Чашечка Хед

Чашечка Help является особенно большим возбуждающим синапсом в млекопитающих слуховой нервной системе , названная так в честь Ханса, который впервые описал его в своей статье 1893 года Die Centrale Gehörleitung [ 1 ] [ 2 ] Из -за его сходства с чашечкой цветка. [ 3 ] Глобулярные густые клетки в антеровентральном кохлеарном ядре (AVCN) [ 4 ] Отправляйте аксоны в контралатеральное медиальное ядро трапеции ( MNTB), где они синаплят через эти Calyces на основных клетках MNTB. [ 5 ] [ 6 ] [ 7 ] Эти основные клетки затем проецируют в ипсилатеральную боковую верхнюю оливковую (LSO) , [ 8 ] где они ингибируют постсинаптические нейроны и обеспечивают основу для обнаружения межполучия (ILD), необходимого для высокочастотной локализации. [ 9 ] Этот синапс был описан как самый большой в мозге. [ 10 ]
Связанная конечная галочка удерживающего также представляет собой больший аксонный терминальный синапс (диаметр 15–30 мкм), обнаруженный в другой слуховой структуре ствола мозга, а именно в антевентральном кохлеарном ядре (AVCN). [ 11 ] Как и в случае с Calyces, эти синапсы способствуют быстрой, эффективной передаче информации.
Чашечка удерживаемой и конечной удерживающей везикул, содержащих глутамат на пресинаптическом терминале; Везикулы высвобождаются при стимуляции (происходящие в Cochlea и AVCN). Затем глутамат связывается с двумя известными глутаматными рецепторами, AMPA- и рецепторами NMDA , быстро инициируя потенциалы действия в постсинаптической клетке. [ 12 ]
Обычно используется в исследованиях из -за его большого размера, чашечка удерживалась для понимания различных механизмов, связанных с развитием и высвобождением пузырьков синапса.
Функция
[ редактировать ]Чашечка удержания является частью слуховой системы , соединяющей глобулярные густые клетки (GBC) переднеровентрального кохлеарного ядра с основными нейронами медиального ядра трапециевидного тела (MNTB). В качестве синапса функция чашечки удерживается для передачи сигнала из GBC на основные нейроны MNTB, которые являются глицинергическими, таким образом, гиперполяризующие клетки в латеральных верхних оливковых (LSO) ядрах и продуцирующие ингибирующие эффекты. [ 12 ] В результате его роли в стимулировании основных нейронов MNTB, первичная функция чашечки удерживается, чтобы позволить дифференцировку между временной активацией кохлеарных волосковых клеток, которые важны для локализации звука (обнаружение межоверного уровня). [ 13 ]
Обнаружение межоверного уровня возможно через систему чашечки из -за большого относительного размера GBC, чашечки удерживаемой и основных нейронов MNTB. Нейроны в LSO особенно важны для проницательности этих различий в межоверных уровнях. Большой диаметр густых клеточных аксонов позволяет ингибирующему сигналу, полученному нейронами MNTB, достигает SOC примерно на 0,2 мс позже, чем ипсилатеральное возбуждение. Эта разница в ~ 0,2 миллисекундной задержке достаточно короткая, чтобы позволить сравнить уровни с двух сторон для оценки ILD, особенно на высоких частотах, а также позволяет некоторую чувствительность ITD, на частотах достаточно низкие, чтобы иметь велосипедную фазовую блокировку. [ 12 ]
Структура
[ редактировать ]
Для каждого основного нейрона MNTB есть одна чашечка, и для большинства аксонов GBC есть только одна чашечка, хотя есть исключения из этого спаривания. [ 1 ] Это в целом создает соотношение один к одному между GBC, Calyces of Help и основными нейронами. Чашечная удержанная охватывает основной нейрон с отличительной морфологией: ветвление чашечки позволяет создавать сети второго и третьего порядка. Каждая ветвь устанавливает связь с основным нейроном, устанавливая большое количество активных зон. Это необычно для синаптических терминалов в мозге, так как большинство создает единую активную зону. [ 14 ] Каждая чашечка содержит от 300 до 700 активных зон, а в каждой из активных зон существует около 100 везикул, содержащих глутамат с примерно 3 пристывшими везикулами одновременно. Эти везикулы большие, что согласуется с результатами, касающимися количественных размеров в других синапсах для взрослых. Также присутствуют также плотные везикулы, обычно содержащие нейропептиды , но для определения их содержания и функции необходимы дальнейшие исследования. [ 15 ]
