Липидно-заякоренный белок
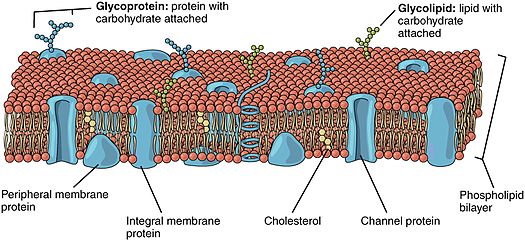
Белки, заякоренные в липидах (также известные как белки, связанные с липидами ), представляют собой белки, расположенные на поверхности клеточной мембраны. [ чего? ] которые ковалентно связаны с липидами , встроенными в клеточную мембрану. Эти белки встраиваются и занимают место в двухслойной структуре мембраны рядом с аналогичными хвостами жирных кислот . Заякоренный в липидах белок может располагаться по обе стороны клеточной мембраны. Таким образом, липид служит для закрепления белка на клеточной мембране. [ 1 ] [ 2 ] Они представляют собой разновидность протеолипидов .
Липидные группы играют роль во взаимодействии белков и могут способствовать функции белка, к которому они прикреплены. [ 2 ] Кроме того, липид служит медиатором мембранных ассоциаций или детерминантом специфических белок-белковых взаимодействий. [ 3 ] Например, липидные группы могут играть важную роль в повышении молекулярной гидрофобности . Это позволяет белкам взаимодействовать с клеточными мембранами и белковыми доменами . [ 4 ] В динамичной роли [ нужны разъяснения ] Липидация может изолировать белок от его субстрата, чтобы инактивировать белок, а затем активировать его путем презентации субстрата .
В целом, существует три основных типа белков, закрепленных на липидах, которые включают пренилированные белки , жирно-ацилированные белки и гликозилфосфатидилинозитол-связанные белки (GPI) . [ 2 ] [ 5 ] Белок может иметь несколько ковалентно связанных с ним липидных групп, но [ нужны разъяснения ] место, где липиды связываются с белком, зависит как от липидной группы, так и от белка. [ 2 ]
Пренилированные белки
[ редактировать ]
Пренилированные белки — это белки с ковалентно присоединенными гидрофобными полимерами изопрена (т.е. разветвленные пятиуглеродные углеводороды). [ 6 ] ) по цистеиновым остаткам белка. [ 2 ] [ 3 ] Более конкретно, эти изопреноидные группы, обычно фарнезил (15 атомов углерода) и геранилгеранил (20 атомов углерода), присоединяются к белку посредством тиоэфирных связей у остатков цистеина вблизи С-конца белка. [ 3 ] [ 4 ] Такое пренилирование липидных цепей до белков облегчает их взаимодействие с клеточной мембраной. [ 1 ]

Мотив пренилирования «CaaX-бокс» является наиболее распространенным сайтом пренилирования в белках, то есть местом, к которому ковалентно прикрепляются фарнезил или геранилгеранил. [ 2 ] [ 3 ] В последовательности CaaX-бокса C представляет собой пренилированный цистеин, A представляет собой любую алифатическую аминокислоту, а X определяет тип пренилирования, которое произойдет. Если X представляет собой Ala, Met, Ser или Gln, белок будет фарнезилироваться с помощью фермента фарнезилтрансферазы , а если X представляет собой Leu, то белок будет геранилгеранилироваться с помощью геранилгеранилтрансферазы I. фермента [ 3 ] [ 4 ] Оба эти фермента схожи, каждый содержит две субъединицы. [ 7 ]
Роли и функции
[ редактировать ]
Пренилированные белки особенно важны для роста, дифференцировки и морфологии эукариотических клеток. [ 7 ] Более того, пренилирование белка представляет собой обратимую посттрансляционную модификацию клеточной мембраны. Это динамическое взаимодействие пренилированных белков с клеточной мембраной важно для их сигнальных функций и часто нарушается при болезненных процессах, таких как рак. [ 8 ] Более конкретно, Ras — это белок, который подвергается пренилированию с помощью фарнезилтрансферазы , и когда он включается, он может активировать гены, участвующие в росте и дифференцировке клеток. Таким образом, чрезмерная активация передачи сигналов Ras может привести к раку. [ 9 ] Понимание этих пренилированных белков и их механизмов сыграло важную роль в разработке лекарств для борьбы с раком. [ 10 ] Другие пренилированные белки включают членов семейств Rab и Rho, а также ламины . [ 7 ]
