования подходов к обработке информации. Эдельман справедливо отмечает, что «вероятно можно с уверенностью предположить, что большинство нейробиологов сочли бы гомункула, а также дуалистические решения (Поппер и Экклс, 1981) проблем субъективного сообщения выходящими за рамки научного рассмотрения». [30]
Необходимые критерии селекционистской теории высших функций мозга
[ редактировать ]Первый теоретический вклад Эдельмана в нейронный дарвинизм был сделан в 1978 году, когда он предложил свой групповой отбор и фазовую реентерабельную передачу сигналов . [31] Эдельман выдвигает пять необходимых требований, которым должна удовлетворять биологическая теория высших функций мозга. [32]
- Теория должна соответствовать областям эмбриологии, нейроанатомии и нейрофизиологии.
- Теория должна учитывать обучение, память и временной отзыв в распределенной системе.
- Теория должна учитывать, как память обновляется на основе опыта реального времени.
- Теория должна объяснить, как высшие системы мозга опосредуют опыт и действия.
- Теория должна учитывать необходимые, если не достаточные, условия возникновения осознания.
Организация теории TNGS
[ редактировать ]Нейрондарвинизм разделяет объяснение TNGS на три части: соматический отбор, эпигенетические механизмы и глобальные функции. Первые две части посвящены тому, как вариации возникают в результате взаимодействия генетических и эпигенетических событий на клеточном уровне в ответ на события, происходящие на уровне развивающейся нервной системы животного. Третья часть пытается построить согласованную во времени модель глобально единой когнитивной функции и поведения, которая возникает снизу вверх посредством взаимодействия групп нейронов в реальном времени.
Эдельман свел ключевые идеи теории TNGS в три основных принципа:
- Первичный репертуар – онтогенетическое формирование и отбор групп нейронов;
- Вторичный репертуар – поведенческий и эмпирический отбор, приводящий к изменению силы связей между синаптическими популяциями, связывающими между собой группы нейронов;
- Реентерабельная передача сигналов – синхронное вовлечение взаимно связанных групп нейронов внутри сенсомоторных карт в ансамбли когерентной глобальной активности.
Первичный репертуар формируется в период от начала нейруляции до окончания апоптоза. Вторичный репертуар охватывает период синаптогенеза и миелинизации, но продолжает демонстрировать пластичность развития на протяжении всей жизни, хотя и в меньшей степени по сравнению с ранним развитием.
Эти два репертуара посвящены проблеме взаимосвязи между генетическими и эпигенетическими процессами при определении общей архитектуры нейроанатомии, стремясь примирить природу, воспитание и изменчивость при формировании окончательного фенотипа любой отдельной нервной системы.
Не существует двухточечной связи, по которой нейронный код проходит через вычислительную логическую схему, которая доставляет результат в мозг, потому что
- во-первых, доказательства не подтверждают такое мнение в такой степени, которая не является проблематичной,
- во-вторых, шум в системе слишком велик, чтобы нейронный код был когерентным,
- и в-третьих, гены могут только способствовать процессам развития и ограничивать их; не определить их во всех подробностях.
Вариации — неизбежный результат динамики развития.
Реентерабельная передача сигналов - это попытка объяснить, как «происходят когерентные временные корреляции ответов листов сенсорных рецепторов, двигательных ансамблей и взаимодействующих групп нейронов в разных областях мозга». [33]
Первичный репертуар – развивающий отбор
[ редактировать ]Первый принцип TNGS касается событий, которые находятся на эмбриональном этапе и продолжаются до неонатального периода. Эта часть теории пытается объяснить уникальное анатомическое разнообразие мозга даже между генетически идентичными людьми. Первый принцип предполагает развитие первичного репертуара дегенеративных групп нейронов с разнообразными анатомическими связями, устанавливаемыми посредством исторических случайностей первичных процессов развития. Он стремится дать объяснение тому, как разнообразие фенотипов организма групп нейронов возникает из генотипа посредством генетических и эпигенетических влияний, которые механо-химически проявляются на поверхности клеток и определяют связность.