Для поддержания структуры синапса, как и в случае с другими синапсами, существует много микротрубочек . Чашечка имеет большое количество микротрубочек у основания терминала. Эти микротрубочки выполняют различные функции, такие как обеспечение стабильности для синапса, ограничение распределения синаптических пузырьков и локализация митохондрий . Митохондрии имеют три важные функции на синаптическом терминале: позволяя синапсу удовлетворить метаболические потребности (особенно для удаления кальция после деполяризации ), буферизацию кальция, позволяя поглощать кальций в митохондрии и обеспечивая энергию для синтеза глутамата. [ 12 ]
Различные глиальные клетки также связаны с чашечкой удерживаемой. Два типа глиальных клеток окружают чашечку: астроциты и NG 2 глиальные клетки . Астроциты экспрессируют переносчики глутамата для удаления глутамата из синапса. Это единственный известный механизм удаления глутамата из синапса. Глиальные клетки NG 2 экспрессируют рецепторы AMPA. [ 12 ]
Разработка
[ редактировать ]Общее развитие
[ редактировать ]Во второй день после второго дня (P2) образуется незрелая чашечка крысы, легко отличающаяся от его характерной морфологии закрытой ложки . [ 12 ] Первичные синаптические контакты, которые образуют чашечку, собираются между нейронами MNTB (медиальное ядро трапеции тела) и VCN (вентральный кохлеарный нерв), в конечном итоге соединяясь друг с другом, проецируя через среднюю линию двух областей. Эти ассоциации начинают появляться сразу после создания нейронов VCN; Можно наблюдать самое раннее образование этих контактов вокруг эмбрионального дня 17 (E17). Эти нейрональные связи, которые составляют важную область улитки, образуют ветви друг с другом, которые заканчиваются в чашечке. В течение следующих двух-трех недель нейрональные контакты, которые сначала образовали эмбриональную чашечку, эволюционируют в форме и функции, кульминацией которых сталкивается зрелая чашечка, которая облегчает последовательное, быстрое распространение сигналов в области MntB-VCN. [ 14 ]
Во время раннего развития нейронов возникают несколько процессов, чтобы обеспечить правильное образование чашечки, в частности, за счет влияния фактора фибробластического фактора роста (FGF), фактора транскрипции Math5, молекулы распознавания нейронных клеток NB-2 и белков эфрина (EPH) в клетках. Math1/Math5 и FGF являются двумя регуляторами, необходимыми для соответствующего роста и развития комплекса кохлеарного ядра , который включает в себя как вентральное кохлеарное ядро (VCN), так и дорсальное кохлеарное ядро (DCN). Достаточные уровни FGF обеспечивают правильную морфологию кохлеарных ядер, в то время как Math5 гарантирует правильный размер и обработку кохлеарного ядра. Math1, еще один фактор транскрипции, необходим для появления нейронов VCN в кохлеарном внешнем потоке, а также нейронах верхнего оливари -комплекса . NB-2 также помогает в продвижении формирования чашечки, а также способствует содержанию контралатерального MNTB. Комбинированные эффекты этих трех молекул друг с другом иллюстрируют тот факт, что существует много семейств белков, участвующих в правильной передаче сигналов и образования чашечки. [ 14 ]

Кроме того, белки EPH являются неотъемлемой частью дальнейшей разработки системы слуховых схем после начальной эмбриональной формирования чашечки. Одной из характеристик, которая отличает белки EPH и их рецепторы от других систем сигнализации, является их способность передавать информацию двунаправленной. Переменная и обратная передача сигналов в клетках VCN и MNTB необходима для правильного числа и образования VCN и ипсилатеральных проекций MNTB в чашечке. Белки EPH также гарантируют, что в то время как аксоны проходят через ипсилатеральный MNTB, разветвление и окончательное прекращение этих проекций происходят только в контралатеральном MNTB, возможно, потому, что белки нацелены только на определенные области на аксонах. [ 14 ]
В целом, есть два ультраструктурных изменения, которые происходят в чашечке удерживаемой. Во -первых, во второй неделе развития миелинизация аксонов VCN в MNTB увеличивается. Этот заметный рост в миелине соответствует хронологическому развитию сигнальной схемы и адаптации чашечки. Второе ультраструктурное изменение включает в себя основные нейроны MNTBS, чьи клеточные тела и ядра увеличиваются в площади поверхности из -за увеличения. Это является прямым результатом индивидуальной, большей постсинаптической плотности, распадающихся на меньшие, многочисленные плотности. [ 14 ]