Некоторые важные цепи пренилирования, участвующие в ГМГ-КоА-редуктазы метаболическом пути [ 1 ] являются геранилгераниол , фарнезол и долихол . Эти полимеры изопрена (например, геранилпирофосфат и фарнезилпирофосфат ) участвуют в конденсации посредством ферментов, таких как пренилтрансфераза , которые в конечном итоге циклизуются с образованием холестерина . [ 2 ]
Жирно-ацилированные белки
[ редактировать ]Жирно -ацилированные белки представляют собой белки, которые были модифицированы посттрансляционно с целью включения ковалентного присоединения жирных кислот к определенным аминокислотным остаткам. [ 11 ] [ 12 ] Наиболее распространенными жирными кислотами, ковалентно связанными с белком, являются насыщенная миристиновая кислота (14 атомов углерода) и пальмитиновая кислота (16 атомов углерода). Белки могут быть модифицированы так, чтобы они содержали одну или обе эти жирные кислоты. [ 11 ]

N- миристоилирование
[ редактировать ]N -миристоилирование (т.е. присоединение миристиновой кислоты) обычно представляет собой необратимую модификацию белка, которая обычно происходит во время синтеза белка. [ 11 ] [ 13 ] в котором миризиновая кислота присоединена к α-аминогруппе N-концевого остатка глицина посредством амидной связи . [ 2 ] [ 12 ] Эту реакцию облегчает N -миристоилтрансфераза . Эти белки обычно начинаются с последовательности Met - Gly и либо с серина, либо с треонина в положении 5. [ 11 ] Белки, которые были миристоилированы, участвуют в каскаде сигнальной трансдукции , белок-белковых взаимодействиях и в механизмах, которые регулируют нацеливание и функцию белков. [ 13 ] Примером важности миристоилирования белка является апоптоз , запрограммированная гибель клеток. белка После того, как агонист смерти взаимодействующего домена BH3 (Bid) миристоилирован, он нацеливает белок на перемещение к митохондриальной мембране для высвобождения цитохрома c , что в конечном итоге приводит к гибели клеток. [ 14 ] Другими белками, которые миристоилированы и участвуют в регуляции апоптоза, являются актин и гельзолин .
S -пальмитоилирование
[ редактировать ]
S-пальмитоилирование (т.е. присоединение пальмитиновой кислоты) представляет собой обратимую модификацию белка, при которой пальмитиновая кислота присоединяется к специфическому остатку цистеина посредством тиоэфирной связи. [ 2 ] [ 11 ] Термин S-ацилирование также можно использовать, когда к пальмитоилированным белкам также присоединяются другие средние и длинные цепи жирных кислот. Консенсусная последовательность пальмитоилирования белка не выявлена. [ 11 ] Пальмитоилированные белки в основном обнаруживаются на цитоплазматической стороне плазматической мембраны, где они играют роль в трансмембранной передаче сигналов. Пальмитоильную группу можно удалить пальмитоилтиоэстеразами. Считается, что это обратное пальмитоилирование может регулировать взаимодействие белка с мембраной и, таким образом, играть роль в процессах передачи сигналов. [ 2 ] Кроме того, это позволяет регулировать субклеточную локализацию, стабильность и транспортировку белка. [ 15 ] Примером того, как пальмитоилирование белка играет роль в сигнальных путях клетки, является кластеризация белков в синапсе . Когда постсинаптический белок плотности 95 (PSD-95) пальмитоилируется, он ограничивается мембраной и позволяет ему связываться и кластеризовать ионные каналы в постсинаптической мембране. Таким образом, пальмитоилирование может играть роль в регуляции высвобождения нейромедиаторов. [ 16 ]
