Клюв

Клюв клюв , птиц или рострум — это внешняя анатомическая структура, встречающаяся в основном у , а также у черепах , нептичьих динозавров и некоторых млекопитающих. Клюв используется для клевания , хватания и удерживания (при поиске пищи, поедания , манипулирования и переноса предметов, убийства добычи или борьбы), прихорашивания , ухаживания и кормления детенышей. Термины «клюв» и «рострум» также используются для обозначения аналогичной ротовой части у некоторых птицетазовых , птерозавров , китообразных , дицинодонтов , бесхвостых головастиков , однопроходных (т.е. ехидн и утконосов , имеющих клювоподобную структуру), сирен , иглобрюхов , марлиновых и головоногие моллюски .
Хотя клювы значительно различаются по размеру, форме, цвету и текстуре, они имеют схожую основную структуру. Два костных выступа – верхняя и нижняя челюсти – покрыты тонким ороговевшим слоем эпидермиса, известным как рамфотека. У большинства видов два отверстия, называемые ноздрями к дыхательной системе ведут .
Этимология
[ редактировать ]Хотя слово «клюв» в прошлом обычно ограничивалось заостренными клювами хищных птиц , [ 1 ] В современной орнитологии термины «клюв» и «клюв» обычно считаются синонимами. [ 2 ] Слово, датируемое 13 веком, происходит от среднеанглийского bec (через англо-французский ), который сам по себе происходит от латинского beccus . [ 3 ]
Анатомия
[ редактировать ]
Хотя клювы значительно различаются по размеру и форме от вида к виду, их основные структуры имеют схожий рисунок. Все клювы состоят из двух челюстей, обычно известных как верхняя (верхняя) и нижняя (нижняя). [ 4 ] (стр. 147) Верхняя, а в некоторых случаях нижняя челюсти укреплены внутри сложной трехмерной сетью костных спикул (или трабекул ), расположенных в мягкой соединительной ткани и окруженных твердыми внешними слоями клюва. [ 5 ] (стр. 149) [ 6 ] Челюстной аппарат птиц состоит из двух узлов: четырехзвенного рычажного механизма и одного пятизвенного рычажного механизма. [ 7 ]
Мандибулы
[ редактировать ]
Верхняя нижняя челюсть поддерживается трехзубчатой костью, называемой межчелюстной. Верхний зубец этой кости прикреплен ко лбу, а два нижних зубца прикрепляются к бокам черепа . У основания верхней челюсти к черепу прикрепляется тонкий листок носовых костей у носолобного шарнира, который придает верхней челюсти подвижность, позволяя ей двигаться вверх и вниз. [ 2 ]
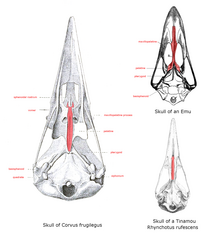
Основанием верхней челюсти, или крышей, если смотреть изо рта, является небо, строение которого у бескилевых сильно различается . Здесь сошник большой и соединяется с предчелюстными и челюстно-небными костями, образуя состояние, называемое «палеогнатным небом». У всех остальных современных птиц узкий раздвоенный сошник, который не соединяется с другими костями, поэтому его называют новорожденным. Форма этих костей варьируется в зависимости от семейства птиц. [ а ]
Нижняя челюсть поддерживается костью, известной как нижняя верхнечелюстная кость, — сложной костью, состоящей из двух отдельных окостеневших частей. Эти окостеневшие пластинки (или ветви ), которые могут иметь U-образную или V-образную форму, [ 4 ] (стр. 147) соединяются дистально (точное расположение сустава зависит от вида), но разделяются проксимально , прикрепляясь по обе стороны головы к квадратной кости. Челюстные мышцы, позволяющие птице закрывать клюв, прикрепляются к проксимальному концу нижней челюсти и к черепу птицы. [ 5 ] (стр. 148) Мышцы, прижимающие нижнюю челюсть, обычно слабы, за исключением некоторых птиц, таких как скворцы и вымершие гуйи , у которых хорошо развиты двубрюшные мышцы , помогающие добывать пищу путем подглядывания или раскрытия зева. [ 8 ] У большинства птиц эти мышцы относительно малы по сравнению с челюстными мышцами млекопитающих такого же размера. [ 9 ]
Рамфотека
[ редактировать ]Наружная поверхность клюва состоит из тонкой кератиновой оболочки, называемой рамфотекой . [ 2 ] [ 5 ] (стр. 148) которые можно разделить на ринотеку верхней челюсти и гнатотеку нижней челюсти. [ 10 ] (стр. 47) Этот покров возникает из мальпигиева слоя птиц эпидермиса . [ 10 ] (стр. 47) растут из пластинок у основания каждой нижней челюсти. [ 11 ] сосудистая прослойка Между рамфотекой и более глубокими слоями дермы имеется , прикрепляющаяся непосредственно к надкостнице костей клюва. [ 12 ] У большинства птиц рамфотека растет непрерывно, а у некоторых видов ее цвет меняется в зависимости от сезона. [ 13 ] У некоторых альцид , таких как тупики, части рамфотеки сбрасываются каждый год после сезона размножения, в то время как некоторые пеликаны сбрасывают часть клюва, называемую «рогом клюва», которая развивается в период размножения. [ 14 ] [ 15 ] [ 16 ]
Хотя большинство современных птиц имеют единственную цельную рамфотеку, виды относятся к нескольким семействам, включая альбатросов. [ 10 ] (стр. 47) и эму имеют сложные рамфотеки, которые состоят из нескольких частей, разделенных и ограниченных более мягкими ороговевшими бороздками. [ 17 ] Исследования показали, что это было примитивное наследственное состояние рамфотеки и что современная простая рамфотека возникла в результате постепенной утраты определяющих борозд в ходе эволюции. [ 18 ]
Томми
[ редактировать ]
Томии ) (единственное число томиум — это режущие края двух нижних челюстей. [ 10 ] (стр.598) У большинства птиц они варьируются от округлых до слегка острых, но у некоторых видов произошли структурные изменения, которые позволяют им лучше справляться с типичными источниками пищи. [ 19 ] Например, у зерноядных (семяядных) птиц на томии есть гребни, которые помогают птицам прорезать внешнюю оболочку семени . [ 20 ] У большинства соколов острый выступ вдоль верхней челюсти и соответствующая выемка на нижней челюсти. Они используют этот «зуб», чтобы смертельно перерезать позвонки своей жертве или разорвать насекомых на части. Некоторые коршуны , особенно те, которые охотятся на насекомых или ящериц, также имеют один или несколько таких острых выступов. [ 21 ] как и сорокопуты . [ 22 ] Томиальные зубы соколов подложены костью, а томиальные зубы сорокопутов полностью ороговевшие. [ 23 ] Некоторые рыбоядные виды, например крохали , имеют пилообразные зубцы вдоль томий, которые помогают им удерживать скользкую, извивающуюся добычу. [ 10 ] (стр. 48)