Эдельман называет следующие факторы жизненно важными для формирования первичного репертуара групп нейронов, но также способствующими их анатомическому разнообразию и вариациям:
- Деление клеток – при формировании популяций нейронов происходят повторяющиеся циклы деления клеток.
- Гибель клеток – в популяциях нейронов происходит обширная заранее запрограммированная гибель клеток, которая происходит посредством апоптоза.
- Расширение и устранение процесса - исследовательское исследование эмбриональной среды развивающимися нейронами включает расширение и устранение процесса, поскольку нейроны обнаруживают молекулярные градиенты на мембранах поверхности соседних клеток и субстрате внеклеточного матрикса.
- Действие CAM и SAM – механохимия молекул клеточной и поверхностной адгезии играет ключевую роль в миграции и связности нейронов, поскольку они образуют группы нейронов в общей распределенной популяции.
Два ключевых вопроса относительно этой проблемы, на которые Эдельман пытается ответить «с точки зрения генетических и эпигенетических событий развития», заключаются в следующем: [34]
- «Как одномерный генетический код определяет трехмерное животное?»
- «Как ответ на этот вопрос согласуется с возможностью относительно быстрых морфологических изменений за относительно короткие периоды эволюционного времени?»
Вторичный репертуар – экспериментальный отбор
[ редактировать ]Второй принцип TNGS касается постнатальных событий, которые управляют развитием вторичного репертуара синаптических связей между популяциями нейрональных групп более высокого порядка, формирование которых обусловлено поведенческим или экспериментальным отбором, действующим на синаптические популяции внутри и между группами нейронов. Представление Эдельмана о вторичном репертуаре во многом заимствовано из работ Жана-Пьера Шанже и его коллег Филиппа Куррежа и Антуана Даншена , а также из их теории избирательной стабилизации синапсов. [35]
Синаптическая модификация
[ редактировать ]Как только базовая разнообразная анатомическая структура первичного репертуара групп нейронов заложена, она более или менее фиксируется. Но, учитывая многочисленную и разнообразную коллекцию нейронных групповых сетей, обязательно найдутся функционально эквивалентные, хотя и анатомически неизоморфные нейронные группы и сети, способные реагировать на определенные сенсорные сигналы. Это создает конкурентную среду, в которой группы нейронов, умеющие реагировать на определенные входные сигналы, «дифференцированно усиливаются» за счет повышения синаптической эффективности выбранной сети групп нейронов. Это приводит к увеличению вероятности того, что та же сеть будет реагировать на аналогичные или идентичные сигналы в будущем. Это происходит за счет укрепления синапсов между нейронами. Эти корректировки позволяют добиться нейронной пластичности в довольно короткие сроки.
Возвращение в атмосферу
[ редактировать ]Третий и последний принцип TNGS – это вход в атмосферу. Реентерабельная передача сигналов «основана на существовании взаимно связанных нейронных карт». [33] Эти топобиологические карты поддерживают и координируют реакции в реальном времени множества реагирующих сетей вторичного репертуара, как унимодальных, так и мультимодальных, а их взаимные реентерабельные связи позволяют им «поддерживать и поддерживать пространственно-временную непрерывность в ответ на сигналы реального мира». [33]
Последняя часть теории пытается объяснить, как мы испытываем пространственно-временную последовательность в нашем взаимодействии со стимулами окружающей среды. Эдельман назвал это « возвращением » и предлагает модель повторной передачи сигналов, согласно которой дизъюнктивная мультимодальная выборка одного и того же стимулирующего события, коррелированная во времени, делает возможным устойчивое физиологическое вовлечение распределенных групп нейронов во временно стабильные глобальные поведенческие единицы действия или восприятия. Другими словами, несколько групп нейронов могут использоваться для параллельной выборки данного набора стимулов и взаимодействия между этими дизъюнктивными группами с возникающей задержкой.