Пальмитоилирование опосредует сродство белка к липидным рафтам и облегчает кластеризацию белков. [ 17 ] Кластеризация может увеличить близость двух молекул. Альтернативно, кластеризация может изолировать белок от субстрата. Например, пальмитоилирование фосфолипазы D (PLD) отделяет фермент от его субстрата фосфатидилхолина. Когда уровни холестерина снижаются или уровни PIP2 увеличиваются, локализация, опосредованная пальмитатом, нарушается, фермент перемещается к PIP2, где он встречает свой субстрат и становится активным за счет презентации субстрата . [ 18 ] [ 19 ] [ 20 ]
белки GPI
[ редактировать ]
Гликозилфосфатидилинозитол-заякоренные белки (GPI-заякоренные белки) присоединены к молекулярной группе комплекса GPI посредством амидной связи карбоксильной группой белка с C-концевой . [ 21 ] Этот комплекс GPI состоит из нескольких основных компонентов, которые все связаны между собой: фосфоэтаноламин , линейный тетрасахарид (состоящий из трех маннозы и глюкозаминила) и фосфатидилинозитол . [ 22 ] Группа фосфатидилинозитола связана гликозидно с не-N-ацетилированным глюкозамином тетрасахарида. образуется фосфодиэфирная связь Затем между маннозой на невосстанавливающем конце (тетрасахарида) и фосфоэтаноламином . Фосфоэтаноламин тогда представляет собой амид, связанный с С-концом карбоксильной группы соответствующего белка. [ 2 ] Прикрепление GPI происходит за счет действия комплекса GPI-трансамидаза. [ 22 ] Цепи жирных кислот фосфатидилинозитола встраиваются в мембрану и, таким образом, закрепляют белок на мембране. [ 23 ] Эти белки расположены только на внешней поверхности плазматической мембраны. [ 2 ]
Роли и функции
[ редактировать ]Остатки сахара в тетрасахариде и остатки жирных кислот в фосфатидилинозитоловой группе различаются в зависимости от белка. [ 2 ] Такое большое разнообразие позволяет белкам GPI выполнять широкий спектр функций, включая действие в качестве гидролитических ферментов , молекул адгезии , рецепторов, ингибиторов протеаз и белков, регулирующих комплемент. [ 24 ] Кроме того, белки GPI играют важную роль в эмбриогенезе, развитии, нейрогенезе, иммунной системе и оплодотворении. [ 21 ] В частности, белок GPI IZUMO1R (также называемый JUNO в честь римской богини плодородия ) в плазме яйцеклетки играет важную роль в слиянии сперматозоида и яйцеклетки . Высвобождение белка IZUMO1R (JUNO) GPI из плазматической мембраны яйцеклетки не позволяет сперматозоидам сливаться с яйцеклеткой, и предполагается, что этот механизм может способствовать блокированию полиспермии на плазматической мембране яйцеклетки. [ 25 ] Другие роли, которые допускает модификация GPI, заключаются в ассоциации с мембранными микродоменами, временной гомодимеризации или апикальной сортировке в поляризованных клетках. [ 21 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Джеральд Карп (2009). Клеточная и молекулярная биология: концепции и эксперименты . Джон Уайли и сыновья. стр. 128–. ISBN 978-0-470-48337-4 . Проверено 13 ноября 2010 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Воет Д., Воет Дж.Г., Пратт К.В. (2013). Основы биохимии: жизнь на молекулярном уровне (4-е изд.). John Wiley & Sons, Inc. с. 263. ИСБН 978-0470-54784-7 .
- ^ Перейти обратно: а б с д и Кейси Пи Джей, Seabra MC (март 1996 г.). «Белковые пренилтрансферазы» . Журнал биологической химии . 271 (10): 5289–92. дои : 10.1074/jbc.271.10.5289 . ПМИД 8621375 .