У птиц примерно 30 семейств томии по всей длине покрыты плотными пучками очень коротких щетинок. Большинство этих видов являются либо насекомоядными (предпочитающими добычу с твердым панцирем), либо питаются улитками , а щеткообразные выступы могут помочь увеличить коэффициент трения между челюстями, тем самым улучшая способность птиц удерживать твердую добычу. [ 24 ] Зубцы на клювах колибри , встречающиеся у 23% всех родов колибри, могут выполнять аналогичную функцию, позволяя птицам эффективно удерживать добычу насекомых. Они также могут позволить колибри с коротким клювом действовать как похитители нектара , поскольку они могут более эффективно удерживать и прорезать длинные или восковые цветочные венчики . [ 25 ] В некоторых случаях цвет томии птицы может помочь отличить схожие виды. У снежного гуся , например, клюв красновато-розовый с черными томиями, а у аналогичного гуся Росса весь клюв розовато-красный, без более темных томий. [ 26 ]
Саммит
[ редактировать ]
Кульмен – это дорсальный гребень верхней челюсти. [ 10 ] (стр. 127) По мнению орнитолога Э. Куэса. [ 4 ] до линии хребта крыши, это «самая высокая средняя продольная линия клюва» и проходит от точки, где верхняя челюсть выходит из перьев лба, до ее кончика. [ 4 ] (стр. 152) Длина клюва вдоль стебля является одним из регулярных измерений, проводимых при кольцевании (кольцовании) птиц. [ 27 ] и особенно полезен при исследованиях кормления. [ 28 ] Существует несколько стандартных измерений: от кончика клюва до точки начала растушевки на лбу, от кончика до переднего края ноздрей, от кончика до основания черепа или от кончика до восковица (для хищников и сов) [ 10 ] (стр. 342) — и ученые из разных уголков мира обычно отдают предпочтение одному методу перед другим. [ 28 ] Во всех случаях это измерения хорды (измеренные по прямой линии от точки к точке, без учета кривизны в кульме), выполненные штангенциркулем . [ 27 ]
Форма или цвет стебля также могут помочь идентифицировать птиц в поле. Например, стебель попугая- клеста сильно изогнут, а у очень похожего красного клеста изогнут более умеренно. [ 29 ] Стебель молодой обыкновенной гагары полностью темный, а у с очень похожим оперением молоди желтоклювой гагары стебель к кончику бледный. [ 30 ]
Конис
[ редактировать ]Гонис — это вентральный гребень нижней челюсти, образованный соединением двух ветвей кости или боковых пластинок. [ 10 ] (стр. 254) Проксимальный конец этого соединения, где две пластинки разделяются, известен как гонидальный угол или гонидальное расширение . У некоторых видов чаек пластинки в этом месте слегка расширяются, создавая заметную выпуклость; размер и форма гонидального угла могут быть полезны при идентификации сходных в остальном видов. Взрослые особи многих видов крупных чаек имеют красноватое или оранжевое пятно гонидального тела вблизи его расширения. [ 31 ] Это место вызывает попрошайничество у птенцов чаек . Птенец клюет место на клюве родителя, что, в свою очередь, стимулирует его срыгивать пищу. [ 32 ]
спайка
[ редактировать ]В зависимости от использования спайка может относиться к стыку верхней и нижней челюсти. [ 4 ] (стр. 155) или, наоборот, к сближению сомкнутых челюстей во всю длину, от углов рта до кончика клюва. [ 10 ] (стр. 105)
Зияние
[ редактировать ]
В анатомии птиц зияние — это внутренняя часть открытого рта птицы, а зияние — это область, где две челюсти соединяются вместе у основания клюва. [ 33 ] Ширина зева может быть фактором при выборе еды. [ 34 ]

Глаза молодых альтрициальных птиц часто ярко окрашены, иногда с контрастными пятнами или другими узорами, и считается, что это является показателем их здоровья, физической подготовки и конкурентоспособности. Исходя из этого, родители решают, как распределить еду между птенцами в гнезде. [ 35 ] У некоторых видов, особенно из семейств Viduidae и Estrildidae , на зеве имеются яркие пятна, известные как зияющие бугорки или зияющие сосочки. Эти узелковые пятна заметны даже при слабом освещении. [ 36 ] Исследование, изучающее зевы птенцов восьми видов воробьиных, показало, что зевы были заметны в ультрафиолетовом спектре (видимом для птиц, но не для человека). [ 37 ] Однако родители не могут полагаться исключительно на окраску зева, и другие факторы, влияющие на их решение, остаются неизвестными. [ 38 ]
В нескольких экспериментах было показано, что красный цвет зева стимулирует кормление. Эксперимент по манипулированию размером выводка и иммунной системой птенцов деревенских ласточек показал, что яркость зева положительно коррелирует с Т-клеточно -опосредованной иммунокомпетентностью , а больший размер выводка и инъекция антигена приводят к менее яркому зеву. [ 39 ] И наоборот, красное зияние обыкновенной кукушки ( Cuculus canorus ) не вызывало дополнительного кормления у родителей-хозяев. [ 40 ] Некоторые выводковые паразиты , такие как ястреб-кукушка Ходжсона ( C. fugax ), имеют цветные пятна на крыльях, имитирующие цвет зева зараженных видов. [ 41 ]
При рождении зияющие края птенца мясистые. По мере того, как птица становится птенцом , ее зияющие края остаются несколько опухшими и, таким образом, могут быть использованы для распознавания того, что конкретная птица молода. [ 42 ] К тому времени, когда он достигнет зрелого возраста, зияющие края уже не будут видны.
