Сирения
| Сирения Временной диапазон: ранний эоцен – голоцен ,
| |
|---|---|

| |
| По часовой стрелке сверху слева: Вест-индийский ламантин , африканский ламантин , стеллерова морская корова , дюгонь , амазонский ламантин. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Клэйд : | Пэнунгулатоморфа |
| Грандордер: | Пэнунгулата |
| Зеркальный порядок: | Тетитерия |
| Заказ: | Сирения Иллигер , 1811 г. |
| Семьи | |

| |
| Вест-Индский - зеленым, амазонский - красным, африканский - оранжевым, дюгонь - синим, стеллерова морская корова - в кружке (желтый) | |
| Синонимы [ 1 ] | |
Сирении , населяющих ( / s aɪ ˈ r iː n i . ə / ), обычно называемые морскими коровами или сиренами , представляют собой отряд полностью водных травоядных млекопитающих болота, реки, устья рек, морские водно-болотные угодья и прибрежные морские воды. Существующие сирени состоят из двух отдельных семейств : Dugongidae ( дюгони и ныне вымершая стеллерова морская корова ) и Trichechidae ( ламантины , а именно амазонский ламантин, западно-индийский ламантин и западноафриканский ламантин), всего четыре вида. [ 2 ] Семейства Protosirenidae (эоценовые сирени ) и Prorastomidae (наземные сирени) вымерли. Сирены отнесены к кладе Paenungulata , наряду со слонами и даманами , и появились в эоцене 50 миллионов лет назад (млн лет назад). Dugongidae отделились от Trichechidae в позднем эоцене или начале олигоцена (30–35 млн лет назад). [ 3 ] [ 4 ]
Сирены вырастают от 2,5 до 4 метров (от 8,2 до 13,1 футов) в длину и до 1500 килограммов (3300 фунтов) в весе. Недавно вымершая стеллерова морская корова была самой крупной из когда-либо живших сирен, достигая длины 10 метров (33 фута) и веса от 5 до 10 тонн (от 5,5 до 11,0 коротких тонн). [ 2 ]
Сирены имеют большое веретенообразное тело, которое уменьшает сопротивление воды, и тяжелые кости, которые действуют как балласт, противодействуя плавучести их жира . Они имеют тонкий слой жира и, следовательно, чувствительны к колебаниям температуры, которые вызывают миграции, когда температура воды падает слишком низко. Сирены движутся медленно, обычно двигаясь со скоростью 8 километров в час (5,0 миль в час), но они могут достигать скорости 24 километра в час (15 миль в час) короткими очередями. Своими сильными губами они вытягивают морскую траву , съедая 10–15% веса своего тела в день.
Во время дыхания сирены держат над поверхностью только ноздри, иногда вставая для этого на хвост. Обычно они обитают в теплых, мелких прибрежных водах или реках. В основном они травоядны, но известно, что они поедают животных, таких как птицы и медузы. Самцы обычно спариваются более чем с одной самкой и могут собираться в токи для спаривания. Сирены K выбираются и проявляют родительскую заботу .

Мясо, масло, кости и шкуры имеют коммерческую ценность. Смертность часто вызвана непосредственной охотой человека или другими причинами, вызванными деятельностью человека, такими как разрушение среды обитания , запутывание в рыболовных снастях и столкновения плавсредств . Стеллерова морская корова вымерла из-за чрезмерной охоты в 1768 году.
Таксономия
[ редактировать ]Этимология
[ редактировать ]Сирении, обычно сирены , также упоминаются под общим названием «сирены» , происходящим от сирен из греческой мифологии . [ 5 ] [ 6 ] [ 7 ]
Классификация
[ редактировать ]Сирены классифицируются в когорте Afrotheria в кладе Paenungulata , наряду с Proboscidea (слоны), Hyracoidea (даманы), Embrithopoda , Desmostylia и Afroinsectiphilia . [ 8 ] [ 9 ] [ 10 ] [ 11 ] Эта клада была впервые установлена Джорджем Гейлордом Симпсоном в 1945 году на основе анатомических данных, таких как тестиконди и подобное развитие плода . Paenungulata, наряду с Afrotheria, являются одной из наиболее хорошо изученных клад млекопитающих в молекулярной филогении. [ 12 ] Sirenia, Proboscidae и Desmotylia сгруппированы вместе в кладе Tethytheria . На основании морфологического сходства ранее считалось, что Tethytheria, Perissodactyla и Hyracoidea сгруппированы вместе как Altungulata , но это было опровергнуто молекулярными данными. [ 9 ]
| Кладограмма, показывающая положение Сирений внутри Афротерии на основе молекулярных данных. [ 8 ] |
Семьи Сирении
[ редактировать ]† = Вымершие
Семейство Dugongidae : [ 13 ]
- Genus Dugong
- Д. дюгон
- Род † Анизосирен.
- † А. Панноника
- Род † Индосирен.
- † И. яваненсе
- Род † Бхаратисирен.
- † Б. указывает
- Род † Callistosiren. [ 14 ]
- † C. boriquensis
- Род † Кренатосирен.
- † К. Олсени
- Род † Користосирен.
- † К. Варгези
- Род † Диоплотерий.
- † Д. аллисони
- † Д. манигуальти
- Род † Домнингия.
- † Д. содхэ
- Род † Кутчисирен.
- † К. цилиндрическая
- Род † Наносирен.
- † Номер гарсии
- † Н. Санчес
- Род † Rytiodus.
- † Р. капгранди
- † Р. хили
- Род † Xenosiren.
- † X. юкатека
- Род † Карибосирен.
- † К. Тернер
- Род † Halitherium.
- † Х. аллени
- † Х. schinzii
- Род † Паралитерий.
- † П. тарканенсе
- Род † Priscosiren. [ 15 ]
- † П. Атлантика
- Род † Сиренавус.
- † Св. Венгерский
- Род † Метакситерий.
- † М. альбифонтанум
- † М. арктодиты
- † М. crataegense
- † М. Флориданум
- † М. Крахулеци
- † М. средний
- † М. серресии
- † М. субапеннины
- Род † Дусисирен.
- † Д. Девана
- † Д. Джордани
- † Д. Рейнхарти
- † Д. такасатенсис
- Род † Hydrodamalis.
- † H. cuestae
- † Х. гигас
Семейство Trichechidae : [ 16 ]
- Род Трихех
- Род † Анотерий.
- Род † Миосирен.
- † М. Канхами
- † М. кубиков
- Род † Потамосирен.
- Род † Рибодон.
†Семейство Protosirenidae : [ 17 ]
- Род † Ашокия.
- Род † Libysiren.
- Род † Протосирен.
- † П. эотен
- † П. фраза
- † П. упомянул
- † П. саттаенсис
- † П. Смита
†Семейство Prorastomidae : [ 18 ]
- Род † Пезосирен.
- † П. двери
- Род † Prorastomus.