- ^ Перейти обратно: а б с Novelli G, D'Apice MR (сентябрь 2012 г.). «Фарнезилирование белков и болезни». Журнал наследственных метаболических заболеваний . 35 (5): 917–26. дои : 10.1007/s10545-011-9445-y . ПМИД 22307208 . S2CID 11555502 .
- ^ Фергюсон, Массачусетс (август 1991 г.). «Липидные якоря на мембранных белках». Современное мнение в области структурной биологии . 1 (4): 522–9. дои : 10.1016/s0959-440x(05)80072-7 .
- ^ изопрен (2003). «Энциклопедия Миллера-Кина и словарь медицины, сестринского дела и сопутствующего здоровья, седьмое изд.» . Проверено 28 ноября 2015 г.
- ^ Перейти обратно: а б с Лейн КТ, Биз Л.С. (апрель 2006 г.). «Серия тематических обзоров: посттрансляционные модификации липидов. Структурная биология белков фарнезилтрансферазы и геранилгеранилтрансферазы типа I» . Журнал исследований липидов . 47 (4): 681–99. doi : 10.1194/jlr.R600002-JLR200 . ПМИД 16477080 .
- ^ Штейн В., Кубала М.Х., Стин Дж., Гриммонд С.М., Александров К. (01.01.2015). «К систематическому картированию и разработке механизма пренилирования белков у Saccharomyces cerevisiae» . ПЛОС ОДИН . 10 (3): e0120716. Бибкод : 2015PLoSO..1020716S . дои : 10.1371/journal.pone.0120716 . ПМЦ 4358939 . ПМИД 25768003 .
- ^ Гудселл Д.С. (1 января 1999 г.). «Молекулярная перспектива: онкоген ras» . Онколог . 4 (3): 263–4. doi : 10.1634/теонколог.4-3-263 . ПМИД 10394594 .
- ^ Reuter CW, Морган М.А., Бергманн Л. (сентябрь 2000 г.). «Нацеливание на сигнальный путь Ras: рациональное, основанное на механизмах лечение гематологических злокачественных новообразований?». Кровь . 96 (5): 1655–69. дои : 10.1182/blood.V96.5.1655 . ПМИД 10961860 .
- ^ Перейти обратно: а б с д и ж Реш, доктор медицинских наук (ноябрь 2006 г.). «Торговля и передача сигналов жирноацилированными и пренилированными белками». Химическая биология природы . 2 (11): 584–90. дои : 10.1038/nchembio834 . ПМИД 17051234 . S2CID 9734759 .
- ^ Перейти обратно: а б Уилсон Дж.П., Рагхаван А.С., Ян Ю.И., Чаррон Дж., Ханг Х.К. (март 2011 г.). «Протеомный анализ жирноацилированных белков в клетках млекопитающих с помощью химических репортеров выявил S-ацилирование вариантов гистона H3» . Молекулярная и клеточная протеомика . 10 (3): М110.001198. дои : 10.1074/mcp.M110.001198 . ПМК 3047146 . ПМИД 21076176 .
- ^ Перейти обратно: а б Фарази Т.А., Ваксман Г., Гордон Дж.И. (октябрь 2001 г.). «Биология и энзимология N-миристоилирования белков» . Журнал биологической химии . 276 (43): 39501–4. дои : 10.1074/jbc.R100042200 . ПМИД 11527981 .
- ^ Мартин Д.Д., Бошан Э., Бертиаум Л.Г. (январь 2011 г.). «Посттрансляционное миристоилирование: жир имеет значение в клеточной жизни и смерти». Биохимия . Биоактивные липиды, питание и здоровье. 93 (1): 18–31. дои : 10.1016/j.biochi.2010.10.018 . ПМИД 21056615 .
- ^ Айкарт-Рамос К., Валеро Р.А., Родригес-Креспо I (декабрь 2011 г.). «Пальмитоилирование белков и субклеточный транспорт». Biochimica et Biophysical Acta (BBA) – Биомембраны . 1808 (12): 2981–94. дои : 10.1016/j.bbamem.2011.07.009 . ПМИД 21819967 .