Нарес
[ редактировать ]
У большинства видов птиц есть наружные ноздри ( ноздри ), расположенные где-то на клюве. Ноздри представляют собой два отверстия круглой, овальной или щелевидной формы, которые ведут в носовые полости внутри черепа птицы и, следовательно, к остальной части дыхательной системы . [ 10 ] (стр. 375) У большинства видов птиц ноздри расположены в базальной трети верхней челюсти. Киви — заметное исключение; их ноздри расположены на кончиках клювов. [ 19 ] У некоторых видов нет внешних ноздрей. У бакланов и стрелотов, когда они птенцы, наружные ноздри примитивны, но они закрываются вскоре после того, как птицы оперяются ; взрослые особи этих видов (а также олуши и олуши всех возрастов, у которых также отсутствуют наружные ноздри) дышат ртом. [ 10 ] (стр. 47) Обычно две ноздри разделяет костная или хрящевая перегородка , но у некоторых семейств (включая чаек, журавлей и стервятников Нового Света) перегородка отсутствует. [ 10 ] (стр. 47) Хотя у большинства видов ноздри открыты, у некоторых групп птиц, включая тетерева и куропатки, ворон и некоторых дятлов, они покрыты перьями. [ 10 ] (стр. 375) Перья над ноздрями куропатки согревают воздух, который она вдыхает. [ 44 ] а те, что над ноздрями дятла, помогают предотвратить засорение носовых ходов частицами древесины. [ 45 ]
У видов птиц отряда Procellariformes ноздри заключены в двойные трубки, которые располагаются на верхней части или по бокам верхней челюсти. [ 10 ] (стр. 375) Эти виды, к которым относятся альбатросы, буревестники, ныряющие буревестники, качурки, глупыши и буревестники, широко известны как «тубенозы». [ 46 ] У ряда видов, в том числе у соколов небольшой костный бугорок , из ноздрей выступает . Функция этого бугорка неизвестна. Некоторые ученые предполагают, что он может действовать как перегородка, замедляя или рассеивая поток воздуха в ноздри (и, таким образом, позволяя птице продолжать дышать, не повреждая ее дыхательную систему) во время высокоскоростных ныряний, но эта теория не была подтверждена экспериментально. Такие бугорки есть не у всех видов, летающих с большой скоростью, в то время как у некоторых видов, летающих с низкой скоростью, они есть. [ 43 ]
Оперкулум
[ редактировать ]
Ноздри некоторых птиц прикрыты жаберной крышкой (множественная жаберная крышка ) — перепончатым, роговым или хрящевым лоскутом. [ 5 ] (стр. 117) [ 47 ] У ныряющих птиц жаберная крышка не допускает попадания воды в полость носа; [ 5 ] (стр. 117) когда птицы ныряют, сила удара воды закрывает жаберную крышку. [ 48 ] У некоторых видов, питающихся цветами, есть крышечки, которые помогают предотвратить засорение носовых ходов пыльцой. [ 5 ] (стр. 117) в то время как жаберные крышки двух видов Attagis бекаса помогают защититься от пыли. [ 49 ] Ноздри птенцов желтовато- коричневых лягушатников покрыты большими куполообразными крышками, которые помогают уменьшить быстрое испарение водяного пара, а также могут способствовать увеличению конденсации внутри самих ноздрей — обе важные функции, поскольку птенцы получают жидкость только из еду, которую им приносят родители. Эти крышки уменьшаются по мере старения птиц и полностью исчезают к тому времени, когда они достигают взрослой жизни. [ 50 ] У голубей жаберная крышка превратилась в мягкую набухшую массу, расположенную у основания клюва, над ноздрями; [ 10 ] (стр. 84) хотя его иногда называют восковицей , это другая структура. [ 4 ] (стр. 151) Тапакулос — единственные известные птицы, способные двигать крышечными крышками. [ 10 ] (стр. 375)
Розетка
[ редактировать ]Некоторые виды, например тупик , имеют мясистую розетку, которую иногда называют «зияющей розеткой». [ 51 ] в углах клюва. У тупика он выращивается как часть демонстрационного оперения. [ 52 ]
Просить
[ редактировать ]Птицы из нескольких семейств, в том числе хищные птицы, совы, поморники, попугаи, индейки и курасовы, имеют восковую структуру, называемую восковицей ( от латинского cera , что означает «воск») или церомой. [ 53 ] [ 54 ] который покрывает основу их счета. Эта структура обычно содержит ноздри , за исключением сов, у которых ноздри расположены дистальнее восковицы. Хотя у попугаев он иногда оперен, [ 55 ] восковица обычно голая и часто ярко окрашенная. [ 19 ] У хищников восковица является половым сигналом, указывающим на «качество» птицы; оранжевый цвет мозгов луня Монтегю коррелирует с массой его тела и физическим состоянием. Например, [ 56 ] Цвет восковицы молодых евразийских сплюшек имеет ультрафиолетовый (УФ) компонент, причем УФ-пик соответствует массе птицы. У цыпленка с меньшей массой тела УФ-пик имеет более высокую длину волны, чем у цыпленка с большей массой тела. Исследования показали, что совы-родители предпочитают кормить птенцов церерой, которая показывает пики УФ-излучения с более высокой длиной волны, то есть птенцами с меньшим весом. [ 57 ]
У некоторых видов по цвету или внешнему виду восковицы можно отличить самцов от самок. Например, у самца большого курасса есть желтая восковица, которой нет у самки (и молодых самцов). [ 58 ] Восковица у самца волнистого попугая королевского синего цвета, а у самки - очень бледно-голубого, белого или коричневого цвета. [ 59 ]
Ноготь
[ редактировать ]
У всех птиц семейства Anatidae (уток, гусей, лебедей) есть ноготь — пластинка из твердой роговой ткани на кончике клюва. [ 60 ] Эта щитовидная структура, которая иногда охватывает всю ширину клюва, часто загибается на кончике, образуя крючок. [ 61 ] Он служит разным целям в зависимости от основного источника пищи птицы. Большинство видов используют ногти, чтобы выкапывать семена из грязи или растительности. [ 62 ] в то время как ныряющие утки используют свой, чтобы вырвать моллюсков из камней. [ 63 ] Есть данные, что ноготь может помогать птице захватывать предметы. Виды, которые используют сильные хватательные движения, чтобы закрепить пищу (например, когда ловят и удерживают большую извивающуюся лягушку), имеют очень широкие ногти. [ 64 ] Под ногтем расположены определенные типы механорецепторов — нервных клеток, чувствительных к давлению, вибрации или прикосновению. [ 65 ]
Иногда форму или цвет когтей можно использовать, чтобы отличить похожие виды или водоплавающие птицы разного возраста. Например, у большой черпачки более широкий черный ноготь, чем у очень похожей на нее малой черпачки . [ 66 ] У молодых « серых гусей » когти темные, тогда как у большинства взрослых особей когти светлые. [ 67 ] Гвоздь дал семейству диких птиц одно из прежних названий: «Unguirostres» происходит от латинского ungus , что означает «ноготь», и rostrum , что означает «клюв». [ 61 ]
Риктальная щетина
[ редактировать ]Риктальные щетинки представляют собой жесткие, похожие на волосы перья, которые возникают вокруг основания клюва. [ 68 ] Они распространены среди насекомоядных птиц, но встречаются и у некоторых ненасекомоядных видов. [ 69 ] Их функция неясна, хотя было предложено несколько возможностей. [ 68 ] Они могут функционировать как «сеть», помогая поймать летающую добычу, хотя на сегодняшний день не существует никаких эмпирических доказательств, подтверждающих эту идею. [ 70 ] Есть некоторые экспериментальные данные, позволяющие предположить, что они могут предотвращать попадание частиц в глаза, если, например, добыча пропущена или разломана при контакте. [ 69 ] Они также могут помочь защитить глаза от частиц, встречающихся в полете, или от случайного контакта с растительностью. [ 70 ] Есть также свидетельства того, что риктальные щетинки некоторых видов могут функционировать тактильно, подобно тому, как это делают усы млекопитающих ( вибриссы ). Исследования показали, что тельца Гербста — в сочетании с риктальными щетинками обнаруживаются механорецепторы, чувствительные к давлению и вибрации. Они могут помочь в обнаружении добычи, навигации в затемненных полостях гнезда, сборе информации во время полета или обращении с добычей. [ 70 ]