| Общее имя | Род | Научное название | Статус | Распределение | Картина |
|---|---|---|---|---|---|
| Вест-Индский ламантин | Трихех (ламантины) | Т. manatus Линней , 1758 г. | ВУ МСОП | 
|

|
| Африканский ламантин | Трихех (ламантины) | Т. senegalensis Link , 1795 г. | ВУ МСОП | 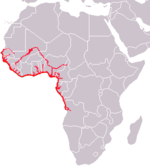
|

|
| Амазонский ламантин | Трихех (ламантины) | Т. inunguis Наттерер , 1883 г. | ВУ МСОП | 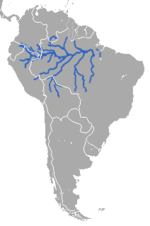
|

|
| Кровь | Кровь | Д. Дюгон Мюллер , 1776 г. | ВУ МСОП | 
|

|
Распределение
Теплые мелководья экватора были центром проживания сирен. [ 19 ] Самая северная из ныне живущих популяций, флоридский подвид вест-индийского ламантина ( T. manatus latirostris ), населяет побережье и часто посещает пресноводные источники, электростанции и каналы во Флориде, чтобы согреться зимой. Особи могут мигрировать на север в теплые летние месяцы, некоторые на расстояние до 1000 километров (около 621,37 миль) от своего зимнего ареала. [ 20 ] Антильский подвид ( T. manatus manatus ) встречается в Карибском бассейне, Южной Америке и Центральной Америке, а также часто встречается на затопленных рифах, в мангровых зарослях, лагунах и зарослях морской травы. [ 21 ]
Амазонский ламантин ( T. inunguis ) был зарегистрирован во всех частях бассейна реки Амазонки в Южной Америке. Соединяющиеся речные каналы позволяют легко добраться до других водных путей, где может быть много еды. Амазонский ламантин обитает только в пресной воде. [ 22 ]
Западноафриканский ламантин ( T. senegalensis ) обитает в мутных изолированных внутренних мангровых зарослях и прибрежных равнинах Западной Африки. Он встречается в водах с температурой выше 18 ° C, а его ареал простирается от Сенегала до Анголы. [ 23 ]
Дюгонь ( Dugong dugong ), ближайший из ныне живущих родственников стеллеровой морской коровы, обитает в Индо-Западной части Тихого океана в более чем 40 различных странах. Это прибрежные животные, обитающие на обширных защищенных лугах с морской травой. [ 24 ]
Стеллерова морская корова была обнаружена в 1741 году вокруг островов в Беринговом море и была приспособлена для холодных субарктических температур. Он простирался от Аляски через Амчитку и Алеутские острова и даже до Японии. [ 25 ] Сообщалось, что стеллерова морская корова собиралась на мелководных песчаных участках вдоль береговой линии и в устьях рек и ручьев, чтобы питаться водорослями. [ 25 ]
Эволюция
[ редактировать ]

Эволюция сирен характеризуется появлением ряда черт, присущих всем сиренам. Ноздри крупные, втянутые , кость верхней челюсти соприкасается с лобной костью , сагиттальный гребень отсутствует, сосцевидный отросток заполняет надвисочное окно (отверстие на верхушке черепа), имеется каплевидная эктотимпанальная (костная кольцо, удерживающее барабанную перепонку ), а кости пахиостеосклеротические (плотные и объемные). [ 9 ]
Сирены впервые появились в летописи окаменелостей в раннем эоцене и разнообразились на протяжении всей эпохи . Они населяли реки, эстуарии и прибрежные морские воды. [ 26 ] Сирены, в отличие от других морских млекопитающих, таких как китообразные , [ 27 ] жил в Новом Свете . В Западной Европе первые и древнейшие останки сирен были найдены на новом палеонтологическом объекте в Санта-Бригиде, Амер (Ла-Сельва, Каталония , Испания). [ 28 ] ). Одним из самых ранних обнаруженных водных сирен является Prorastomus , который появился 40 миллионов лет назад, а первый известный сирен, четвероногий Pezosiren , жил 50 миллионов лет назад. [ 26 ] В Тунисе была найдена древняя окаменелость каменистой кости сирен , датируемая примерно тем же временем, что и Прорастом . [ 29 ] Это самая старая окаменелость сирен, найденная в Африке, и она подтверждает молекулярные данные, предполагающие, что сирены, возможно, произошли из Африки. [ 29 ] Prorastomidae и Protosirenidae , самые ранние семейства сиреновых , состояли из свиноподобных земноводных существ, вымерших в конце эоцена. С появлением в это время Dugongidae у сирен развились характеристики современного отряда, в том числе водное обтекаемое тело с ластообразными передними конечностями и отсутствием задних конечностей, а также мощный хвост с горизонтальными хвостовыми плавниками, поднимающимися вверх. движением вниз, чтобы переместить их через воду. [ 30 ]
Последнее из появившихся семейств сирен, Trichechidae , по-видимому, произошло от ранних дюгонгид в позднем эоцене или начале олигоцена. В 1994 году семейство было расширено за счет включения не только подсемейства Trichechinae ( Potamosiren , Ribodon и Trichechus ), [ 31 ] но также Miosireninae ( Anomotherium и Miosiren ). Африканский ламантин и вест-индийский ламантин более тесно связаны друг с другом, чем с амазонским ламантином. [ 9 ]
Dugongidae включает подсемейства Dugonginae и Hydrodamalinae и парафилетическое Halitheriinae. Бивни современных дюгоней, возможно, изначально использовались для рытья земли, но теперь они используются для социального взаимодействия. Род Дюгонь, вероятно, возник в Индо-Тихоокеанском регионе. [ 9 ]
Описание
[ редактировать ]Адаптации
[ редактировать ]
Хвостовой плавник дюгоня имеет выемку и похож на таковой у дельфинов , тогда как хвостовой плавник ламантина имеет веслообразную форму. [ 9 ] Двуустка накачивается вверх и вниз длинными движениями, чтобы переместить животное вперед, или поворачивается, чтобы повернуть. Передние конечности представляют собой ласты, похожие на весла, которые помогают поворачиваться и замедляться. [ 30 ] [ 32 ] В отличие от ламантинов, у дюгоня на ластах отсутствуют ногти , которые составляют всего 15% длины тела дюгоня. [ 33 ] Ламантины обычно планируют со скоростью 8 километров в час (5 миль в час), но могут достигать скорости 24 километра в час (15 миль в час) короткими очередями. [ 34 ] Тело веретенообразное, что позволяет уменьшить сопротивление воды. Как и у китообразных, задние конечности внутренние и рудиментарные . Морда кормление наклонена вниз, чтобы облегчить со дна . [ 35 ] Сирены обычно совершают двух- или трехминутные погружения. [ 36 ] но ламантины могут задерживать дыхание до 15 минут во время отдыха [ 34 ] и дюгони до шести минут. Они могут стоять на хвосте, чтобы держать голову над водой. [ 37 ]


Как и слоны, ламантины — полифиодонты , постоянно заменяющие зубы на задней части челюсти. У взрослых отсутствуют резцы, клыки и премоляры, вместо них имеется от 8 до 10 щечных зубов . У ламантинов неограниченное количество зубов, появляющихся сзади и выпадающих спереди; они непрерывно образуются зубной капсулой позади зубного ряда. Эти зубы постоянно изнашиваются абразивными сосудистыми растениями, которые они добывают в пищу, особенно водными травами. В отличие от зубов ламантинов, зубы дюгоня не вырастают постоянно за счет горизонтальной замены зубов. [ 38 ] У дюгоня есть два бивня, которые появляются у самцов в период полового созревания, а у самок — позже, после достижения основания предчелюстной кости . [ 33 ] Количество групп слоев роста в бивне указывает на возраст дюгоня. [ 39 ]
У сирен наблюдается пахиостоз – состояние, при котором ребра и другие длинные кости твердые и содержат мало или вообще не содержат костного мозга . У них одни из самых плотных костей в животном мире. Они могут действовать как балласт , противодействуя плавучести их жира и помогая им оставаться в подвешенном состоянии немного ниже поверхности воды. [ 40 ] Ламантины сами по себе не имеют жира, но имеют толстую кожу и, следовательно, чувствительны к изменениям температуры. Они часто мигрируют в более теплые воды, когда температура воды падает ниже 20 ° C (68 ° F). Легкие сирен нелопастные; [ 41 ] Вместе с диафрагмой они простираются на всю длину позвоночного столба, помогая животным контролировать свою плавучесть и уменьшая опрокидывание в воде. [ 42 ] [ 43 ]
Существующие сирены вырастают до 2,5–4 метров (от 8,2 до 13,1 футов) в длину и могут весить до 1500 кг (3300 фунтов). Морская корова Стеллера была самой крупной из когда-либо живших сирен и могла достигать длины 9 метров (30 футов). [ 41 ] и массой от 8 до 10 тонн (от 8,8 до 11,0 коротких тонн). [ 44 ] Мозг дюгоня весит максимум 300 граммов (11 унций), что составляет около 0,1% от веса тела животного. [ 33 ] Тела сирен редко покрыты короткими волосами ( вибриссами ), за исключением того, что они становятся более густыми на морде, что может позволить тактильно интерпретировать окружающую их среду. [ 45 ] Ламантины — единственный известный организм с равномерно васкуляризированной роговицей. Это может быть результатом раздражения или защиты от гипотонической пресноводной среды. [ 46 ]
Диета
[ редактировать ]
Сирен называют «морскими коровами», потому что их рацион состоит в основном из морских водорослей . Дюгони рыщут по морскому дну в поисках морских трав, используя обоняние, поскольку у них плохое зрение. [ 47 ] Они поедают все растение, включая корни. [ 48 ] хотя, если это невозможно, они будут питаться только листьями. [ 39 ] Известно, что западно-индийский ламантин, используя свою разделенную верхнюю губу, потребляет более 60 различных пресноводных и морских растений, таких как мелководье , водяной салат , мускус , ламантиновая трава и черепаховая трава . Взрослый ламантин обычно съедает до 10–15% веса своего тела, или 50 килограммов (110 фунтов), в день, что требует, чтобы ламантин пасся в течение нескольких часов в день. [ 49 ] Напротив, 10% рациона африканского ламантина составляют рыба и моллюски . [ 50 ] Известно, что ламантины поедают небольшое количество рыбы из сетей. [ 51 ]
В отличие от массового кормления , дюгони нацелены на травы с высоким содержанием азота, чтобы максимизировать потребление питательных веществ, и, хотя они преимущественно травоядны , дюгони иногда поедают беспозвоночных, таких как медузы , морские асцидии и моллюски . Некоторые популяции дюгоней, например, в заливе Мортон в Австралии, всеядны и питаются беспозвоночными, такими как полихеты. [ 48 ] или морские водоросли, когда запасы морских трав в них невелики. В других популяциях дюгоней в западной и восточной Австралии есть свидетельства того, что дюгони активно ищут крупных беспозвоночных. [ 39 ]