Расширенная теория выбора групп нейронов – гипотеза динамического ядра
[ редактировать ]После публикации « Нейронного дарвинизма » Эдельман продолжил развивать и расширять свою теорию TNGS, а также свою гипотезу регулятора. Эдельман занимался морфологическими проблемами топобиологии и начал расширять теорию TNGS в «Вспомнившемся настоящем» . Периодически в течение прошедших лет Эдельман публиковал новую обновленную информацию о своей теории и достигнутом прогрессе.
В «Вспомнившемся настоящем » Эдельман заметил, что центральная нервная система млекопитающих, по-видимому, имеет две отдельные морфологически организованные системы: одна — лимбическая система ствола мозга, которая в первую очередь отвечает за «аппетитное, потребительское и защитное поведение»; [36] Другая система - это таламокортикальная система с высокой степенью реентерабельности, состоящая из таламуса, а также «первичных и вторичных сенсорных областей и ассоциативной коры». [36] которые «тесно связаны с экстерорецепторами и тщательно и широко картированы полимодальным образом». [36]
Лимбико-стволовая система – внутренний мир сигналов
[ редактировать ]Нейронная анатомия системы гедонической обратной связи находится в стволе мозга, вегетативной, эндокринной и лимбической системах . Эта система передает свою оценку состояния внутренних органов остальной части центральной нервной системы. Эдельман называет эту систему лимбически-стволовой системой .
Таламокортикальная система – внешний мир сигналов
[ редактировать ]Таламус — это ворота в неокортекс для всех чувств, кроме обоняния. Спиноталамические тракты доставляют сенсорную информацию от периферии к таламусу, где мультимодальная сенсорная информация интегрируется и запускает быстрые подкорковые рефлекторные двигательные реакции через миндалевидное тело, базальные ганглии, гипоталамус и центры ствола мозга. Одновременно каждая сенсорная модальность также передается в кору параллельно для рефлексивного анализа более высокого порядка, мультимодальной сенсомоторной ассоциации и включения медленной модуляторной реакции, которая будет точно настраивать подкорковые рефлексы.
Корковые придатки – органы наследственности.
[ редактировать ]В книге «Вспомнившееся настоящее » Эдельман признает ограниченность своей теории TNGS для моделирования динамики временной последовательности двигательного поведения и памяти. Его ранние попытки создания автоматов репликации оказались неадекватными задаче объяснения секвенирования в реальном времени и интеграции групповых взаимодействий нейронов с другими системами организма. «Ни оригинальная теория, ни моделируемые автоматы распознавания не имеют достаточно подробного описания последовательного порядка событий во времени, опосредованного несколькими основными компонентами мозга, которые способствуют памяти, особенно в том, что касается сознания». [37] Эта проблема заставила его сосредоточиться на том, что он называл органами преемственности; мозжечок, базальные ганглии и гиппокамп.
Прием
[ редактировать ]Ранний обзор книги «Нейронный дарвинизм» в The New York Review of Books. [38] Исследование Исраэля Розенфилда вызвало живой отклик со стороны нейробиологического сообщества. [39] Взгляды Эдельмана можно было бы рассматривать как атаку на доминирующую парадигму вычислительных алгоритмов в когнитивной психологии и вычислительной нейробиологии, вызывая критику со многих сторон.
Было бы много жалоб на языковые трудности. Некоторые сочли бы Эдельмана высокомерным или вторгшимся в область нейробиологии из соседней молекулярной биологии. Были выдвинуты законные аргументы относительно того, сколько экспериментальных и наблюдательных данных было собрано в поддержку теории в то время. Или была ли теория вообще оригинальной или нет.