Яичный зуб
[ редактировать ]
Доношенные птенцы большинства видов птиц имеют на клюве небольшой острый кальцинированный выступ, который они используют, чтобы вылезти из яйца . [ 10 ] (стр. 178) Этот белый шип, широко известный как яичный зуб, обычно находится на кончике верхней челюсти, хотя у некоторых видов он есть на кончике нижней челюсти, а у некоторых видов по одному на каждой нижней челюсти. [ 71 ] Несмотря на свое название, выступ не является настоящим зубом , как одноименные выступы некоторых рептилий ; вместо этого он является частью покровной системы , как и когти и чешуя . [ 72 ] Вылупившийся цыпленок сначала использует свой яичный зуб, чтобы сломать мембрану вокруг воздушной камеры на широком конце яйца. Затем он клюет яичную скорлупу, медленно вращаясь внутри яйца, в конечном итоге (в течение часов или дней) создавая в скорлупе серию небольших круглых трещин. [ 5 ] (стр. 427) Прорвав поверхность яйца, цыпленок продолжает колоть его, пока не проделает большую дырку. Ослабленное яйцо со временем разбивается под давлением движений птицы. [ 5 ] (стр. 428)
Яичный зуб настолько важен для успешного выхода из яйца, что птенцы большинства видов погибнут невылупившимися, если у них не получится его развить. [ 71 ] Однако есть несколько видов, у которых нет яичных зубов. У птенцов мегаподов еще в яйце есть яичный зуб, но они теряют его перед вылуплением. [ 5 ] (стр. 427) в то время как у цыплят киви он никогда не развивается; птенцы обеих семей покидают яйца, выпрыгивая наружу. [ 73 ] Большинство цыплят теряют яичные зубы в течение нескольких дней после вылупления. [ 10 ] (стр. 178) хотя буревестники держат свое почти три недели [ 5 ] (стр. 428) а у мраморных муррелей срок до месяца. [ 74 ] Обычно яичный зуб отпадает, хотя у певчих птиц он рассасывается. [ 5 ] (стр. 428)
Цвет
[ редактировать ]Цвет клюва птицы обусловлен концентрацией пигментов — прежде всего меланинов и каротиноидов — в слоях эпидермиса, включая рамфотеку. [ 75 ] Эумеланин , который содержится в голых частях тела многих видов птиц, отвечает за все оттенки серого и черного цвета; чем плотнее отложения пигмента в эпидермисе, тем темнее получается цвет. Феомеланин дает «земляные тона» от золотого и рыжего до различных оттенков коричневого. [ 76 ] : 62 Хотя считается, что феомеланин встречается в сочетании с эумеланином в клювах желтовато-коричневого, коричневого или рогового цвета, исследователям еще предстоит изолировать феомеланин из какой-либо структуры клюва. [ 76 ] : 63 За окраску большинства красных, оранжевых и желтых клювов отвечают более десятка типов каротиноидов. [ 76 ] : 64
Оттенок . цвета определяется точным сочетанием красных и желтых пигментов, а насыщенность определяется плотностью нанесенных пигментов Например, ярко-красный цвет создается плотными отложениями преимущественно красных пигментов, а тусклый желтый создается диффузными отложениями преимущественно желтых пигментов. Ярко-оранжевый цвет создается плотными отложениями красного и желтого пигментов примерно в равных концентрациях. [ 76 ] : 66 Окраска клюва помогает сделать демонстрацию использования этих клювов более заметной. [ 77 ] (стр. 155) птицы В целом цвет клюва зависит от сочетания гормонального состояния и рациона . Цвета обычно становятся самыми яркими по мере приближения сезона размножения и бледнее после размножения. [ 31 ]
Птицы способны видеть цвета в ультрафиолетовом диапазоне, а известно, что у некоторых видов на клюве имеются пики ультрафиолетового отражения (указывающие на наличие ультрафиолетового цвета). [ 78 ] Наличие и интенсивность этих пиков могут указывать на приспособленность птицы. [ 56 ] половая зрелость или статус парной связи. [ 78 ] королевских и императорских пингвинов Например, у пятна, отражающие ультрафиолет, появляются только во взрослом возрасте. Эти пятна ярче у парных птиц, чем у ухаживающих. Расположение таких пятен на клюве может иметь важное значение, позволяя птицам идентифицировать сородичей . Например, королевские и императорские пингвины с очень похожим оперением имеют на клювах пятна, отражающие УФ-излучение, в разных местах. [ 78 ]
Диморфизм
[ редактировать ]
Размер и форма клюва могут различаться как у разных видов, так и между ними; у некоторых видов размер и пропорции клюва у самцов и самок различаются. Это позволяет представителям обоих полов использовать разные экологические ниши, тем самым уменьшая внутривидовую конкуренцию . [ 79 ] Например, самки почти всех куликов имеют более длинные клювы, чем самцы того же вида. [ 80 ] а у самок американской шилоклювки клювы немного более загнуты вверх, чем у самцов. [ 81 ] У самцов более крупных видов чаек клювы больше и толще, чем у самок того же вида, а у неполовозрелых особей клювы могут быть меньше и более тонкие, чем у взрослых особей. [ 82 ] У многих птиц-носорогов наблюдается половой диморфизм в размере и форме как клювов, так и шлемов , а тонкий изогнутый клюв самки гуи был почти в два раза длиннее, чем прямой и толстый клюв самца. [ 10 ] (стр. 48)
Цвет также может различаться в зависимости от пола или возраста внутри вида. Обычно такая разница в цвете обусловлена наличием андрогенов . Например, у домашних воробьев меланины вырабатываются только в присутствии тестостерона ; кастрированные домашние воробьи-самцы, как и домашние воробьи-самки, имеют коричневые клювы. Кастрация также предотвращает нормальное сезонное изменение цвета клювов самцов черноголовых чаек и овсянок индиго . [ 83 ]
Разработка
[ редактировать ]Клюв современных птиц имеет сросшуюся предчелюстную кость, которая модулируется экспрессией гена Fgf8 в лобно-носовой эктодермальной зоне во время эмбрионального развития. [ 84 ]
Форма клюва определяется двумя модулями: предносовым хрящом на ранней эмбриональной стадии и предчелюстной костью на более поздних стадиях. Развитие предносового хряща регулируется генами Bmp4 и CaM , тогда как развитие предчелюстной кости контролируется TGFβllr , β-катенином и Dickkopf-3. [ 85 ] [ 86 ] TGFβllr кодирует серин/треониновую протеинкиназу, которая регулирует транскрипцию генов при связывании лиганда; предыдущие работы подчеркнули его роль в развитии черепно-лицевого скелета млекопитающих. [ 87 ] β-катенин участвует в дифференцировке терминальных костных клеток. Dickkopf-3 кодирует секретируемый белок, который, как известно, также экспрессируется при черепно-лицевом развитии млекопитающих. Комбинация этих сигналов определяет рост клюва по осям длины, глубины и ширины. Снижение экспрессии TGFβllr значительно уменьшало глубину и длину эмбрионального клюва курицы из-за недоразвития предчелюстной кости. [ 88 ] Напротив, увеличение передачи сигналов Bmp4 может привести к уменьшению предчелюстной кости из-за чрезмерного развития преназального хряща, который потребляет больше мезенхимальных клеток для формирования хряща, а не кости. [ 85 ] [ 86 ]
Функции
[ редактировать ]
Еда
[ редактировать ]Клювы разных видов развивались в зависимости от их рациона; например, у хищников есть острые клювы, которые облегчают рассечение и откусывание тканей жертвенных животных , тогда как воробьиные птицы, специализирующиеся на поедании семян с особенно прочной оболочкой (такие как гробоклювы и кардиналы ), имеют большие, толстые клювы с высокой сжимающей способностью ( по тому же принципу, что и придуманный человеком щелкунчик ). У птиц, которые зарабатывают себе на жизнь рыбной ловлей, клювы приспособлены для этого занятия; например, клювы пеликанов хорошо приспособлены для вычерпывания и проглатывания рыбы целиком. Клювы дятлов хорошо приспособлены для расклевывания древесины во время охоты на членистоногих .