Популяции амазонских ламантинов ограничиваются озерами в засушливый сезон с июля по август , когда уровень воды начинает падать, и считается, что в этот период они голодают. Их большие запасы жира и низкая скорость метаболизма (всего 36% от обычной скорости метаболизма плацентарных млекопитающих) позволяют им выживать до семи месяцев с небольшим количеством еды или вообще без нее. [ 52 ]
Пищевое поведение
[ редактировать ]Важная анатомия при кормлении
[ редактировать ]Периоральные щетинки используются не только для ощущения предметов, но и для захвата пищи и манипулирования ею. Из шести отдельных полей щетинок на верхней и нижней губе периоральные поля имеют четкое соотношение длины и диаметра, определяющее их границы. Макровибриссы используются для обнаружения пищи по ее размеру, а микровибриссы — для манипулирования ею. Их можно использовать для обламывания листьев и нежелательных частей во время кормления. Сирены используют свою сложную лицевую мускулатуру вместе с периоральной щетиной, чтобы добывать, манипулировать и поглощать водную растительность. Морда представляет собой мышечный гидростат — биологическую структуру, которая опирается на мышечное давление и мышечные сокращения, чтобы манипулировать пищей и перемещать ее. Ламантин использует свои большие верхние периоральные щетинки для выполнения хватательного движения: он выполняет расклешение, которое напрягает мышечный гидростат, в то время как большие верхние щетинки выталкиваются, а нижняя челюсть опускается и, закрываясь, сметает растительность. Основными щетинками, используемыми для поглощения растительности, являются поля U2 и L1. Дюгони и трихехиды различаются тем, как они используют поля щетинок U1 и U2 во время кормления. Дюгони используют медиально-латеральные движения щетинок U2, а трихехиды используют цепкие хватательные движения латерально-медиально. Такое различное пищевое поведение позволяет дюгоням более эффективно, чем трихехидам, использовать донные корма, включая потребление корневищ. [ 53 ]
Обработка пищевых продуктов
[ редактировать ]Обращение с пищей измеряли путем наблюдения за продолжительностью циклических движений (циклов питания) периоральных щетинок ламантинов, используемых для введения пищи в рот. Средняя продолжительность цикла кормления варьировалась в зависимости от размера тела ламантинов и вида потребляемых растений. Скорость введения пищи, рассчитанная на основе средней продолжительности цикла кормления, была сопоставима со скоростью жевания, о которой сообщалось в других исследованиях. Ламантины поедали растения с трубчатыми стеблями и многочисленными ветвями быстрее, чем растения с плоскими пластинками. Время обработки пищи с использованием периоральной щетинки различалось в зависимости от вида потребляемых растений, что отражалось в средней продолжительности цикла кормления. Таким образом, ламантины Флориды адаптируют свое пищевое поведение в зависимости от особенностей потребляемых ими растений. Они демонстрируют разные стратегии и эффективность обработки пищевых продуктов в зависимости от вида растений, при этом более быстрое потребление наблюдается у растений с трубчатыми стеблями и многочисленными ветвями. Это исследование дает ценную информацию об экологии кормления. [ 54 ]
Выращивание Выпас
[ редактировать ]Дюгони ограничены в питании из-за рудиментарного зубного ряда и ограниченного содержания азота в морских травах. Чтобы противостоять этому, они используют стратегию, называемую «выпас с культивированием». Этот выпас может изменить состав сообществ морских водорослей и благоприятствовать видам. Ранние и быстрорастущие виды превзойдут медленнорастущие виды. Часто эти «первопроходческие» виды могут содержать много азота и мало клетчатки, что делает их предпочтительным рационом для дюгоней. Чтобы обеспечить обилие любимых морских трав, дюгони оказывают постоянное давление на участки с водорослями в течение месяца и более. Выпас максимально увеличивает присутствие видов, предпочитаемых дюгонями, за счет менее питательных и менее предпочтительных видов. Этот метод выпаса также способствует быстрому восстановлению лугов с морской травой: дюгони пасутся на извилистых одиночных тропах, оставляющих необрезанные участки морской травы. Этот невыпасаемый заповедник с сохранившимися корневищами является ключом к расширению и восстановлению морских трав. Морские травы реагируют на урожай увеличением уровня азота и уменьшением количества лигнина. Культивирование выпаса позволяет дюгоням увеличить не только питательную ценность морских трав, но и общее питательное качество морских трав. Поддерживая морские травы в незрелом состоянии, дюгони обеспечивают высочайший уровень питания. [ 55 ]
Воспроизведение
[ редактировать ]Несмотря на то, что сирены в основном ведут одиночный образ жизни, они собираются в группы, пока у самок наступает течка . Эти группы обычно включают одну самку и несколько самцов. Сирены являются K -селекторами ; Несмотря на свое долголетие, самки рожают всего несколько раз за свою жизнь и вкладывают значительную родительскую заботу в своих детенышей. Дюгони обычно собираются группами менее дюжины особей на один-два дня. Поскольку они собираются в мутной воде, об их репродуктивном поведении мало что известно. Самцов часто можно увидеть со шрамами, а бивни у дюгоней растут первыми у самцов, что позволяет предположить, что они играют важную роль в токе. Также известно, что они бросались друг на друга. Возраст, когда женщина впервые рожает, является спорным и варьируется от 6 до 17 лет. [ 33 ] Время между родами неясно, по оценкам, от 2 до 7 лет. [ 56 ] [ 39 ] В Сарасоте, штат Флорида, 53 самки под наблюдением произвели не менее 55 детенышей за пятилетний период. [ 57 ]
Ламантины могут достичь половой зрелости уже в возрасте от двух до пяти лет. [ 57 ] ламантинов Беременность длится около года, а затем они лактируют в течение одного-двух лет. Вест-индийские и африканские ламантины могут размножаться круглый год, а самка спаривается с несколькими самцами. [ 58 ] У амазонских ламантинов есть сезон размножения, обычно спаривание происходит, когда уровень реки начинает повышаться, что варьируется от места к месту. [ 59 ]
Ламантины в неволе
[ редактировать ]Ламантины могут быть взяты в неволу после того, как их нашли застрявшими, чтобы облегчить их восстановление. [ 60 ] и есть много случаев успешной реабилитации и выпуска ламантинов в дикую природу. [ 61 ] Поскольку все существующие виды сирен относятся к уязвимым, эти программы реабилитации представляют собой полезный способ поддержки этих видов. Однако уязвимость этих животных также означает, что изъятие ламантинов из дикой природы в коммерческих целях является проблемой сохранения.
Диета в неволе
[ редактировать ]Ламантины, как правило, хорошо себя чувствуют в неволе и, как известно, процветают. [ 62 ] Однако может быть сложно воспроизвести условия их естественной среды в той степени, которая необходима для поддержания здоровья ламантина; Типичный рацион содержащихся в неволе популяций ламантинов может содержать недостаточное количество необходимых им питательных веществ. [ 49 ]
Диета ламантинов, которых кормят в неволе, сильно отличается от диеты ламантинов в дикой природе. В неволе ламантинов кормят 70–80 % зеленых листовых овощей, 10–20 % сушеных кормов и 5 % овощей и фруктов. [ 63 ] Сушеные корма — это такие продукты, как сено и тимофеевка, которые часто используются в качестве корма для лошадей и крупного рогатого скота. Овощи и фрукты, которыми кормят ламантинов, включают салат ромэн, морковь и яблоки. В естественной среде обитания примерно половину рациона ламантинов составляют морские или устьевые растения. [ 64 ] По сравнению с рационом, содержащимся в неволе, водные растения содержат больше сухого вещества и растворимой нейтральной детергентной клетчатки, а также меньше усвояемых питательных веществ. [ 64 ] Хотя может показаться, что более легкоусвояемые питательные вещества представляют собой лучшую диету, желудочно-кишечный тракт ламантина адаптируется к дикой диете посредством микробных процессов ферментации. [ 64 ]
В усилиях по спасению и реабилитации часто участвуют осиротевшие детеныши ламантинов. В неволе молодых ламантинов будут кормить из бутылочки молочной смесью на основе аминокислот, которая включает источник белка, масла и стабилизирующий агент. Эта смесь дополнена витаминами. Во время приема пищи молодым ламантинам могут потребоваться электролиты посредством внутривенной гидратации или даже кормления через зонд, если они постоянно отказываются от бутылочки. [ 65 ] Через шесть месяцев их познакомят с твердой пищей, такой как салат ромэн и салат айсберг, тыква и корнеплоды. Через полтора года начнется процесс отлучения от груди, и молодым ламантинам во время кормления будут предлагать все меньше и меньше молока, постепенно переходя на полностью твердую пищу. [ 65 ]
Угрозы и сохранение
[ редактировать ]
Три современных вида ламантинов (семейство Trichechidae) и дюгонь (семейство Dugongidae) внесены как уязвимые в Красный список угрожаемых видов МСОП . Все четыре вида уязвимы к вымиранию из-за утраты среды обитания и других негативных последствий, связанных с ростом населения и освоением прибрежных зон. [ 50 ] [ 66 ] [ 67 ] [ 68 ] Стеллерова морская корова , вымершая с 1768 года, была истреблена людьми. [ 69 ]
Мясо, масло, кости и кожа ламантинов имеют коммерческую ценность. В некоторых странах, таких как Нигерия и Камерун, африканские ламантины продаются в зоопарках, аквариумах и в Интернете в качестве домашних животных, а иногда и отправляются по всему миру. Хотя охота на них незаконна, отсутствие правопорядка в этих районах допускает браконьерство . Некоторые жители западноафриканских стран, таких как Мали и Чад, верят, что масло африканского ламантина может вылечить такие недуги, как ушные инфекции, ревматизм и кожные заболевания. [ 50 ] Охота является крупнейшим источником смертности амазонских ламантинов, и планов управления этим видом нет, за исключением Колумбии. [ 70 ] Амазонских ламантинов, особенно детенышей, иногда незаконно продают в качестве домашних животных, но существует несколько учреждений, которые заботятся об этих сиротах и спасают их с возможностью выпустить их на волю. [ 66 ] Части тела дюгоней используются в качестве лекарственных средств по всему Индийскому океану. [ 39 ]