Но чаще всего критика сосредотачивается не на критике Эдельманом вычислительных подходов, а на том, является ли система Эдельмана действительно правильным дарвиновским объяснением. Тем не менее, «Нейронный дарвинизм» , как книга, так и концепция, получил довольно широкое признание критиков.
Одной из самых известных критических статей о нейронном дарвинизме стала критическая рецензия Фрэнсиса Крика » 1989 года «Нейронный эдельманизм . [40] Фрэнсис Крик основывал свою критику на том, что группы нейронов получают инструкции от окружающей среды, а не подвергаются слепым изменениям. В 1988 году нейрофизиолог Уильям Кальвин предположил истинную репликацию в мозге. [41] тогда как Эдельман выступал против идеи настоящих репликаторов в мозгу. Стивен Смоляр опубликовал еще один обзор в 1989 году. [42]
Англии и ее нейробиологическому сообществу пришлось бы полагаться на контрафактные копии книги до 1990 года, но как только книга прибыла на английские берега, британский социальный обозреватель и нейробиолог Стивен Роуз поспешил похвалить и критиковать ее идеи, написав: стиль, предположения и выводы. [43] Писатель New York Times » Джордж Джонсон опубликовал «Эволюцию между ушами», критическую рецензию на книгу Джеральда Эдельмана 1992 года « Блестящий воздух, блестящий огонь . [44] В 2014 году Джон Хорган написал письмо Джеральду Эдельману в журнале Scientific American , подчеркнув его высокомерие, блестящие способности и своеобразный подход к науке. [45]
предположил Чейз Херрманн-Пиллат , что Фридрих Хайек ранее предложил аналогичную идею в своей книге «Сенсорный порядок: исследование основ теоретической психологии» , опубликованной в 1952 году. [46] Среди других ведущих сторонников селекционистских предложений можно назвать Жан-Пьера Шанже (1973, 1985), [35] [47] Дэниел Деннетт и Линда Б. Смит . Обзоры работ Эдельмана будут продолжать публиковаться по мере распространения его идей.
Недавний обзор Фернандо, Сатмари и Хасбандса объясняет, почему нейронный дарвинизм Эдельмана не является дарвинистским, поскольку он не содержит единиц эволюции, как это определил Джон Мейнард Смит . Она является селекционистской в том смысле, что удовлетворяет уравнению Прайса , но в теории Эдельмана нет механизма, объясняющего, как информация может передаваться между группами нейронов. [48] Недавняя теория под названием «эволюционная нейродинамика», разработанная Эорсом Сатмари и Крисантой Фернандо, предложила несколько способов, с помощью которых в мозге может происходить истинная репликация. [49]
Эти нейронные модели были расширены Фернандо в более поздней статье. [50] В самой последней модели три механизма пластичности: i) мультипликативный STDP, ii) LTD и iii) гетеросинаптическая конкуренция ответственны за копирование паттернов связности из одной части мозга в другую. Точно такие же правила пластичности могут объяснить экспериментальные данные о том, как младенцы осуществляют причинное обучение в экспериментах, проведенных Элисон Гопник . Также было показано, что если добавить обучение Хебба к репликаторам нейронов, мощность нейронных эволюционных вычислений может фактически превзойти естественный отбор в организмах. [51]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Работа Родни Портера с ферментом папаином привела к расщеплению антитела на фрагменты Fab и Fc , а работа Джеральда Эдельмана привела к восстановлению дисульфидных мостиков и разделению молекулы на фрагменты легкой и тяжелой цепи. В совокупности эта работа позволила секвенировать и реконструировать структуру антител, что привело к присуждению Нобелевской премии по физиологии и медицине в 1972 году.
Цитаты
[ редактировать ]- ^ Эдельман и Портер 1972 .
- ^ Эдельман 1987b .
- ^ Маунткасл и Эдельман 1978 , с. 7-50, Организационный принцип церебральной функции: модульный модуль и распределенная система.
- ^ Маунткасл и Эдельман 1978 , с. 51-100, Групповой отбор и фазовая реентерабельная сигнализация: теория функций высшего мозга.