Самозащитный клевок
[ редактировать ]Чтобы защититься, птицы могут кусать или колоть клювом. [ 89 ]
Демонстрации (для ухаживания, территориальности или сдерживания)
[ редактировать ]Некоторые виды используют свои клювы в различных проявлениях. Например, во время ухаживания самец гаргани прикасается клювом к синим зеркальным перьям на крыльях, изображая прихорашивающуюся демонстрацию, а самец утки-мандаринки делает то же самое со своими оранжевыми перьями паруса. [ 77 ] (стр. 20) Ряд видов используют зияющий открытый клюв, демонстрируя страх и/или угрозу. Некоторые дополняют впечатление шипением или тяжелым дыханием, а другие хлопают клювом. Краснобрюхие дятлы у кормушек для птиц часто машут своими грозными клювами конкурирующим птицам, которые подбираются слишком близко, ясно сигнализируя: «Это семя мое, вы не можете его получить».
Сенсорное обнаружение
[ редактировать ]Утконос использует свой клюв , чтобы ориентироваться под водой, находить пищу и копать. Клюв содержит электрорецепторы и механорецепторы, вызывающие мышечные сокращения, помогающие обнаружить добычу. Это один из немногих видов млекопитающих, использующих электрорецепцию . [ 90 ] [ 91 ] В клюве водоплавающих птиц содержатся тельца Грандри , которые помогают определять скорость во время фильтрационного кормления.
Прихорашиваюсь
[ редактировать ]Клюв птиц играет роль в удалении кожных паразитов ( эктопаразитов ), таких как вши. В основном это делает кончик клюва. Исследования показали, что введение насадки, чтобы птицы не могли использовать насадку, приводит к увеличению паразитной нагрузки на голубей. [ 92 ] Также было отмечено, что птицы, у которых клюв деформирован естественным путем, имеют более высокий уровень паразитов. [ 93 ] [ 94 ] [ 95 ] [ 96 ] Считается, что выступ на конце верхней части клюва (то есть той части, которая начинает загибаться вниз) скользит по нижнему клюву, раздавливая паразитов. [ 92 ]
Считается, что этот выступ клюва находится под действием стабилизирующего естественного отбора . Считается, что против очень длинных клювов следует выбирать, поскольку они склонны к большему количеству переломов, как это было продемонстрировано на сизых голубях. [ 97 ] Клювы без выступа не смогут эффективно удалять и убивать эктопаразитов, как упоминалось выше. Исследования подтвердили, что существует давление отбора на промежуточную величину вылета. При тестировании было обнаружено, что у западных кустовых сойок, у которых были более симметричные клювы (т.е. те, у которых меньше выступающих вперед), было обнаружено большее количество вшей. [ 98 ] Та же картина наблюдалась и при исследованиях перуанских птиц. [ 99 ]
Кроме того, поскольку клювы играют роль в чистке зубов, это является свидетельством совместной эволюции морфологии выступа клюва и морфологии тела паразитов. Было показано, что искусственное лишение птиц способности прихорашиваться с последующим восстановлением способности прихорашиваться приводит к изменению размеров тела вшей. После того, как способность птиц прихорашиваться была восстановлена, было обнаружено, что вши стали уменьшать размеры тела, что позволяет предположить, что они могут эволюционировать в ответ на давление со стороны птиц. [ 92 ] которые могли бы отреагировать, в свою очередь, изменениями в морфологии клюва. [ 92 ]
Коммуникативная перкуссия
[ редактировать ]Ряд видов, в том числе аисты , некоторые совы , лягушачьи рта и шумный шахтер , используют хлопанье в ладоши как форму общения. [ 77 ] (стр. 83) Известно, что некоторые виды дятлов используют перкуссию в качестве средства ухаживания, тогда как самцы привлекают ( на слух ) внимание самок на расстоянии, а затем впечатляют их громкостью и характером звука. Это объясняет, почему люди иногда доставляют неудобства клеванием, которое явно не имеет никакой цели кормления (например, когда птица неоднократно клюет листовой металл).
Теплообмен
[ редактировать ]Исследования показали, что некоторые птицы используют клювы, чтобы избавиться от лишнего тепла. Токо -тукан , имеющий самый большой клюв по сравнению с размером тела среди всех видов птиц, способен изменять приток крови к своему клюву. Этот процесс позволяет клюву работать как «переходный тепловой радиатор», который, как сообщается, может соперничать с ушами слона по своей способности излучать тепло тела. [ 100 ]
Измерения размеров клюва нескольких видов американских воробьев, обитающих в солончаках вдоль побережья Северной Америки, показывают сильную корреляцию с летними температурами, зарегистрированными в местах размножения воробьев; только широта показала гораздо более слабую корреляцию. Отводя избыточное тепло через свои клювы, воробьи могут избежать потери воды, которая потребовалась бы при испарительном охлаждении, что является важным преимуществом в ветреной среде обитания, где не хватает пресной воды. [ 101 ] Некоторые бескилевые , в том числе обыкновенный страус , эму и южный казуар , используют различные голые части своего тела (включая клювы), чтобы рассеивать до 40% вырабатываемого ими метаболического тепла. [ 102 ] С другой стороны, исследования показали, что птицы из более холодного климата (большие высоты или широты и более низкие температуры окружающей среды) имеют клювы меньшего размера, что уменьшает потерю тепла из этой структуры. [ 103 ]
Биллинг
[ редактировать ]
Во время ухаживания спаривающиеся пары многих видов птиц соприкасаются или сжимают клювы друг друга. Термин «биллинг» (также «неббинг» в британском английском), [ 104 ] такое поведение, по-видимому, укрепляет парные связи . [ 105 ]
Количество контактов варьируется в зависимости от вида. Некоторые нежно касаются лишь части клюва партнера, в то время как другие энергично стучат клювами друг о друга. [ 106 ]
Олуши высоко поднимают клювы и неоднократно ими цокают, самец тупика грызет клюв самки, самец свиристеля кладет свой клюв в рот самки, а вороны держат клювы друг друга в продолжительном «поцелуе». [ 107 ] Выставление счетов также может использоваться как жест умиротворения или подчинения. Подчиненные канадские сойки обычно клюют более доминирующих птиц, опуская тело и вздрагивая крыльями, как молодые еду . птицы, выпрашивающие при этом [ 108 ] Известно, что ряд паразитов, в том числе ринонисиды и Trichomonas Gallinae, передаются от птицы во время выклева. [ 109 ] [ 110 ]