Ламантины на Кубе столкнулись с браконьерством, запутыванием и загрязнением окружающей среды. В этом районе находится одна из самых обширных и лучших мест обитания ламантинов на Карибах, но популяция там не смогла процветать. [ 71 ] Существующая информация о ламантинах на Кубе ограничена; это затрудняет распространение информации, что, следовательно, повышает риск браконьерства и запутывания в рыболовных сетях в прибрежных общинах. [ 72 ] Браконьерство на ламантинов стало серьезной проблемой с 1970-х годов, когда первоначально сообщалось, что охота наносит ущерб популяции ламантинов на Кубе. [ 71 ] В 1975 году сообщалось, что популяция ламантинов на Кубе была редкой и сокращалась с угрожающей скоростью из-за загрязнения окружающей среды и охоты. В 1996 году ламантины были помещены под защиту в соответствии с Законом № 164 о рыболовстве. Этот закон предусматривал наказания для тех, кто манипулирует ламантинами, причиняет им вред или ранит их. Охота на ламантинов на Кубе в 1990-х годах, возможно, была результатом экономических трудностей, поскольку ламантины рассматривались как источник белка. [ 71 ] Хотя были предприняты усилия по защите популяции ламантинов на Кубе, они не оказались эффективными, как надеялись те, кто занимается защитой популяции. Многие из этих территорий рассматриваются как парки, существующие только на бумаге, и они не оказывают существенного влияния на общение и охрану. [ 73 ]

Экологические опасности, вызванные деятельностью человека, также подвергают сирен опасности. Сирены, особенно вест-индийские ламантины, сталкиваются с высокой смертностью в результате столкновений с водными транспортными средствами, и около половины всех смертей вест-индийских ламантинов вызваны столкновениями с водными транспортными средствами. Увеличение использования гидроэлектроэнергии и последующее перекрытие рек плотинами увеличивают движение водных путей, что может привести к столкновениям судов, а ламантины могут попасть в судоходные шлюзы . Урбанизация береговой линии таких регионов, как Карибский бассейн , Флорида и Австралия, может привести к сокращению популяций морских водорослей. Луга с водорослями также очень восприимчивы к загрязнению и в настоящее время входят в число наиболее уязвимых экосистем на Земле. [ 74 ] Надежные районы с теплой водой во Флориде, как правило, являются результатом сбросов с электростанций, но новые электростанции с более эффективными системами охлаждения могут нарушить структуру убежищ с теплой водой, а также увеличить спрос на артезианские источники , естественный источник теплой воды для животных. , для использования человеком уменьшается количество убежищ с теплой водой. Собрание в теплых водах промышленных районов Флориды может подвергнуть ламантинов воздействию загрязняющих веществ и токсинов в то время года, когда их иммунная система уже ослаблена. [ 75 ]
Сирен могут поймать в качестве прилова при рыболовстве, а также рассматривать как вредителей, которые мешают местным рыбакам и повреждают их сети. [ 50 ] [ 66 ] [ 67 ] [ 68 ] Известно также, что африканские ламантины проникают на рисовые поля и уничтожают посевы во время сезона дождей, и эти столкновения с местными жителями могут привести к преднамеренному убийству ламантинов. [ 76 ]
Красный прилив — вредоносное цветение водорослей Karenia brevis , выделяющее в воду токсины, убивает многие морские виды. В 1982 году многие ламантины заболели из-за употребления в пищу бреветоксинов , которые накопились в фильтрующих организмах, прикрепленных к стеблям морских водорослей. Ламантины также могут вдыхать эти бревотоксины с поверхности воды, поднимаясь на воздух, что приводит к респираторным симптомам и даже к утоплению. [ 75 ] Гибель ламантинов в результате воздействия токсинов красного прилива была зарегистрирована Комиссией по охране рыбы и дикой природы Флориды на юго-западе Флориды в 2002, 2003, 2005, 2007 и 2013 годах. Цветение красного прилива в 2018 году распространилось от округа Паско до округа Коллиер на западе. побережье Флориды. [ 77 ] По состоянию на январь 2018 года в общей сложности 472 случая смерти ламантинов были вызваны этим красным приливом, а также водными транспортными средствами, холодовым стрессом и другими факторами. [ 78 ]
На ламантинов негативно влияет пластик и другой мусор, попадающий в океан и другие водные пути. Пластик и мусор могут привести к запутыванию, проглатыванию, ампутации или даже смерти ламантина. Когда ламантин проглатывает пластик, об этом часто становится известно только после смерти, когда проводится вскрытие и в желудочно-кишечном тракте ламантина обнаруживаются остатки. Из 40 антильских ламантинов, которые были спасены, реабилитированы и выпущены в рамках программы реинтродукции ламантинов в морском аквариуме Клируотера вдоль побережья Центральной и Южной Америки, у четырех был обнаружен пластик в желудочно-кишечном тракте. Лечение было завершено, и ламантины были освобождены. Позже трое из четырех были найдены мертвыми: двое - в результате проглатывания пластика, а у третьего - с кусочками пластика в желудочно-кишечном тракте. Предметы, обнаруженные в желудочно-кишечном тракте умерших ламантинов, включали презервативы, пластиковые пакеты, полиэстер Raschel , неизвестный пластиковый мусор, а также обертки от мороженого и предметов гигиены. [ 79 ]
В ходе исследования, проведенного в Университете Майами во Флориде, было оценено 439 туш ламантинов, которые были обнаружены и вскрыты в период с 1978 по 1986 год. Из них 63 (14,4%) проглотили остатки; Четверо животных погибли в результате проглатывания пластика или другого мусора. Среди мусора, обнаруженного в желудочно-кишечном тракте животных, были монофильная леска (наиболее распространенный предмет), пластиковые пакеты, веревка, бечевка, веревка, рыболовные крючки, проволока, бумага, целлофан, синтетические губки, резиновые ленты и чулки. [ 80 ]
Инфекционные заболевания также могут играть важную роль в заболеваемости и смертности. Хотя вирусы были идентифицированы только у ламантинов Флориды, паразиты и бактерии наблюдались как минимум у трех из четырех видов сирен. [ 81 ] [ 82 ] [ 83 ] [ 84 ] Вирусы, обнаруженные у ламантинов Флориды, включают трихехидный герпесвирус 1 (TrHV-1) и папилломавирусы ламантина (TmPV) 1–4. [ 85 ] [ 86 ] [ 87 ] Сообщается, что микобактериоз привел к смертности содержащихся в неволе ламантинов Флориды и заболеванию амазонских ламантинов, в то время как такие бактерии, как Vibrio , Pasteurella , Pseudomonas , Streptococcus и Clostridium , были культивированы из мертвых дюгоней в Австралии. [ 88 ] [ 89 ] [ 90 ] Сальмонеллез связан со смертностью дюгоней, по крайней мере, с 1981 года. [ 82 ] Известно, что сенегальский ламантин, хотя и недостаточно изучен, является хозяином нематоды Heterocheilus tunicatus, как и его родственный вид, вест-индийский ламантин. [ 91 ] [ 84 ] Еще многое предстоит узнать об угрозе, которую инфекционные заболевания представляют как для диких, так и для содержащихся в неволе популяций ламантинов. Взаимосвязь между наличием определенных потенциальных патогенов, в том числе перечисленных выше, и их влиянием на заболевание у людей до сих пор в значительной степени неизвестна, хотя установлено, что многие дикие ламантины являются положительными на вирус папилломы без каких-либо известных негативных последствий для здоровья. У людей с ослабленным иммунитетом и положительным результатом теста на вирус папилломы иногда могут развиваться кожные поражения; однако кожный папилломатоз не всегда коррелирует с папилломавирусной инфекцией, и необходимы дальнейшие исследования. [ 92 ]
Во Флориде сельскохозяйственные стоки могут негативно повлиять на среду обитания ламантинов, а во время сезона дождей более 50 округов практикуют запрет на использование удобрений, чтобы попытаться ограничить попадание загрязняющих веществ в водные пути. [ 93 ] Погодные катастрофы и другие природные явления также являются источниками смертности. Вест-Индский ламантин и дюгонь сталкиваются с риском ураганов и циклонов , который, по прогнозам, усилится в будущем. Эти штормы могут распространять загрязняющие вещества [ 94 ] и может нанести ущерб популяциям морских водорослей. [ 68 ] [ 67 ] Африканские ламантины могут оказаться на мели во время засушливого сезона, когда реки и озера становятся слишком маленькими или полностью пересыхают. [ 50 ]
Изменение климата вызывает растущую обеспокоенность у ламантинов, поскольку изменения температуры могут повлиять на уровень моря, pH, осадки, соленость и характер циркуляции прибрежных экосистем. [ 94 ] Прогнозируется, что изменение климата сделает зимние месяцы еще холоднее, что приведет к увеличению случаев холодового стресса у ламантинов. [ 95 ] Известно, что ламантины обладают невероятно низкой скоростью обмена веществ и плохой изоляцией, поэтому им сложнее терморегуляция в условиях холодной воды. Обычно они мигрируют в более теплые воды, как только температура воды начинает падать ниже 20 градусов по Цельсию. Это может включать в себя более теплые воды естественным путем или искусственные среды обитания с теплой водой, созданные электростанциями/выпусками энергетических центров. [ 96 ] Синдром холодового стресса у ламантинов может возникнуть при длительном воздействии воды с температурой ниже 20 градусов по Цельсию, что в конечном итоге может привести к обморожениям кожи, анорексии, жировой атрофии, лимфоидному истощению, а также вторичным инфекциям и заболеваниям. [ 97 ] По данным Комиссии по охране рыбы и дикой природы Флориды, за последние три года было зарегистрировано самое большое на сегодняшний день количество смертей, связанных с простудой. [ 97 ] Это распространенное заболевание, которое лечат в центрах реабилитации ламантинов в штате Флорида, таких как SeaWorld Orlando, зоопарк Тампы в Лоури-парке, океанариум Майами и зоопарк и сады Джексонвилля.