Разработка калиевого канала
[ редактировать ]
Калиевые каналы жизненно важны для проведения потенциала пресинаптического действия . Чашечка содержит несколько типов калиевых каналов, каждая из которых различается по местоположению и чувствительности. Оба низкопороговых k + каналы и высокопороговый тип выпрямителя K + Каналы присутствуют в пресинаптических нейронах. [ 15 ] Есть четыре низкопороговых + Каналы присутствуют: k v 1.1, k v 1,2, k v 1,3 и k v 7.5. K V 1.1 и K V 1.2 расположены в переходной зоне между аксоном и терминалом, а K V 1,3 K V 7,5 расположены в чашечке. [ 15 ] Существует активированный кальциевый калиевый канал, экспрессированный в чашечке, однако этот тип канала не способствует высвобождению нейротрансмиттера. [ 12 ]
В течение одной недели субъекты мышей (P7 до P14) показали, что плотность K V 1 и K v 3 низкого порогового канала увеличивается, что, в свою очередь, влияет на кинетику каналов. [ 15 ]
Разработка натриевого канала
[ редактировать ]Изменения в натриевых каналах во время созревания позволяют увеличить скорость потенциала пресинаптического действия. Здесь потенциалы действия становятся быстрее благодаря способности натриевых каналов быстрее восстанавливаться после проводимости . Данные показывают, что экспрессия в альфа -субъединице NA V 1,6 , специфического типа натриевого канала, отвечает за повышенную скорость передачи. NA V 1.2 , другой натриевый канал, выраженный в аксонах и узлах, известен, демонстрирует более медленную кинетику. [ 14 ]
Чтобы компенсировать миелинизацию (повышенную емкость ), ведущую к чашечке в последнем узле (область между миелиновой оболочкой) до того, как терминал аксона содержит высокую плотность NA + Каналы, чтобы позволить большому притоку (внутреннему потоку) натрия, чтобы запустить управляемые напряжением кальциевые каналы открываться в пресинаптическом терминале, вызывая приток кальция.
Разработка кальциевого канала
[ редактировать ]Незрелая чашечка удерживающей кальции (CA 2+ ) ионы входят в нейроны MNTB через N- , P/Q- и R-тип CA 2+ каналы , но в зрелом Calyces, CA 2+ Приток происходит в основном через каналы P/Q-типа. [ 14 ] N- и r- тип ca 2+ Рецепторы менее склонны запускать высвобождение пузырьков, так как эти типы рецепторов находятся дальше от сайтов высвобождения. Следовательно, ионы кальция, входящие в каналы N- и R-, увеличивают концентрацию ионов кальция в областях, более важных для функции чашечки.
Блокировка CA 2+ Каналы могут возникать с помощью G-белковых рецепторов , активированных следующими нейротрансмиттерами: [ 12 ]
- Норадреналин (норпинефрин)
- Серотонин
- γ-аминобутирическая кислота (ГАМК)
- Глутамат (глутаминовая кислота)
- Аденозин
Изменения происходят в ионных каналах, чтобы стимулировать более быструю передачу: [ 12 ]
- НА + и k + Каналы изменяются, чтобы позволить до- и постсинаптическим потенциалам действий быть быстрее
- K V 3 каналы также активируются гораздо быстрее.
- Размер пресинаптического CA 2+ токи увеличиваются.
- Стробирующая механика рецепторов глутамата становится быстрее
 |
 |
 |
 |
 |
Разработка канала, управляемого лигандами
[ редактировать ]Помимо глутаматного рецептора, лишь несколько других лигандных каналов в незрелых поэлиазах удерживались : ионотропный GABA A и рецептор глицина . Эти рецепторы позволяют хлорид (cl − ) течь через мембрану и из -за высокой концентрации хлорида в терминале эти рецепторы деполяризуются. [ 12 ]
Фенестрация
[ редактировать ]Между вторым и третьим послеродовыми неделями, во время начала слуха, чашечка Help разрабатывает свой характерный, высокоэлектрированный ( многие отверстия). [ 14 ] Фенестрация приводит к тому, что мембрана уменьшается до многочисленных небольших компартментов, что увеличивает соотношение поверхности к объему содержания чашечки. По мере того, как мембрана становится все более ущипнутыми в эти лампообразные структуры, синаптические пузырьки дополнительно сгруппированы в эти пространства, что приводит к увеличению количества пристыкованных везикул. [ 12 ]
Чтобы компенсировать доступные пространства в чашечке, для заполнения открытых пространств используются глиальные клетки с глутаматными рецепторами и транспортами, обеспечивая эффективное поглощение глутамата в синапсе.
Механизм
[ редактировать ]
В качестве синапса, чашечка HELD следует за механизмом, аналогичным другим синапсам. Тщательное описание можно найти в нейротрансмиссии .