- ^ Дарвин 1859 .
- ^ Эдельман 1988 .
- ^ Эдельман 1989 .
- ^ Эдельман 1992 .
- ^ Эдельман и Тонони 2000 .
- ^ Эдельман 2004 .
- ^ Эдельман 2006 .
- ^ Бернет и Медавар 1960 .
- ^ Маунткасл и Эдельман 1978 , с. 56.
- ^ Эдельман 1972 .
- ^ Эдельман 1974 .
- ^ Эдельман 1987a .
- ^ Дехал и Бур 2005 .
- ^ Тонони, Спорнс и Эдельман 1999 .
- ^ Эдельман 1988 , с. 45.
- ^ Эдельман 1992 , Глава 5 Морфология и разум: завершение программы Дарвина.
- ^ Хаксли 1942 .
- ^ Левонтин 1974 .
- ^ Эдельман 1988 , с. 45-47.
- ^ Эдельман 1987b , с. 93-100, Нормативная гипотеза.
- ^ Эдельман 1988 , с. 127–172, Гипотеза морфорегулятора: механохимия, связанная с генетикой развития.
- ^ Эдельман 1988 , с. 67-71219.
- ^ Эдельман 1988 , с. 217.
- ^ Эдельман 1988 , с. 219.
- ^ Jump up to: а б Эдельман 1987b , с. 4.
- ^ Эдельман 1987b , с. 41.
- ^ Маунткасл и Эдельман 1978 , с. 51–100, Групповой отбор и фазовая реентерабельная передача сигналов: теория функций высшего мозга.
- ^ Маунткасл и Эдельман 1978 , с. 52.
- ^ Jump up to: а б с Эдельман 1987b , с. 5.
- ^ Эдельман 1987b , с. 75.
- ^ Jump up to: а б Шанжё, Курреж и Данчин, 1973 г.
- ^ Jump up to: а б с Эдельман 1989 , с. 152.
- ^ Эдельман 1989 , с. 112.
- ^ Розенфилд 1986 .
- ^ Янг, Аяла и Сентаготаи, 1987 .
- ^ Крик 1989 .
- ^ Кальвин 1988 .
- ^ Смоляр 1989 .
- ^ Роуз 1990 .
- ^ Джонсон 1992 .
- ^ Хорган 2014 .
- ^ Херрманн-Пиллат 2006 .
- ^ Changeux 1985 .
- ^ Фернандо, Сатмари и мужья 2012 .
- ^ Фернандо, Каришма и Сатмари 2008 .
- ^ Фернандо 2013 .
- ^ Фернандо, Гольдштейн и Сатмари 2010 .
Ссылки
[ редактировать ]Этот раздел может потребовать очистки Википедии , чтобы соответствовать стандартам качества . Конкретная проблема: удалить кавычки и лишний текст из параметров заголовка. ( Июль 2021 г. ) |
- Бернет, Фрэнк Макфарлейн; Медавар, Питер Брайан (1960). «Нобелевская премия по физиологии и медицине 1960 года» . NobelPrize.org .
- Кальвин, Уильям (24 июня 1988 г.). «Нейронный дарвинизм: теория отбора групп нейронов» . Наука .
- Шанжё, Жан-Пьер (1985). Нейронный человек – Биология разума . Перевод Лоуренса Кэри. Книги Пантеона , Нью-Йорк. ISBN 0-394-53692-4 .
- Шанжё, Жан-Пьер; Курреж, Филипп; Данчин, Антуан (1973). «Теория эпигенеза нейронных сетей путем избирательной стабилизации синапсов» . Учеб. Натл. акад. наук. США . 70 (10): 2974–2978. Бибкод : 1973PNAS...70.2974C . дои : 10.1073/pnas.70.10.2974 . ПМК 427150 . ПМИД 4517949 .