Реабилитация ламантинов от таких заболеваний, как синдром холодового стресса, возможна благодаря поддержке ветеринарного персонала, работников зоопарков, исследователей и добровольцев на местах. Число ламантинов, нуждающихся в вмешательстве, вероятно, увеличится по мере уменьшения количества мест обитания в теплой воде из-за снижения весенних расходов и вывода из эксплуатации электростанций. [ 98 ] Обычные методы лечения синдрома холодового стресса ламантина могут включать теплую чистую воду, антибиотики, регидратацию, клизмы и минеральное масло от запоров и инородных тел, а также, что наиболее важно, пищевые добавки. [ 99 ] Некоторым пациентам с ламантинами, испытывающим поражения, похожие на обморожения, для оптимального восстановления может быть рекомендован длительный прием добавок. [ 99 ] Больше всего от синдрома холодового стресса страдают те, кто недавно был отнят от груди, и лечить их сложнее из-за опасений, связанных с переохлаждением.
Повышение температуры океана может вызвать вредоносное цветение водорослей, которое может заглушить свет, необходимый для роста морских водорослей. [ 100 ] Уменьшение зарослей морских водорослей означает, что все больше ламантинов собираются на меньших территориях для кормления, что усиливает конкуренцию за ресурсы и распространение патогенов. [ 75 ] Воздействие бреветоксина во время красного прилива также является источником смертности; ламантины могут подвергнуться воздействию бреветоксина после утихания красного прилива, поскольку он может накапливаться на морских травах. [ 67 ] Поедание растительности также приводит к образованию отложений, что приводит к попаданию в организм загрязняющих веществ, попавших в грязь. [ 75 ]
Все сирены находятся под защитой Закона США о защите морских млекопитающих 1972 года, Закона США об исчезающих видах 1973 года и Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения ( СИТЕС ). [ 101 ] В дополнение к этому, эти четыре вида дополнительно охраняются различными специализированными организациями. Дюгонь внесен в список Конвенций о биологическом разнообразии , Конвенции о мигрирующих видах и Инициативы кораллового треугольника . [ 68 ] Во Флориде ламантины находятся под защитой Закона о заповедниках ламантинов Флориды 1978 года, который предусматривает такие действия, как запрет на водные транспортные средства или ограничение их скорости там, где существуют ламантины. [ 102 ] Программы реабилитации морских млекопитающих реализуются и регулируются в Соединенных Штатах уже более 40 лет. В 1973 году во Флориде были спасены или оказаны помощь раненым и пострадавшим ламантинам. В конце концов, программа была преобразована в Программу спасения, реабилитации и выпуска ламантинов, управляемую Службой охраны рыбы и дикой природы США. В 2012 году программа стала Партнерством по спасению и реабилитации ламантинов при разрешении и надзоре со стороны USFWS. С 1973 по 2014 год в рамках этой программы было спасено 1619 ламантинов и выпущено на свободу 526. [ 103 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Шошани 2005 .
- ^ Jump up to: а б Уокер, Мэтт. «Гигантская корова, переплывшая океан» . Би-би-си . Проверено 29 октября 2020 г.
- ^ Домнинг, Дэрил П. (1982). «Эволюция ламантинов: умозрительная история» . Журнал палеонтологии . 56 (3): 599–619. ISSN 0022-3360 . JSTOR 1304394 .
- ^ де Соуза, Эрика Мартинья Силва; Фрейтас, Лукас; да Силва Рамос, Элиза Карен; Селлегин-Вейга, Джованна; Рашид-Рибейро, Мишель Карнейро; СИЛЬВА, Фелипе Андре; МАРМОНТЕЛЬ, Мириам; душ Сантос, Фабрисио Родригес; Лаудисуа, Энн; Верхейен, Эрик; Домнинг, Дэрил П. (11 февраля 2021 г.). «Эволюционная история ламантинов, рассказанная их митогеномами» . Научные отчеты . 11 (1): 3564. Бибкод : 2021NatSR..11.3564D . дои : 10.1038/s41598-021-82390-2 . ISSN 2045-2322 . ПМЦ 7878490 . ПМИД 33574363 .
- ^ «Сирения Иллигер, 1811 год» . Интегрированная таксономическая информационная система .
- ^ Что такое сирены? Архивировано 20 июля 2012 г. в Wayback Machine Sirenian International - Исследования, образование и охрана ламантинов и дюгоней.
- ^ Берта 2005 , с. 103 .
- ^ Jump up to: а б Табусе, Р.; Ашер, Р.Дж.; Леманн, Т. (2008). «Афротерийские млекопитающие: обзор современных данных» (PDF) . Млекопитающие . 72 : 2–14. дои : 10.1515/МАММ.2008.004 . S2CID 46133294 . Архивировано из оригинала (PDF) 24 февраля 2021 г. Проверено 2 января 2017 г.
- ^ Jump up to: а б с д и ж Берта 2005 , стр. 89–100 .
- ^ Свартман, М.; Станьон, Р. (2012). «Хромосомы афротерий и их влияние на эволюцию генома млекопитающих» . Цитогенетические и геномные исследования . 137 (2–4): 144–153. дои : 10.1159/000341387 . ПМИД 22868637 . S2CID 24353318 .
- ^ Симпсон, Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85 : 1–350.
- ^ Роуз, Кеннет Д.; Арчибальд, Дж. Дэвид (2005). Возникновение плацентарных млекопитающих: происхождение и взаимоотношения основных существующих клад . Университет Джонса Хопкинса. п. 99. ИСБН 978-0-8018-8022-3 .
- ^ «Классификация семейства Dugongidae» . Ископаемые работы. Архивировано из оригинала 12 декабря 2021 года . Проверено 1 января 2017 г.
- ^ Велес-Хуарбе, Хорхе; Домнинг, Дэрил П. (2015). «Ископаемая Sirenia Западной Атлантики и Карибского региона. XI. Callistosiren boriquensis , gen. et sp. nov». Журнал палеонтологии позвоночных . 35 (1): e885034. Бибкод : 2015JVPal..35E5034V . дои : 10.1080/02724634.2014.885034 . S2CID 84600341 .
- ^ Велес-Хуарбе, Хорхе; Домнинг, Дэрил П. (2014). «Ископаемая сирения Западной Атлантики и Карибского региона. X. Priscosiren atlantica , sp. nov». Журнал палеонтологии позвоночных . 34 (4): 951. doi : 10.1080/02724634.2013.815192 . S2CID 85297028 .
- ^ «Классификация семейства Trichechidae» . Ископаемые работы. Архивировано из оригинала 12 декабря 2021 года . Проверено 1 января 2017 г.
- ^ «Классификация семейства Protosirenidae» . Ископаемые работы. Архивировано из оригинала 12 декабря 2021 года . Проверено 1 января 2017 г.
- ^ «Классификация семейства Prorastomidae» . Ископаемые работы. Архивировано из оригинала 12 декабря 2021 года . Проверено 1 января 2017 г.
- ^ Кнудсен., Бонде, Роберт. Популяционная генетика и сохранение флоридского ламантина: прошлое, настоящее и будущее . OCLC 704839872 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Амазонский ламантин» . ГРАНЬ Существования . Архивировано из оригинала 07 февраля 2022 г. Проверено 25 июля 2022 г.
- ^ ЛаКоммар, Кэтрин С.; СамоСалливан, Кэрин; Бро, Соланж (1 января 2008 г.). «Распространение и использование среды обитания антильских ламантинов (<I>Trichechus manatus manatus</I>) в районе затонувших островов Белиза, Центральная Америка» . Водные млекопитающие . 34 (1): 35–43. дои : 10.1578/am.34.1.2008.35 . ISSN 0167-5427 .
- ^ де Соуза, Диого Александр; Гонсалвес, Андре Луис Соуза (июль – сентябрь 2021 г.). «Оценка численности и вероятности обнаружения амазонского ламантина (Trichechus inunguis) в Центральной Амазонии, Бразилия» . Перспективы экологии и охраны природы . 19 (3): 354–361. Бибкод : 2021PEcoC..19..354D . дои : 10.1016/j.pecon.2021.03.009 .
- ^ «Западноафриканский ламантин | млекопитающее | Британика» . www.britanica.com . Проверено 21 февраля 2024 г.
- ^ «Дюгонь-дюгон - Общество морской млекопитающей» . www.marinemammalscience.org . Проверено 25 июля 2022 г.
- ^ Jump up to: а б (США), Геологическая служба. Стеллерова морская корова (Hydrodamalis gigas) позднеплейстоценового возраста с Амчитки, Алеутские острова, Аляска . OCLC 148863032 .
- ^ Jump up to: а б Домнинг, Д.П. (2001). «Самый ранний известный полностью четвероногий сирен». Природа . 413 (6856): 625–627. Бибкод : 2001Natur.413..625D . дои : 10.1038/35098072 . ПМИД 11675784 . S2CID 22005691 .