Приток кальция
[ редактировать ]Приток кальция для незрелой чашечки удерживается опосредованными кальциевыми каналами N , P / Q- и R ; Однако при созревании только кальциевые каналы P/Q становятся доминирующими. [ 14 ] При притоке кальция незрелая чашечка удерживаемой очень реактивна из -за ее небольшой способности кальциевого буфера - это вызывает высвобождение глутамата даже при низких уровнях притока кальция. В рамках терминала, как и в случае с другими синапсами, два иона кальция связываются с синаптагмином , чтобы вызвать высвобождение пузырьков - для телятиков удерживания глутамат высвобождается в везикулах. В дополнение к высвобождению пузырьков, сигнал ионов кальция для терминала Calyx вернется в неактивное состояние. После притока кальция белок, связывающий элемент цАМФ-ответа (CREB), фосфорилируется , изменяя концентрации калия в клетке, чтобы вернуть терминал в неактивное состояние. [ 14 ] Удаление кальция осуществляется различными методами, в том числе: удаление из терминала, воспитание в митохондрии или связывание с кальциевыми связывающими белками, такими как парвальбумин и кальретинин . [ 12 ]
Пресинаптическое запрещение
[ редактировать ]Ретроградная передача сигналов необходима в чашечке для регулирования уровней кальция в пресинаптическом терминале. Активация метаботропных глутаматных рецепторов (MGLURS) активирует вторичный мессенджер G-белка, который взаимодействует с кальциевыми каналами P/Q-типа для снижения проводимости. Кроме того, размер пула везикул увеличивается, а вероятность высвобождения уменьшается. Другие методы пресинаптического ингибирования включают норадреналин, серотонин и аденозин - эти методы наблюдаются только в незрелых телевизиках удерживаемых. [ 14 ]
Постсинаптические глутаматные рецепторы
[ редактировать ]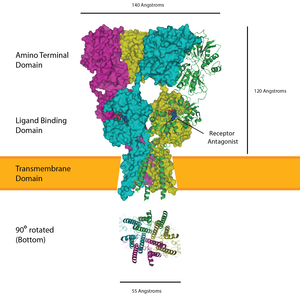
Рецепторы глутамата присутствуют на постсинаптическом терминале- два типа включают ионотропные AMPA и рецепторы NMDA . В качестве возбуждающего нейротрансмиттера, глутамат почти всегда вызывает запуск потенциала действия на постсинаптической стороне - дополнительно поощряется низким внутренним натрием основных нейронов. [ 12 ] В зрелой чашечке рецепторы AMPA сосредоточены на основном нейроне для локализации передачи для большей вероятности потенциала действия. Также обратите внимание, что вклад глутамата типа NMDA вклады рецепторов уменьшаются после начала слуха. [ 12 ]
Преснаптический везикул эндоцитоз
[ редактировать ]Механизм, лежащий в основе эндоцитоза синаптического везикула, по мере того, как чашечка становится более зрелой. Кальмодулин и кальциневрин в их активной форме необходимы для эндоцитоза пузырьков в незрелой чашечке; Однако в зрелой чашечке ни кальмодулин, ни кальциневрин не необходимы. Скорее, процесс опосредуется энергией, созданной гидролизом GTP. [ 14 ] Чтобы загрузить глутамат в везикулы в терминальных двух белках: везикулярный транспортер глутамата 1 (VGLUT1) и VGLUT2.
Ответ
[ редактировать ]Высокопороговые каналы калия в постсинаптической мембране позволяют быстро реполяризировать целевого нейрона. Низкопороговые каналы калия постсинаптического нейрона снижают возбудимость нейрона, чтобы ограничить его активацию только самым большим синаптическим входом. [ 12 ]
Важность исследования
[ редактировать ]Чашечка удержания стала популярной модельной системой в области нейробиологии. Присутствие этого синапса в нервной системе млекопитающих позволило провести прямые исследования в модели млекопитающих, а большой размер увеличивает простоту электрофизиологической записи. По этим причинам это было популярно в понимании выпуска передатчика.
В частности, чашечка удерживается из -за: [ 12 ]
- Простота записей пресинаптического зажима .
- Способность контролировать высвобождение передатчика при измерении до и постсинаптических эффектов.
- Простота визуализации и измерения емкости.
- Использование вирусов для наблюдения за чашечкой удерживаемой в качестве экзогенной системы экспрессии.
- Возможность провести эксперименты in vivo.