Использование этого термина выходит за рамки поведения птиц; «Выставление счетов и воркование» в отношении ухаживания человека (особенно поцелуев) используется со времен Шекспира . [ 111 ] и происходит от ухаживания голубей. [ 112 ]
Обрезка клюва
[ редактировать ]Поскольку клюв является чувствительным органом со множеством сенсорных рецепторов, обрезка клюва (иногда называемая «обрезанием клюва») «очень болезненна». [ 113 ] птицам, над которыми это делается. Тем не менее, это регулярно делается в отношении интенсивно выращиваемых птиц , особенно в стадах-несушках и бройлерных стадах, потому что это помогает уменьшить ущерб, который стада наносят себе из-за ряда стрессовых поведений, включая каннибализм , клевание жерл и перьев . Прижигающим лезвием или инфракрасным лучом отрезается примерно половина верхнего клюва и около трети нижнего клюва. Боль и чувствительность могут сохраняться в течение недель или месяцев после процедуры, а невромы по краям разреза могут образовываться . Потребление пищи обычно снижается на некоторое время после обрезки клюва. обработанной птицы Однако исследования показывают, что надпочечники весят меньше, а уровень кортикостерона в плазме ниже, чем у необрезанной птицы, что указывает на то, что в целом они испытывают меньший стресс. [ 113 ]
Похожая, но отдельная практика, обычно выполняемая птичьим ветеринаром или опытным птицеводом, включает обрезку, подпиливание или шлифовку клювов содержащихся в неволе птиц в оздоровительных целях – чтобы исправить или временно облегчить разрастания или деформации и дать птице возможность лучше передвигаться. это обычная деятельность по кормлению и уборке. [ 114 ]
Среди владельцев хищников эта практика широко известна как «преодоление». [ 115 ]
орган Билла
[ редактировать ]
— Конечный орган клюва это область, обнаруженная возле кончика клюва у некоторых видов птиц, которые добывают пищу, в частности, зондированием. Эта область имеет высокую плотность нервных окончаний, известных как тельца Гербста . Он состоит из ямок на поверхности клюва, которые у живой птицы заняты клетками, реагирующими на изменения давления. Предполагается, что это позволяет птице осуществлять «дистанционное прикосновение», то есть она может обнаруживать движения животных, которых птица не касается напрямую. Виды птиц, которые, как известно, имеют «орган на кончике клюва», включают ибисов , куликов семейства Scolopacidae и киви . [ 116 ]
Есть предположение, что у этих видов орган кончика клюва лучше развит у видов, кормящихся во влажных средах обитания (толщина воды или мягкая грязь), чем у видов, использующих более наземный корм. Однако он был описан и у наземных птиц, в том числе у попугаев , которые известны своими ловкими методами добывания пищи. В отличие от зондирующих собирателей, тактильные ямки у попугаев встроены в твердый кератин (или рамфотеку ) клюва, а не в кость, и вдоль внутренних краев изогнутого клюва, а не на внешней стороне клюва. [ 117 ]
См. также
[ редактировать ]- Анатомия птиц – Анатомия птиц.
- Рострум (анатомия) - анатомический термин
- Морда - расширенная часть рта животного.
Сноски
[ редактировать ]- ^ Объяснение понятий desmognathous , aegithhognathous и т. д. с изображениями см. «Каталог видов» . 1891 г. - через Archive.org. .
Ссылки
[ редактировать ]- ^ Партингтон, Чарльз Фредерик (1835). Британская циклопедия естественной истории: объединение научной классификации животных, растений и минералов . Орр и Смит. п. 417 .
- ^ Перейти обратно: а б с Проктор, Ноубл С.; Линч, Патрик Дж. (1998). Руководство по орнитологии: Строение и функции птиц . Нью-Хейвен, Коннектикут: Издательство Йельского университета. п. 66. ИСБН 978-0-300-07619-6 .
- ^ «Клюв» . Мерриам-Вебстер . Проверено 1 июля 2016 г.
- ^ Перейти обратно: а б с д и ж
Куэс, Эллиотт (1890). Справочник по полевой и общей орнитологии . Лондон, Великобритания: Macmillan and Co., стр. 1 , 147, 151–152, 155. OCLC 263166207 . - ^ Перейти обратно: а б с д и ж г час я дж к
Гилл, Фрэнк Б. (1995). Орнитология (2-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman and Company. стр. 149, 427–428. ISBN 978-0-7167-2415-5 . - ^ Секи, Ясуаки; Бодд, Сара Г.; Мейерс, Марк А. (2009). «Клювы тукана и птицы-носорога: сравнительное исследование» (PDF) . Акта Биоматериалы . 6 (2): 331–343. doi : 10.1016/j.actbio.2009.08.026 . ПМИД 19699818 . Архивировано из оригинала (PDF) 2 апреля 2012 г.
- ^ Олсен, AM (3–7 января 2012 г.). За клювом: моделирование черепного кинезиса птиц и эволюция форм черепа птиц . Общество интегративной и сравнительной биологии. Чарльстон, Южная Каролина. Архивировано из оригинала 4 марта 2016 года . Проверено 27 июля 2015 г.
- ^ Майр, Джеральд (2005). «Новая эоценовая мышевидная птица, похожая на Chascacocolius (Aves: Coliiformes), с замечательной зияющей адаптацией» (PDF) . Организмы, разнообразие и эволюция . 5 (3): 167–171. дои : 10.1016/j.ode.2004.10.013 .
- ^ Кайзер, Гэри В. (2007). Внутренняя птица: анатомия и эволюция . Ванкувер, Британская Колумбия: UBC Press. п. 19. ISBN 978-0-7748-1343-3 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с
Кэмпбелл, Брюс; Лэк, Элизабет, ред. (1985). Словарь птиц . Карлтон, Англия: Т. и А.Д. Пойзер. ISBN 978-0-85661-039-4 . - ^ Гирлинг (2003), с. 4.
- ^ Самур (2000), с. 296.
- ^ Бонсер, Р.Х. и Виттер, М.С. (1993). «Твердость кератина клюва европейского скворца» (PDF) . Кондор . 95 (3): 736–738. дои : 10.2307/1369622 . JSTOR 1369622 .
- ^ Беддард, Фрэнк Э. (1898). Строение и классификация птиц . Лондон, Великобритания: Longmans, Green and Co. p. 5.
- ^ Питокчелли, Джей; Джон Ф. Пиатт; Гарри Р. Картер (2003). «Изменение оперения, линьки и морфологии усатой конюги ( Aethia pygmaea ) на Аляске». Журнал полевой орнитологии . 74 (1): 90–98. дои : 10.1648/0273-8570-74.1.90 . S2CID 85982302 .
- ^ Кнопф, Флорида (1974). «Расписание преддополнительной линьки белых пеликанов с примечаниями на роге клюва» (PDF) . Кондор . 77 (3): 356–359. дои : 10.2307/1366249 . JSTOR 1366249 .
- ^ Чернова, О.Ф.; Фадеева, Е.О. (2009). «Особенная архитектоника контурных перьев эму ( Dromaius novaehollandiae , Struthioniformes)». Доклады биологических наук . 425 : 175–179. дои : 10.1134/S0012496609020264 . S2CID 38791844 .
- ^ Иероним, Тобин Л.; Уитмер, Лоуренс М. (2010). «Гомология и эволюция птичьих сложных рамфотек». Аук . 127 (3): 590–604. дои : 10.1525/аук.2010.09122 . S2CID 18430834 .