- ^ Тьювиссен, JGM; Баджпай, Сунил (2001). «Происхождение китов как образец макроэволюции» . Бионаука . 51 (12): 1037–1049. doi : 10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2 .
- ^ Фуэнтес-Буксо, Р., Фуэнтес-Буксо, А. 2016. Обнаружение останков сирен в лютетинском периоде (средний эоцен) Санта-Бригиды (Амер, Ла-Сельва, Жирона) . Treballs del Museu de Geologia de Barcelona, 22: 19-24.
- ^ Jump up to: а б Бенуа, Жюльен; Аднетт, Сильвен; Мабрук, Эссид; Хаяти, Хайет; Бен Хадж Али, Мустафа; Мариво, Лоран; Мерзеро, Жиль; Мериго, Самуэль; Виани-Лио, Моник (16 января 2013 г.). «Остатки черепов из Туниса дают новые сведения о происхождении и эволюции Sirenia (Mammalia, Afrotheria) в Африке » ПЛОС ОДИН 8 (1): e54307. Бибкод : 2013PLoSO... 854307B дои : 10.1371/journal.pone.0054307 . ISSN 1932-6203 . ПМЦ 3546994 . ПМИД 23342128 .
- ^ Jump up to: а б Берта, Аннализ (2012). «Разнообразие, эволюция и адаптация сирен и других морских млекопитающих» . Возвращение в море: жизнь и эпоха эволюции морских млекопитающих . Беркли, Калифорния: Калифорнийский университет. п. 127. ИСБН 978-0-520-27057-2 .
- ^ «Тричехины» . Ископаемые работы . Вход в базу данных палеобиологии . Проверено 28 июля 2017 г.
- ^ Берта 2005 , с. 250.
- ^ Jump up to: а б с д Марш, Хелен (1987). «Глава 57: Дюгонгиды» (PDF) . Фауна Австралии . Том. 1Б. ЦСИРО. ISBN 978-0-644-06056-1 . Архивировано из оригинала 11 мая 2013 г.
{{cite book}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Jump up to: а б «Ламантин» . Нэшнл Географик . Архивировано из оригинала 14 января 2010 года . Проверено 16 января 2017 г.
- ^ Фельдхамер, Джорджия; Дрикамер, LC; Весси, Ш.; Мерритт, Дж. Ф.; Краевски, Кэри (2015). Маммология: адаптация, разнообразие, экология (4-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. стр. 402–418. ISBN 978-1-4214-1588-8 .
- ^ Чилверс, Б. Луиза ; Делин, С.; Гейлс, Нью-Джерси; Холли, Дания; Лоулер, ИК; Марш, Х.; Прин, Арканзас (2004). «Ныряющее поведение дюгоней, дюгонь-дюгон ». Журнал экспериментальной морской биологии и экологии . 304 (2): 203. Бибкод : 2004JEMBE.304..203L . дои : 10.1016/j.jembe.2003.12.010 .
- ^ «Дюгонь» . Нэшнл Географик . Архивировано из оригинала 12 января 2010 года . Проверено 16 января 2017 г.
- ^ Селф-Салливан, Кэрин, Эволюция Сирении (PDF) , sirenian.org, заархивировано из оригинала (PDF) 31 декабря 2006 г. , получено 10 марта 2007 г.
- ^ Jump up to: а б с д и Марш, Х.; Эрос, К.; Хьюз, Дж. (2002). Дюгонь: отчеты о состоянии и планы действий для стран и территорий (PDF) . Международный союз охраны природы. п. 7. ISBN 978-92-807-2130-0 .
- ^ Уоллер, Джеффри; Дандо, Марк (1996). Sealife: Полное руководство по морской среде . Смитсоновский институт. стр. 413–420 . ISBN 978-1-56098-633-1 .
- ^ Jump up to: а б Элдридж, Нил (2002). Жизнь на Земле: Энциклопедия биоразнообразия, экологии и эволюции . АВС-КЛИО. стр. 532 . ISBN 978-1-57607-286-8 .
- ^ Домнинг, Дэрил; Вивиан Буффренил (1991). «Гидростаз в Сирении: количественные данные и функциональные интерпретации». Наука о морских млекопитающих . 7 (4): 331–368. Бибкод : 1991MMamS...7..331D . дои : 10.1111/j.1748-7692.1991.tb00111.x .
- ^ Роммель, Сентиэль; Джон Э. Рейнольдс (2000). «Структура и функции диафрагмы ламантина Флориды ( Trichechus manatus latirostris )» . Анатомическая запись . 259 (1). Wiley-Liss, Inc.: 41–51. doi : 10.1002/(SICI)1097-0185(20000501)259:1<41::AID-AR5>3.0.CO;2-Q . ПМИД 10760742 .
- ^ Шеффер, Виктор Б. (ноябрь 1972 г.). «Вес стеллеровой коровы». Журнал маммологии . 53 (4): 912–914. дои : 10.2307/1379236 . JSTOR 1379236 .
- ^ Рип, РЛ; Маршалл, CD; Столл, МЛ (2002). «Тактильные волосы на посткраниальном теле ламантинов Флориды: боковая линия млекопитающих?» (PDF) . Мозг, поведение и эволюция . 59 (3): 141–154. дои : 10.1159/000064161 . ПМИД 12119533 . S2CID 17392274 . Архивировано из оригинала (PDF) 11 января 2012 года.
- ^ Амбати, Британская Колумбия; Нодзаки, М.; Сингх, Н.; Такеда, А.; Яни, доктор медицинских наук; Сутар, Т.; Альбукерке, RJC; Рихтер, Э.; Сакурай, Э.; Ньюкомб, Монтана; Кляйнман, Мэн; Колдуэлл, РБ; Лин, К.; Огура, Ю.; Ореккья, А. ; Самуэльсон, Д.А.; Агнью, Вашингтон; Сент-Леже, Дж.; Грин, WR; Махасрешти, П.Дж.; Кюриэль, DT ; Кван, Д.; Марш, Х.; Икеда, С.; Лейпер, LJ; Коллинсон, Дж. М.; Богданович С. ; Хурана, Т.С.; Сибуя, М.; Болдуин, Мэн (2006). «Аваскуляризация роговицы из-за растворимого рецептора VEGF-1» . Природа 443 (7114): 993–997. Бибкод : 2006Nature.443..993A . дои : 10.1038/nature05249 . ПМК 2656128 . ПМИД 17051153 .
- ^ «Дюгонь дюгонь Дюгонь». Млекопитающие Африки: вводные главы и афротерия . 2013. дои : 10.5040/9781472926913.0013 . ISBN 978-1-4729-2691-3 .
- ^ Jump up to: а б Берта 2005 , стр. 438–444.
- ^ Jump up to: а б Сигал-Уиллотт, Джессика Л.; Харр, Кендал; Хайек, Ли-Энн С .; Скотт, Карен С.; Герлах, Тревор; Сируа, Поль; Рейтер, Майк; Крюз, Дэвид В.; Хилл, Ричард К. (2010). «Приблизительный анализ питательных веществ четырех видов подводной водной растительности, потребляемых флоридским ламантином ( Trichechus manatus latirostris ), по сравнению с салатом ромэн ( Lactuca sativa var. longifolia )». Журнал медицины зоопарков и дикой природы . 41 (4): 594–602. дои : 10.1638/2009-0118.1 . JSTOR 40962301 . ПМИД 21370638 . S2CID 4884633 .
- ^ Jump up to: а б с д и Кейт Диагне, Л. (2016) [версия оценки 2015 года с опечатками]. « Трихехус сенегальский » . Красный список исчезающих видов МСОП . 2015 : e.T22104A97168578. doi : 10.2305/IUCN.UK.2015-4.RLTS.T22104A81904980.en . Проверено 11 августа 2021 г.
- ^ Пауэлл, Джеймс (1978). «Свидетельства плотоядности ламантина ( Trichechus manatus )». Журнал маммологии . 59 (2): 442. дои : 10.2307/1379938 . JSTOR 1379938 .
- ^ Бест, Робин С. (1983). «Очевидное голодание в сухой сезон у амазонских ламантинов (млекопитающие: Sirenia)». Биотропика . 15 (1): 61–64. Бибкод : 1983Биотр..15...61Б . дои : 10.2307/2388000 . JSTOR 2388000 .
- ^ Маршалл, Кристофер Д.; Маэда, Хироши; Ивата, Мацумицу; Фурута, Масами; Асано, Сиро; Розес, Фердинанд; Рип, Роджер Л. (28 февраля 2006 г.). «Орофациальная морфология и пищевое поведение дюгоней, амазонских, западноафриканских и антильских ламантинов (Mammalia: Sirenia): функциональная морфология мышечно-вибриссального комплекса» Журнал зоологии 259 (3):245 дои : 10.1017/S0952836902003205 .
- ^ Маршалл, Кристофер; Кубилис, Пол; Хут, Гленн; Эдмондс, Вирджиния; Халин, Дебора; Рип, Роджер (август 2000 г.). «Способность к переработке пищи и продолжительность цикла питания ламантинов, питающихся несколькими видами водных растений». Журнал маммологии . 81 (3): 649. doi : 10.1093/jmammal/81.3.649 .
- ^ А, Прин (10 августа 1995 г.). «Воздействие кормления дюгоня на среду обитания морских водорослей: наблюдательные и экспериментальные данные для выращивания выпаса» . Серия «Прогресс в области морской экологии» . 124 : 201–213. Бибкод : 1995MEPS..124..201P . дои : 10.3354/meps124201 . ISSN 0171-8630 .