- ^ Дитятьев, Александр (2006). Эль-Хусейни, Алаа (ред.). Молекулярные механизмы синаптогенеза . Нью-Йорк: Спрингер. стр. 72–75.
- ^ Левенталь, И.; Лингвуд, Д.; Гжибек, М.; Джошкун, У.; Саймонс, К. (3 декабря 2010 г.). «Пальмитоилирование регулирует сродство рафта к большинству интегральных белков рафта» . Труды Национальной академии наук . 107 (51): 22050–22054. Бибкод : 2010PNAS..10722050L . дои : 10.1073/pnas.1016184107 . ПМК 3009825 . ПМИД 21131568 .
- ^ Петерсен, Э.Н.; Чунг, Х.В.; Наебосадри, А; Хансен, С.Б. (15 декабря 2016 г.). «Кинетическое разрушение липидных рафтов является механосенсором фосфолипазы D». Природные коммуникации . 7 : 13873. Бибкод : 2016NatCo...713873P . дои : 10.1038/ncomms13873 . ПМК 5171650 . ПМИД 27976674 .
- ^ Робинсон, CV; Рохач, Т; Хансен, SB (сентябрь 2019 г.). «Инструменты для понимания наномасштабной липидной регуляции ионных каналов» . Тенденции биохимических наук . 44 (9): 795–806. дои : 10.1016/j.tibs.2019.04.001 . ПМК 6729126 . ПМИД 31060927 .
- ^ Петерсен, Э.Н.; Павел, М.А.; Ван, Х; Хансен, С.Б. (28 октября 2019 г.). «Нарушение пальмитат-опосредованной локализации; общий путь силовой и анестезирующей активации каналов TREK-1» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1862 (1): 183091. doi : 10.1016/j.bbamem.2019.183091 . ПМК 6907892 . ПМИД 31672538 .
- ^ Перейти обратно: а б с Киносита Т., Фудзита М. (январь 2016 г.). «Биосинтез белков, закрепленных на GPI: особый акцент на ремоделировании липидов GPI» . Журнал исследований липидов . 57 (1): 6–24. дои : 10.1194/jlr.R063313 . ПМЦ 4689344 . ПМИД 26563290 .
- ^ Перейти обратно: а б Икезава, Хиро (01 января 2002 г.). «Гликозилфосфатидилинозитол (GPI)-заякоренные белки» . Биологический и фармацевтический вестник . 25 (4): 409–417. дои : 10.1248/bpb.25.409 . ПМИД 11995915 .
- ^ Киносита Т., Фудзита М. (январь 2016 г.). «Биосинтез белков, закрепленных на GPI: особый акцент на ремоделировании липидов GPI» . Журнал исследований липидов . 57 (1): 6–24. дои : 10.1194/jlr.R063313 . ПМЦ 4689344 . ПМИД 26563290 .
- ^ Киношита Т (2014). «Биосинтез и дефицит гликозилфосфатидилинозитола» . Труды Японской академии. Серия Б, Физические и биологические науки . 90 (4): 130–43. Бибкод : 2014PJAB...90..130K . дои : 10.2183/pjab.90.130 . ПМК 4055706 . ПМИД 24727937 .
- ^ Кунрод С.А., Нааби-Хансен С., Шетти Дж., Шибахара Х., Чен М., Уайт Дж.М., Герр Дж.К. (март 1999 г.). «Обработка мышиных ооцитов PI-PLC высвобождает кластеры белков массой 70 кДа (pI 5) и от 35 до 45 кДа (pI 5,5) с поверхности яйцеклетки и ингибирует связывание и слияние сперматозоолеммы» . Биология развития . 207 (2): 334–49. дои : 10.1006/dbio.1998.9161 . ПМИД 10068467 .
Внешние ссылки
[ редактировать ] СМИ, связанные с липидно-заякоренным белком, на Викискладе?
СМИ, связанные с липидно-заякоренным белком, на Викискладе?