- Крик, Фрэнсис (1989). «Нейронный эдельманизм». Тенденции нейробиологии . 12 (7): 240–248. дои : 10.1016/0166-2236(89)90019-2 . ПМИД 2475933 . S2CID 3947768 .
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь . Д. Эпплтон и компания, Нью-Йорк, 1869 г.
- Дарвин, Чарльз (1872). Выражение эмоций у человека и животных – с предисловием Конрада Лоренца . Издательство Чикагского университета, Чикаго и Лондон, 1965. ISBN. 0-226-13656-6 .
- Дарвин, Чарльз (1887). Автобиография Чарльза Дарвина 1809–1882 гг. С восстановлением первоначальных упущений. Отредактировано, с приложением и примечаниями его внучки Норы Барлоу . WW Norton & Company , 1969. ISBN. 0-393-00487-2 .
- Дехал, Парамвир; Бур, Джеффри Л. (6 сентября 2005 г.). «Два раунда дупликации целого генома у предков позвоночных» . ПЛОС Биология . 3 (10): е314. дои : 10.1371/journal.pbio.0030314 . ISSN 1545-7885 . ПМК 1197285 . ПМИД 16128622 .
- Эдельман, Джеральд М. (1972). Ян Линдстен (ред.). Структура антител и молекулярная иммунология (В: Нобелевские лекции по физиологии и медицине, 1971–1980 гг.) . World Scientific Publishing Co. , Сингапур, 1992. ISBN. 978-9810207915 .
- Эдельман, Джеральд М. (1974). Джеральд М. Эдельман (ред.). Истоки и механизмы специфичности клональной селекции (в книге: Клеточная селекция и регуляция иммунного ответа) . Том. 29. Raven Press , Нью-Йорк, 1974. ISBN. 0-911216-71-5 .
{{cite book}}:|journal=игнорируется ( помогите ) - Эдельман, Джеральд М. (1992b). «Молекулярное распознавание в иммунной и нервной системах» . В Уордене, ФГ; Свази, JP; Адельман, Г. (ред.). Нейронауки: пути открытий, I. Бостон: И. Биркхойзер (опубликовано в 1990 г.). стр. 65–74. дои : 10.1007/978-1-4684-6817-5_4 . ISBN 978-1-4684-6817-5 .
- Эдельман, Джеральд М. (1984). «Клеточная адгезия и морфогенез: гипотеза регулятора» (PDF) . Труды Национальной академии наук США . 81 (5): 1460–1464. Бибкод : 1984PNAS...81.1460E . дои : 10.1073/pnas.81.5.1460 . ПМЦ 344856 . ПМИД 6584892 .
- Эдельман, Джеральд М. (1987a). «CAMs и Ig: клеточная адгезия и эволюционное происхождение иммунитета» . Иммунологические обзоры . 100 : 11–45. дои : 10.1111/j.1600-065x.1987.tb00526.x . ПМИД 3326819 . S2CID 24972419 .
- Эдельман, Джеральд М. (1987b). Нейронный дарвинизм – Теория отбора групп нейронов . Основные книги , Нью-Йорк. ISBN 0-465-04934-6 .
- Эдельман, Джеральд М. (1988). Топобиология – введение в молекулярную эмбриологию . Основные книги, Нью-Йорк. ISBN 978-0-465-08653-5 .
- Эдельман, Джеральд М. (1989). Запомненное настоящее – биологическая теория сознания . Основные книги, Нью-Йорк. ISBN 0-465-06910-Х .
- Эдельман, Джеральд М. (1992). Яркий воздух, яркий огонь – о деле разума . Basic Books, Inc. ISBN 0-465-05245-2 .
- Эдельман, Джеральд М. (1993). «Нейронный дарвинизм: отбор и реентерабельная передача сигналов в высших функциях мозга» (PDF) . Нейрон . 10 (2): 115–25. дои : 10.1016/0896-6273(93)90304-а . ПМИД 8094962 . S2CID 8001773 .