- ^ Берта 2005 , с. 502 .
- ^ Jump up to: а б Кельш, Дж. К. (2001). «Размножение самок ламантинов наблюдалось в заливе Сарасота, Флорида». Наука о морских млекопитающих . 17 (2): 331–342. Бибкод : 2001MMamS..17..331K . дои : 10.1111/j.1748-7692.2001.tb01274.x .
- ^ Бест, Робин (1984). Макдональд, Д. (ред.). Энциклопедия млекопитающих . Нью-Йорк: факты в архиве. стр. 292–298 . ISBN 978-0-87196-871-5 .
- ^ Бест, RC (1982). «Сезонное размножение амазонских ламантинов Trichechus inunguis (Mammalia: Sirenia)». Биотропика . 14 (1): 76–78. Бибкод : 1982Биотр..14...76Б . дои : 10.2307/2387764 . JSTOR 2387764 .
- ^ Родственник; Лейте, Кристиан; Эйнхардт Вергара-Кинд, Джосиери; Лима, РП (2004). «Выбросы антильских ламантинов Trichechus manatus manatus на северо-востоке Бразилии» (PDF) . Латиноамериканский журнал водных млекопитающих . 3 (1): 69–75. дои : 10.5597/запись00050 . [ мертвая ссылка ]
- ^ Минуччи-Джаннони, Антонио А. «Плен морских млекопитающих на северо-востоке Карибского моря, с заметками о реабилитации выброшенных на мель китов, дельфинов и ламантинов». Карибский научный журнал 34.3–4 (1998): 191–203.
- ^ Боссарт, Грегори Д. «Ламантины». Справочник CRC по медицине морских млекопитающих . CRC Press, 2001. 989–1010.
- ^ Ларкин, Исканде Л.В.; Фаулер, Вивьен Ф.; Рип, Роджер Л. (6 августа 2007 г.). «Скорость прохождения Digesta у флоридского ламантина (Trichechus manatus latirostris)». Зоопарковая биология . 26 (6): 503–515. дои : 10.1002/zoo.20150 . ПМИД 19360597 .
- ^ Jump up to: а б с Харшоу, Т. Лорен. Оценка питания ламантинов Флориды (Trichechus manatus latirostris). Университет Флориды. Август 2012 г. https://ufdcimages.uflib.ufl.edu/UF/E0/04/45/49/00001/HARSHAW_L.pdf.
- ^ Jump up to: а б Адимей, Н. (2012). Усилия по спасению, реабилитации и выпуску ламантинов как инструмент сохранения видов (PDF) . Сохранение сирен: проблемы и стратегии в развивающихся странах.
- ^ Jump up to: а б с Мармонтель, М.; де Соуза Д. и Кендалл С. (2016). « Трихекус inunguis » . Красный список исчезающих видов МСОП . 2016 : e.T22102A43793736. doi : 10.2305/IUCN.UK.2016-2.RLTS.T22102A43793736.en .
- ^ Jump up to: а б с д Дойч, CJ; Селф-Салливан, К. и Минуччи-Джаннони, А. (2008). « Трихехус Манатус » . Красный список исчезающих видов МСОП . 2008 : e.T22103A9356917. doi : 10.2305/IUCN.UK.2008.RLTS.T22103A9356917.en .
- ^ Jump up to: а б с д Марш, Х. и Собцик, С. (2015). « Дюгонь-дюгон » . Красный список исчезающих видов МСОП . 2015 : e.T6909A43792211. doi : 10.2305/IUCN.UK.2015-4.RLTS.T6909A43792211.en .
- ^ Домнинг, Д. (2016). « Гидродамалис гигас » . Красный список исчезающих видов МСОП . 2016 : e.T10303A43792683. doi : 10.2305/IUCN.UK.2016-2.RLTS.T10303A43792683.en . Проверено 11 августа 2021 г.
- ^ Ривз, Рэндалл Р.; Лезервуд, Стивен; Джефферсон, Томас А.; Карри, Барбара Э.; Хеннингсен, Томас (1996). «Амазонские ламантины, Tricheus inunguis , в Перу: распространение, эксплуатация и статус охраны» (PDF) . Интерсиенсия . 21 (6).
- ^ Jump up to: а б с Альварес-Герман, Анмари; Гарсиа Альфонсо, Эдди; Форнер Мартин-Вианна, Янет; Эрнандес Гонсалес, Заимиури; Лестница Доменека, Раиса; Уртадо, Эндрю; Пауэлл, Джеймс; Джейкоби, Чарльз А; Фрейзер, Томас К. (2017). «Статус и сохранение ламантинов на Кубе: исторические наблюдения и недавние открытия » Бюллетень морской науки . 94 (2): 313–327. дои : 10.5343/bms.2016.1132 . ISSN 0007-4977 .
- ^ «Сохранение Антильских ламантинов на Кубе | Альянс от моря до берега» . Альянс от моря до берега . Архивировано из оригинала 23 июля 2018 г. Проверено 22 июля 2018 г.
- ^ Альварес-Герман, Анмари; Англ-Вальдес, Джордж А.; Альфонсо, Эдди Гарсия; Пауэлл, Джеймс А.; Тейлор, Синтия Р. (апрель 2017 г.). «Обнаружение находящегося под угрозой исчезновения антильского ламантина Trichechus manatus manatus на охраняемой морской территории острова Молодежи, Куба» . Орикс 51 (2): 324–331. дои : 10.1017/S0030605315001143 . ISSN 0030-6053 .
- ^ Резек, Райан Дж.; Фурман, Брэдли Т.; Юнг, Робин П.; Холл, Маргарет О.; Белл, Сьюзан С. (29 октября 2019 г.). «Долгосрочная реализация проектов по восстановлению морских водорослей во Флориде, США» . Научные отчеты . 9 (1): 15514. Бибкод : 2019НатСР...915514Р . дои : 10.1038/s41598-019-51856-9 . ISSN 2045-2322 . ПМК 6820728 . ПМИД 31664068 .
- ^ Jump up to: а б с д Бонде, Роберт К.; Агирре, А.Алонсо; Пауэлл, Джеймс (13 мая 2004 г.). «Ламантины как стражи здоровья морской экосистемы: они — канарейки весом 2000 фунтов?» . ЭкоЗдоровье . 1 (3). дои : 10.1007/s10393-004-0095-5 . ISSN 1612-9202 . S2CID 20044389 .
- ^ «Африканский ламантин ( Trichechus senegalensis )» . Дикий экран. Архивировано из оригинала 30 ноября 2011 г. Проверено 24 января 2017 г.
- ^ Служба охраны рыбы и дикой природы США. «Текущее состояние Красного прилива» . Архивировано из оригинала 11 декабря 2018 года . Проверено 12 января 2023 г.
- ^ «Смертность ламантинов в 2018 году». Факты о мечехвостах, myfwc.com/research/manatee/rescue-mortality-response/mortality-statistics/2018/
- ^ Бек, Кэти А.; Баррос, Нелио Б. (октябрь 1991 г.). «Воздействие обломков на ламантин Флориды» . Бюллетень о загрязнении морской среды . 22 (10): 508–510. Бибкод : 1991MarPB..22..508B . дои : 10.1016/0025-326x(91)90406-i . ISSN 0025-326X .
- ^ Аттадемо, Фернанда Лоффлер Нимейер; Баленсифер, Дейзи Кристиан; Фрейре, Аугусто Карлуш да Боавиажем; де Соуза, Главсия Перейра; да Кунья, Фабио Адонис Гувейя Карнейро; Луна, Фабия де Оливейра (декабрь 2015 г.). «Антильский ламантин, проглатывающий мусор (Trichechus manatus manatus)» . Бюллетень о загрязнении морской среды . 101 (1): 284–287. Бибкод : 2015MarPB.101..284A . дои : 10.1016/j.marpolbul.2015.09.040 . ISSN 0025-326X . ПМИД 26431561 .
- ^ Аттадемо, Фернанда Лоффлер Нимейер; ОЛИВЕЙРА, Радан Элвис Матиас де; Де Соуза, Главсия Перейра; Луна, Фабия Оливейра (22 ноября 2020 г.). «Инфекционные и неинфекционные заболевания ламантинов в Бразилии» . Acta Scientiae Veterinariae . 48 . дои : 10.22456/1679-9216.107571 . ISSN 1679-9216 .
- ^ Jump up to: а б Эллиотт, Хизер; Томас, Аннетт; Лэддс, П.В.; Хейнсон, GE (1981). «Смертельный случай сальмонеллеза у дюгоня» . Журнал болезней дикой природы . 17 (2): 203–208. дои : 10.7589/0090-3558-17.2.203 . ISSN 0090-3558 . ПМИД 7241705 . S2CID 44490155 .
- ^ Люкот, Морган; Теллез, Мариса; Виверос, Диана (2020). «Отчет о гельминтах-паразитах вскрытого антильского ламантина (Trichechus manatus manatus) в Белизе» . Ветеринарная паразитология: региональные исследования и отчеты . 21 : 100446. doi : 10.1016/j.vprsr.2020.100446 . ПМИД 32862905 . S2CID 221383880 .
- ^ Jump up to: а б Срент, JFA (1983). «Аскаридоидные нематоды сирен — новый вид сенегальского ламантина» . Журнал гельминтологии . 57 (1): 69–76. дои : 10.1017/S0022149X00007902 . ISSN 0022-149X . S2CID 86097133 .