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Help, H. «Центральная линия слуха». Анат
- ^ Рюго, Дэвид К.; Spirou, George A. (2017-01-01), «Гигантские синаптические терминалы: конечные биологические и калисы слуховой системы ☆» , Справочный модуль в нейробиологии и биобедоральной психологии , Elsevier, ISBN 978-0-12-809324-5 Получено 2021-03-05
- ^ Sätzler, K.; Söhl, LF; Боллманн, JH; Борст, JG; Frotscher, M.; Sakmann, B.; Lübke, JH (2002). «Трехмерная реконструкция чашечки и его постсинаптического основного нейрона в медиальном ядре трапеции тела» . Журнал нейробиологии . 22 (24): 10567–10579. doi : 10.1523/jneurosci.22-24-10567.2002 . PMC 6758464 . PMID 12486149 . S2CID 18520148 .
- ^ Ян, Х.; Xu-Friedman, MA (2013). «Стохастические свойства высвобождения нейротрансмиттера расширяют динамический диапазон синапсов» . Журнал нейробиологии . 33 (36): 14406–14416. doi : 10.1523/jneurosci.2487-13.2013 . PMC 3761050 . PMID 24005293 .
- ^ Смит, Ph; Йорис, PX; Карни, LH; Инь, TCT (1991). «Проекции физиологически охарактеризованных глобулярных густых клеточных аксонов из кохлеарного ядра кошки». Журнал сравнительной неврологии . 304 (3): 387–407. doi : 10.1002/cne.903040305 . PMID 2022755 . S2CID 18844385 .
- ^ Смит, Ph; Йорис, PX; Инь, Т.С. (1998). «Анатомия и физиология основных клеток медиального ядра трапециевидного тела (MNTB) кошки». Журнал нейрофизиологии . 79 (6): 3127–3142. doi : 10.1152/jn.1998.79.6.3127 . PMID 9636113 .
- ^ Борст, JGG; Сория Ван Хове, Дж. (2012). «Чашечная синапс: от синапса модели до слухового реле». Ежегодный обзор физиологии . 74 : 199–224. doi : 10.1146/annurev-physiol-020911-153236 . PMID 22035348 .
- ^ Spangler, KM; Уор, WB; Хенкель, CK (1985). «Проекции основных клеток медиального ядра трапециевидного тела в кошке». Журнал сравнительной неврологии . 238 (3): 249–262. doi : 10.1002/cne.902380302 . PMID 4044914 . S2CID 35642962 .
- ^ Цучитани, С. (1997). «Вход из медиального ядра трапециевидного тела до детектора международного уровня». Исследование слуха . 105 (1–2): 211–224. doi : 10.1016/s0378-5955 (96) 00212-2 . PMID 9083818 . S2CID 4780393 .
- ^ Morest, DK (1968). «Обеспеченная система медиального ядра трапециевидного тела кошки, ее нейрональная архитектура и отношение к оливококлеарной пачке». Brain Res . 9 (2): 288–311. doi : 10.1016/0006-8993 (68) 90235-7 . PMID 5679830 .
- ^ Ryugo DK, Montey KL, Wright AL, Bennett ML, Pongstaporn T (2006). «Постнатальное развитие большого терминала слухового нерва: конечная удержанная у кошек». Исследование слуха . 216–217: 100–115. doi : 10.1016/j.heares.2006.01.007 . PMID 16497457 . S2CID 4676902 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Борст, JGG; Русу, С.И. (2012). «Глава 5: чашечка удерживаемого синапса». В Трусселле, Лоуренс; Поппер, Артур; Фэй, Ричард (ред.). Синаптические механизмы в слуховой системе . Нью-Йорк: Springer-Verlag. С. 95–134. ISBN 978-1-4419-9516-2 .
- ^ Цучитани, Чиеко (март 1997 г.). «Вход из медиального ядра трапециевидного тела до детектора международного уровня». Исследование слуха . 105 (1–2): 211–224. doi : 10.1016/s0378-5955 (96) 00212-2 . PMID 9083818 . S2CID 4780393 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Накамура, Пол А.; Крамер, Карина С. (июнь 2011 г.). «Образование и созревание чашечки» . Исследование слуха . 276 (1–2): 70–78. doi : 10.1016/j.heares.2010.11.004 . PMC 3109188 . PMID 21093567 .
- ^ Jump up to: а беременный в дюймовый Борст, Г.; Сория Ван Хове, Джон; Джерард, Дж. (17 марта 2012 г.). «Чашечная синапс: от синапса модели до слухового реле». Ежегодный обзор физиологии . 74 (1): 199–224. doi : 10.1146/annurev-physiol-020911-153236 . PMID 22035348 .