- Эдельман, Джеральд М. (1998). ГМ. Эдельман; JP Changuex (ред.). Построение изображения мозга (В: Мозг) . Рутледж, Taylor & Francisco Group, Лондон и Нью-Йорк, 2000. ISBN 978-0-7658-0717-5 .
- Эдельман, Джеральд М. (2004). Шире, чем небо – феноменальный дар сознания . Издательство Йельского университета . ISBN 978-0-300-10761-6 .
- Эдельман, Джеральд М. (2006). Вторая природа – наука о мозге и человеческие знания . Издательство Йельского университета. ISBN 978-0-300-12594-8 .
- Эдельман, Джеральд М.; Галли, Джозеф А. (2001). «Вырождение и сложность биологических систем» . Труды Национальной академии наук США . 98 (24): 13763–13768. Бибкод : 2001PNAS...9813763E . дои : 10.1073/pnas.231499798 . ПМК 61115 . ПМИД 11698650 .
- Эдельман, Джеральд М.; Галли, Джозеф А. (2013). «Повторный вход: ключевой механизм интеграции функций мозга» . Границы интегративной нейронауки . 7 (63): 63. doi : 10.3389/fnint.2013.00063 . ПМЦ 3753453 . ПМИД 23986665 .
- Эдельман, Джеральд М.; Портер, Родни Р. (1972). «Нобелевская премия по физиологии и медицине 1972 года» . NobelPrize.org .
- Эдельман, Джеральд М.; Тонони, Джулио (2000). Вселенная сознания – как материя становится воображением . Basic Books, Inc. ISBN 978-0-465-01377-7 .
- Эрикссон, Питер С.; и др. (1998). «Нейрогенез в гиппокампе взрослого человека» . Природная медицина . 4 (11): 1313–1317. дои : 10.1038/3305 . ПМИД 9809557 .
- Фернандо, К.; Каришма, КК; Сатмари, Э. (2008). «Копирование и эволюция топологии нейронов» . ПЛОС ОДИН . 3 (11): 3775. Бибкод : 2008PLoSO...3.3775F . дои : 10.1371/journal.pone.0003775 . ПМЦ 2582483 . ПМИД 19020662 .
- Фернандо, К.; Гольдштейн, Р.; Сатмари, Э. (2010). «Гипотеза нейронального репликатора» . Нейронные вычисления . 22 (11): 2809–2857. дои : 10.1162/NECO_a_00031 . ПМИД 20804380 . S2CID 17940175 .
- Фернандо, К.; Сатмари, Э.; Мужья, П. (2012). «Селекционистский и эволюционный подходы к функциям мозга: критическая оценка» . Границы вычислительной нейронауки . 6 (24): 24. дои : 10.3389/fncom.2012.00024 . ПМЦ 3337445 . ПМИД 22557963 .
- Фернандо, К. (2013). «От бликетов к синапсам: определение временных причинных сетей путем наблюдения» . Когнитивная наука . 37 (8): 1426–1470. дои : 10.1111/cogs.12073 . ПМИД 23957457 .
- Хайек, Ф.А. (1952). Сенсорный порядок: исследование основ теоретической психологии . Рутледж и Кеган Пол, Лондон. ISBN 0-226-32094-4 .
- Херрманн-Пиллат, Карстен (10 декабря 2006 г.). «Мозг, его сенсорный порядок и эволюционная концепция разума, о вкладе Хайека в эволюционную эпистемологию». Журнал социальных и эволюционных систем . 15 (2): 145–187. дои : 10.1016/1061-7361(92)90003-в . ССНР 950592 .
- Хилл, Шарлотта; Ван, Иихуа (2020). «Важность эпителиально-мезенхимального перехода и аутофагии в устойчивости рака к лекарствам» . Лекарственная устойчивость рака . 3 (1): 38–47. дои : 10.20517/cdr.2019.75 . ПМЦ 7100899 . ПМИД 32226927 .