- ^ Боссарт, Грегори Д.; Юинг, Рут Ю.; Лоу, Марк; Пот, Марк; Декер, Сьюзен Дж.; Уолш, Кэтрин Дж.; Гим, Синдже; Дженсон, А.Беннетт (2002). «Вирусный папилломатоз у ламантинов Флориды (Trichechus manatus latirostris)» . Экспериментальная и молекулярная патология . 72 (1): 37–48. дои : 10.1006/exmp.2001.2407 . ПМИД 11784121 .
- ^ Ферранте, Джейсон А.; Кортес-Инохоса, Галактика; Арчер, Линда Л.; Веллехан, Джеймс FX (2017). «Разработка количественного ПЦР-анализа для измерения нагрузки трихехидного герпесвируса 1 у ламантина Флориды (Trichechus manatus latirostris)» . Журнал ветеринарных диагностических исследований . 29 (4): 476–482. дои : 10.1177/1040638717707554 . ISSN 1040-6387 . ПМИД 28467282 .
- ^ Захин, Марьям; Гим, Синдже; Ханал, Суджита; Боссарт, Грегори Д.; Дженсон, Альфред Б.; Джо, Чжунхо (01 декабря 2015 г.). «Молекулярная характеристика новых мукозотропных папилломавирусов флоридского ламантина (Trichechus manatus latirostris)» . Журнал общей вирусологии . 96 (12): 3545–3553. дои : 10.1099/jgv.0.000293 . ISSN 0022-1317 . ПМИД 26395390 .
- ^ Нильсен, Кристен А.; Оуэн, Хелен С.; Миллс, Пол С.; Флинт, Марк; Гибсон, Жюстин С. (2013). «БАКТЕРИИ, ВЫДЕЛЕННЫЕ ОТ ДЮГОНЕЙ (ДЮГОНЬ-ДЮГОН), ПРЕДСТАВЛЕННЫЕ НА ПОСМЕРТНОЕ ИССЛЕДОВАНИЕ В КВИНСЛЕНДЕ, АВСТРАЛИЯ, 2000–2011 гг.» . Журнал медицины зоопарков и дикой природы . 44 (1): 35–41. дои : 10.1638/1042-7260-44.1.35 . ISSN 1042-7260 . ПМИД 23505701 . S2CID 36939237 .
- ^ Рейсфельд, Л; Икута, Сай; Ипполито, Л; Сильватти, Б; Чистый Кузнец, Дж.С.; Кэт-Дэй, младший; Роуз, Fcw; Д'Аффонсека Нет, Джа; да Силва, Vmf (05 марта 2018 г.). «Кожный микобактериоз у амазонского ламантина Trichechus inunguis, содержащегося в неволе» . Болезни водных организмов 127 (3): 231–236. дои : 10.3354/dao03196 . ISSN 0177-5103 . ПМИД 29516862 .
- ^ Сато, Т.; Сибуя, Х.; Охба, С.; Нодзири, Т.; Шираи, В. (2003). «Микобактериоз у двух содержащихся в неволе ламантинов Флориды (Trichechus Manatus Latirostris)» . Журнал медицины зоопарков и дикой природы . 34 (2): 184–188. doi : 10.1638/1042-7260(2003)034[0184:MITCFM]2.0.CO;2 . ISSN 1042-7260 . ПМИД 12885137 . S2CID 1436226 .
- ^ Хатсон, Кейт Сюзанна; Воан, Дэвид Брендан; Блэр, Дэвид (2019). «Первая запись о «рыбьем» кровяном сосальщике (Digenea: Aporocotylidae) у морского млекопитающего: Cardicola dhangali n. sp» . Международный журнал паразитологии: Паразиты и дикая природа . 10 : 23–28. Бибкод : 2019IJPPW..10...23H . дои : 10.1016/j.ijppaw.2019.06.009 . ПМК 6624246 . ПМИД 31334029 .
- ^ Боссарт, Грегори Д.; Юинг, Рут Ю.; Лоу, Марк; Пой, Марк; Декер, Сьюзен Дж.; Уолш, Кэтрин Дж.; Гим, Синдже; Дженсон, А.Беннетт (февраль 2002 г.). «Вирусный папилломатоз у ламантинов Флориды (Trichechus manatus latirostris)» . Экспериментальная и молекулярная патология . 72 (1): 37–48. дои : 10.1006/exmp.2001.2407 . ISSN 0014-4800 . ПМИД 11784121 .
- ^ Киркпатрик, Барбара; Колер, Кейт; Бирн, Маргарет; Флеминг, Лора Э.; Шеллер, Карен; Райх, Эндрю; Хичкок, Гэри; Киркпатрик, Гэри; Ульманн, Стивен; Хоугланд, Портер (сентябрь 2014 г.). «Реакция человека на красные приливы во Флориде: осведомленность о политике и соблюдение местных постановлений об удобрениях» . Наука об общей окружающей среде . 493 : 898–909. Бибкод : 2014ScTEn.493..898K . doi : 10.1016/j.scitotenv.2014.06.083 . hdl : 1912/6871 . ISSN 0048-9697 . ПМИД 25003583 .
- ^ Jump up to: а б Эдвардс, Холли Х. (2 октября 2013 г.). «Потенциальное воздействие изменения климата на тепловодную мегафауну: пример ламантина Флориды (Trichechus manatus latirostris)» . Климатические изменения . 121 (4): 727–738. Бибкод : 2013ClCh..121..727E . дои : 10.1007/s10584-013-0921-2 . hdl : 10.1007/s10584-013-0921-2 . ISSN 0165-0009 . S2CID 154816079 .
- ^ Рунге, Майкл С.; Лангтимм, Кэтрин А.; Мартин, Жюльен; Фоннесбек, Кристофер Дж. (2015). «Анализ статуса и угроз для ламантина Флориды ( Trichechus manatus latirostris ), 2012» . Отчет об открытом файле . дои : 10.3133/ofr20151083 . ISSN 2331-1258 .
- ^ «Среда обитания ламантинов» . Комиссия Флориды по охране рыбы и дикой природы . Проверено 7 августа 2023 г.
- ^ Jump up to: а б Бейли1, Джеймс Э.; Фланаган2, Карла; Миган1, Дженни; Ле-Берт1, Каролина; Джонсон1, Шон; Гомес1, Форрест; Лутмердинг1, Бетси; Смит1, Синтия; Дженсен3, Эрик; Сильва2, Нуно; Сильва2, Жоана; Колитц4, Кармен; Латимер4, Федерико Г.; Нуньес2, Антоньета; Сильвейра2, Мигель (30 марта 2015 г.). «МАААМ 2012» . ВИН.com .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Харди, Стейси К.; Дойч, Чарльз Дж.; Кросс, Тиффани А.; де Вит, Мартина; Хостетлер, Джеффри А. (21 ноября 2019 г.). Хиренбах, Дэвид (ред.). «Смертность ламантинов Флориды, связанная с холодом, в зависимости от температуры воздуха и воды» . ПЛОС ОДИН . 14 (11): e0225048. Бибкод : 2019PLoSO..1425048H . дои : 10.1371/journal.pone.0225048 . ISSN 1932-6203 . ПМК 6871784 . ПМИД 31751355 .
- ^ Jump up to: а б Колдуэлл, Мельба К.; Колдуэлл, Дэвид К.; БК Таунсенд-младший (30 марта 2015 г.). «Архив МАААМ» . ВИН.com .
- ^ Аллен, Аарин Конрад; Бек, Кэти А.; Саттельбергер, Даниэль С.; Кишка, Джереми Дж. (май 2022 г.). «Свидетельства изменения рациона ламантина из Флориды (Trichechus manatus latirostris) в лагуне реки Индиан, полученные на основе анализа содержимого желудка» . Устьевые, прибрежные и шельфовые науки . 268 : 107788. Бибкод : 2022ECSS..26807788A . дои : 10.1016/j.ecss.2022.107788 . ISSN 0272-7714 . S2CID 247048024 .
- ^ «Морские млекопитающие» . Служба охраны рыбы и дикой природы США. Архивировано из оригинала 5 января 2017 года . Проверено 26 января 2017 г.
- ^ «Статуты Флориды 2016 года» . Онлайн Солнце . Проверено 26 января 2017 г.
- ^ Спасение, реабилитация и выпуск ламантинов Флориды: анализ факторов, влияющих на выживание. Автор: Адимей, Нью-Мексико; Раушенбергер, Х.; Рид, JP; и др. P 18 BIENN C BIOL MA Опубликовано: декабрь 2009 г.
Дальнейшее чтение
[ редактировать ]- Дэрил П. Домнинг. «Библиография и указатель Сирений и Демостилий» . Архивировано из оригинала 3 ноября 2013 г.
- Шошани, Дж. (2005). Уилсон, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. стр. 92–93. ISBN 978-0-8018-8221-0 . OCLC 62265494 .
- Берта, А.; Сумич, Дж.Л.; Ковач, К.М. (2015). Морские млекопитающие: эволюционная биология (3-е изд.). Академическая пресса. ISBN 978-0-12-397002-2 .
- Гаррисон, Том. Океанография , 5-е изд., Брукс Коул, 30 июля 2008 г. ISBN 978-0-495-55531-5
Внешние ссылки
[ редактировать ]

Сирениан генерирует |
|---|