- ^ Перейти обратно: а б с Штеттенхайм, Питер Р. (2000). «Покровная морфология современных птиц — обзор» . Интегративная и сравнительная биология . 40 (4): 461–477. дои : 10.1093/icb/40.4.461 .
- ^ Класинг, Кирк К. (1999). «Анатомия и физиология желудочно-кишечного тракта птиц». Семинары по медицине птиц и экзотических домашних животных . 8 (2): 42–50. дои : 10.1016/S1055-937X(99)80036-X .
- ^ Фергюсон-Лис, Джеймс; Кристи, Дэвид А. (1 января 2001 г.). Хищники мира . Лондон, Великобритания: Кристофер Хелм. п. 66. ИСБН 978-0-7136-8026-3 .
- ^ Харрис, Тони; Франклин, Ким (2000). Сорокопуты и кустарниковые сорокопуты . Лондон, Великобритания: Кристофер Хелм. п. 15. ISBN 978-0-7136-3861-5 .
- ^ В.Л. Белс; Ян К. Уишоу, ред. (2019). Питание у позвоночных: эволюция, морфология, поведение, биомеханика . Чам, Швейцария. ISBN 978-3-030-13739-7 . OCLC 1099968357 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Госнер, Кеннет Л. (июнь 1993 г.). «Скопат томия : приспособление для обращения с добычей с твердым панцирем?» (PDF) . Бюллетень Уилсона . 105 (2): 316–324.
- ^ Орнелас, Хуан Франциско. «Зубчатая Томия: адаптация к краже нектара у колибри?» (PDF) . Аук . 111 (3): 703–710.
- ^ Мэдж, Стив; Берн, Хилари (1988). Дикая птица . Лондон, Великобритания: Кристофер Хелм. стр. 143–144. ISBN 978-0-7470-2201-5 .
- ^ Перейти обратно: а б с Пайл, Питер; Хауэлл, Стив Н.Г.; Юник, Роберт П.; ДеСанте, Дэвид Ф. (1987). Справочник по идентификации воробьинообразных Северной Америки . Болинас, Калифорния: Slate Creek Press. стр. 6–7. ISBN 978-0-9618940-0-9 .
- ^ Перейти обратно: а б Боррас, А.; Паскаль, Дж.; Сенар, JC (осень 2000 г.). «Что измеряют различные размеры клюва и какой метод лучше всего использовать у зерноядных птиц?» (PDF) . Журнал полевой орнитологии . 71 (4): 606–611. дои : 10.1648/0273-8570-71.4.606 . JSTOR 4514529 . S2CID 86597085 .
- ^ Малларни, Свенссон, Зеттерстрем и Грант (1999) стр. 357
- ^ Малларни, Свенссон, Зеттерстрем и Грант (1999) стр. 15
- ^ Перейти обратно: а б Хауэлл (2007), с. 23.
- ^ Рассел, Питер Дж.; Вулф, Стивен Л.; Герц, Пол Э.; Старр, Сеси (2008). Биология: динамическая наука . Том. 2. Белмонт, Калифорния: Томсон Брукс / Коул. п. 1255. ИСБН 978-0-495-01033-3 .
- ^ Ньюман, Кеннет Б. (2000). Птицы Ньюмана по цветам . Струйк. п. 14. ISBN 978-1-86872-448-2 . [ постоянная мертвая ссылка ]
- ^ Уилрайт, Северная Каролина (1985). «Размер плода, ширина зева и рацион птиц, питающихся фруктами» (PDF) . Экология . 66 (3): 808–818. дои : 10.2307/1940542 . JSTOR 1940542 . Архивировано из оригинала (PDF) 8 апреля 2016 г. Проверено 31 октября 2013 г.
- ^ Солер, Джей-Джей; Авилес, ЖМ (2010). Хэлси, Льюис Джордж (ред.). «Конкуренция между братьями и сестрами и заметность зияний птенцов у альтрициальных птиц: сравнительное исследование» . ПЛОС ОДИН . 5 (5): е10509. Бибкод : 2010PLoSO...510509S . дои : 10.1371/journal.pone.0010509 . ПМЦ 2865545 . ПМИД 20463902 .
- ^ Хаубер, Марк и Ребекка М. Килнер (2007). «Коэволюция, общение и мимикрия хозяина и птенца у паразитических вьюрков: кто кого имитирует?» (PDF) . Поведение. Экол. Социобиол . 61 (4): 497–503. дои : 10.1007/s00265-006-0291-0 . S2CID 44030487 . Архивировано из оригинала (PDF) 20 марта 2012 г.
- ^ Хант, Сара; Килнер, Ребекка М.; Лэнгмор, Наоми Э.; Беннетт, Эндрю Т.Д. (2003). «Заметный ультрафиолетовый цвет рта у попрошаек цыплят» . Письма по биологии . 270 (Приложение 1): С-25–С-28. дои : 10.1098/rsbl.2003.0009 . ПМК 1698012 . ПМИД 12952627 .
- ^ Шуец, Джастин Г. (октябрь 2005 г.). «Снижение роста, но не выживаемость цыплят с измененным рисунком зева». Поведение животных . 70 (4): 839–848. дои : 10.1016/j.anbehav.2005.01.007 . ISSN 0003-3472 . S2CID 53170955 .
- ^ Никола, Саино; Роберто, Амброзини; Роберта, Мартинелли; Паола, Нинни; Андерс Папе, Мёллер (2003). «Окраска зева надежно отражает иммунокомпетентность птенцов ласточки ( Hirundo Rustica )» (PDF) . Поведенческая экология . 14 (1): 16–22. дои : 10.1093/beheco/14.1.16 . Архивировано из оригинала (PDF) 11 июля 2011 года . Проверено 27 июня 2010 г.
- ^ Благородный, Д.Г.; Дэвис, Северная Каролина; Хартли, ИК; Макрей, С.Б. (июль 1999 г.). «Красное зияние птенца кукушки ( Cuculus canorus ) не является сверхъестественным стимулом для трех общих хозяев». Поведение . 136 (9): 759–777. дои : 10.1163/156853999501559 . JSTOR 4535638 .
- ^ Танака, Кейта Д.; Моримото, генерал; Уэда, Кейсуке (2005). «Желтое пятно на крыле птенца ястреба-кукушки Хорсфилда Cuculus fugax вызывает непонимание со стороны хозяев: имитирует зияние?» . Журнал птичьей биологии . 36 (5): 461–64. дои : 10.1111/j.2005.0908-8857.03439.x . Архивировано из оригинала 21 октября 2012 г.
- ^ Зикфуз, Джули. «Таинственные птицы двора» . Дайджест орнитолога . Проверено 25 июня 2010 г.
- ^ Перейти обратно: а б Капайноло, Питер; Батлер, Кэрол (2010). Насколько быстро может нырнуть сокол? . Нью-Брансуик, Нью-Джерси: Издательство Университета Рутгерса. п. 51 . ISBN 978-0-8135-4790-9 .
- ^ Геллхорн, Джойс (2007). Белохвостая куропатка: Призраки высокогорной тундры . Боулдер, Колорадо: Johnson Books. п. 110. ИСБН 978-1-55566-397-1 .