- Хорган, Джон (22 мая 2014 г.). «Моя раздражительная встреча с покойным великим Джеральдом Эдельманом» . Научный американец . Проверено 30 апреля 2021 г.
- Хуттенлохер, PR (1990). «Морфометрическое исследование развития коры головного мозга человека». Нейропсихология . 28 (6): 517–527. дои : 10.1016/0028-3932(90)90031-i . ПМИД 2203993 . S2CID 45697561 .
- Хаксли, Джулиан (1942). Эволюция: современный синтез . Лондон, G. Allen & Unwin Ltd.
- Джонсон, Джордж (19 апреля 1992 г.). «Эволюция между ушами» . Нью-Йорк Таймс .
- Левонтин, Ричард К. (1974). Генетическая основа эволюционных изменений . Издательство Колумбийского университета . ISBN 0231033923 .
- Маунткасл, Вернон Бенджамин; Эдельман, Джеральд М. (1978). Внимательный мозг – корковая организация и группово-селективная теория функций высшего мозга . МТИ Пресс. ISBN 978-0-262-55007-9 .
- Роуз, Стивен (9 июня 1990 г.). «Рецензия: Дарвин о мозге» . www.newscientist.com . Новый учёный . Проверено 18 апреля 2021 г.
- Розенфилд, Израиль (9 октября 1986 г.). «Нейронный дарвинизм: новый подход к памяти и восприятию» . Нью-Йоркское обозрение книг . Проверено 27 апреля 2021 г.
- Смоляр, Стивен В. (1989). «Рецензия на Г. М. Эдельмана (рецензия на книгу)» . У Уильяма Дж. Клэнси; Стивен В. Смоляр; Марк Стефик (ред.). Созерцая разум: форум искусственного интеллекта . MIT Press (опубликовано в 1996 г.). стр. 431–446. ISBN 978-0-262-53119-1 . (первоначально опубликовано в журнале Artificial Intelligence 39 (1989) 121–139.)
- «Памяти: Джеральда Эдельмана (1929–2014)» . Новости и обзоры – Том 14, Выпуск 17 . Исследовательский институт Скриппса (TSRI). 2 июня 2014 года . Проверено 21 июля 2021 г.
- Тонони, Джулио; Спорнс, Олаф; Эдельман, Джеральд М. (1999). «Меры вырождения и избыточности в биологических сетях» . ПНАС . 96 (6): 3257–3262. Бибкод : 1999PNAS...96.3257T . дои : 10.1073/pnas.96.6.3257 . ЧВК 15929 . ПМИД 10077671 .
- Янг, Джей Зи; Айяла, Франсиско Дж.; Сентаготаи, Дж. (12 марта 1987 г.). «Нейронный дарвинизм: обмен (ответ Исраэля Розенфилда)» . Нью-Йоркское обозрение книг . Проверено 27 апреля 2021 г.
Дальнейшее чтение
[ редактировать ]- Как думает мозг: развитие интеллекта тогда и сейчас Уильям Х. Кальвин
- Нейрогенез в мозге взрослого человека
Внешние ссылки
[ редактировать ]- Полное собрание сочинений Чарльза Дарвина онлайн
- Викиверситет – Нейронауки
- Викиверситет – Основы нейронауки
- Викиверситет – Введение в негенетический дарвинизм
- Веб-страница Уильяма Кальвина
- Веб-страница Дэниела Деннета
- Веб-страница Крисанты Фернандо
- Научно-популярные книги 1978 года
- Научно-популярные книги 1987 года
- Научно-популярные книги 1988 года
- Научно-популярные книги 1989 года
- Научно-популярные книги 1992 года
- 2000 научно-популярных книг
- Научно-популярные книги 2004 года
- Научно-популярные книги 2006 г.
- Книги по биологии
- Сознание
- дарвинизм
- Неврология