- ^ Эрлих, Пол Р.; Добкин, Дэвид С.; Сыворотка, Дэррил (1998). Справочник птицелова: полевой справочник по естественной истории птиц Северной Америки . Нью-Йорк, штат Нью-Йорк: Саймон и Шустер. п. 209 . ISBN 978-0-671-65989-9 .
- ^ Карбонерас, Карлос (1992). «Семейство Diomedeidae (Альбатросы)». Ин дель Ойо, Хосеп; Эллиотт, Эндрю; Саргатал, Джорди (ред.). Справочник птиц мира . Том. 1: От страуса до уток. Барселона: Lynx Edicions. п. 199 . ISBN 978-84-87334-10-8 .
- ^ Уитни, Уильям Дуайт; Смит, Бенджамин Эли (1911). Словарь и циклопедия Century . Том. 6. Нью-Йорк: Компания Century. п. 4123. LCCN 11031934 .
- ^ Бок, Уолтер Дж. (1989). «Организмы как функциональные машины: объяснение связности» . Американский зоолог . 29 (3): 1119–1132. дои : 10.1093/icb/29.3.1119 . JSTOR 3883510 .
- ^ Тадж, Колин (2009). Птица: естественная история того, кто такие птицы, откуда они пришли и как они живут . Нью-Йорк, штат Нью-Йорк: Издательство Crown Publishers. п. 140 . ISBN 978-0-307-34204-1 .
- ^ Каплан, Гизела Т. (2007). Тони Фрогмут . Коллингвуд, Виктория: Cyrus Publishing. стр. 100-1 40 –4 ISBN 978-0-643-09239-6 .
- ^ Харрис, Майк П. (2014). «Старение атлантических тупиков Fratercula arctica летом и зимой» (PDF) . Морская птица . 27 . Центр экологии и гидрологии: 22–40. Архивировано из оригинала (PDF) 11 июня 2016 г.
- ^ «Тупок с острова Скомер» (PDF) . www.welshwildlife.org (информационный бюллетень). Май 2011.
- ^ Полный словарь английского языка Вебстера
- ^ Элеонора Лоуренс (2008). Биологический словарь Хендерсона (14-е изд.). Пирсон Бенджамин Каммингс Прентис Холл. п. 111. ИСБН 978-0-321-50579-8 .
- ^ Юпитер, Тони; Парр, Майк (2010). Попугаи: Путеводитель по попугаям мира . А&С Черный. п. 17. ISBN 978-1-4081-3575-4 .
- ^ Перейти обратно: а б Мужо, Франсуа; Арройо, Беатрис Э. (22 июня 2006 г.). «Отражение ультрафиолета в мозговиках хищников» . Письма по биологии . 2 (2): 173–176. дои : 10.1098/rsbl.2005.0434 . ПМК 1618910 . ПМИД 17148356 .
- ^ Парехо, Желаемый; Авилес, Хесус М.; Родригес, Хуан (23 апреля 2010 г.). «Визуальные сигналы и родительский фаворитизм у ночной птицы» . Письма по биологии . 6 (2): 171–173. дои : 10.1098/rsbl.2009.0769 . ПМК 2865047 . ПМИД 19864276 .
- ^ Леопольд, Альдо Старкер (1972). Дикая природа Мексики: дичь, птицы и млекопитающие . Беркли, Калифорния: Издательство Калифорнийского университета. п. 202 . ISBN 978-0-520-00724-6 .
- ^ Олдертон, Дэвид (1996). Руководство для птицеводов по волнистым попугайчикам . Тетра Пресс. п. 12.
- ^ Кинг и Маклелланд (1985) п. 376
- ^ Перейти обратно: а б Эллиот, Даниэль Жиро (1898). Дикая птица Соединенных Штатов и британских владений . Нью-Йорк, штат Нью-Йорк: Ф. П. Харпер. п. XVIII. LCCN 98001121 .
- ^ Перринс, Кристофер М. (1974). Птицы . Лондон, Великобритания: Коллинз. п. 24. ISBN 978-0-00-212173-6 .
- ^ Петри, Чак (2006). Почему утки так делают: объяснены и сфотографированы 40 отличительных особенностей поведения уток . Миноква, Висконсин: Willow Creek Press. п. 31. ISBN 978-1-59543-050-2 .
- ^ Гудман, Дональд Чарльз; Фишер, Харви И. (1962). Функциональная анатомия пищевого аппарата водоплавающих птиц (Aves: Anatidae) . Карбондейл, Иллинойс: Издательство Университета Южного Иллинойса. п. 179. OCLC 646859135 .
- ^ Кинг и Маклелланд (1985) п. 421
- ^ Данн, Джон Л.; Альдерфер, Джонатан, ред. (2006). Полевой справочник по птицам Северной Америки (5-е изд.). Вашингтон, округ Колумбия: National Geographic. п. 40. ИСБН 978-0-7922-5314-3 .
- ^ Малларни, Свенссон, Зеттерстрем и Грант (1999) стр. 40
- ^ Перейти обратно: а б Ледерер, Роджер Дж. «Роль птичьих риктальных щетинок» (PDF) . Бюллетень Уилсона . 84 (2): 193–197.
- ^ Перейти обратно: а б Коновер, Майкл Р.; Миллер, Дон Э. (ноябрь 1980 г.). «Функция риктальной щетины у ивовой мухоловки» (PDF) . Кондор . 82 (4): 469–471. дои : 10.2307/1367580 . JSTOR 1367580 .
- ^ Перейти обратно: а б с Каннингем, Сьюзен Дж.; Элли, Морис Р.; Кастро, Изабель (январь 2011 г.). «Гистология и морфология лицевой щетины перьев у новозеландских птиц: влияние на функцию». Журнал морфологии (PDF) . 272 (1): 118–128. дои : 10.1002/jmor.10908 . ПМИД 21069752 . S2CID 20407444 .
- ^ Перейти обратно: а б Перринс, Кристофер М.; Аттенборо, Дэвид; Арлотт, Норман (1987). Путеводитель нового поколения по птицам Британии и Европы . Остин, Техас: Издательство Техасского университета. п. 205. ИСБН 978-0-292-75532-1 .
- ^ Кларк, Джордж А. младший (сентябрь 1961 г.). «Появление и время появления яичных зубов у птиц» (PDF) . Бюллетень Уилсона . 73 (3): 268–278.
- ^ Харрис, Тим, изд. (2009). National Geographic Полный список птиц мира . Вашингтон, округ Колумбия: National Geographic. п. 23. ISBN 978-1-4262-0403-6 .
- ^ Кайзер, Гэри В. (2007). Внутренняя птица: анатомия и эволюция . Ванкувер, Британская Колумбия: Вашингтонский университет Press. п. 26. ISBN 978-0-7748-1344-0 .
- ^ Ральф, Чарльз Л. (май 1969 г.). «Контроль цвета у птиц» . Американский зоолог . 9 (2): 521–530. дои : 10.1093/icb/9.2.521 . JSTOR 3881820 . ПМИД 5362278 .
- ^ Перейти обратно: а б с д
Хилл, Джеффри Э. (2010). Окраска птиц National Geographic . Вашингтон, округ Колумбия: National Geographic. стр. 62–66. ISBN 978-1-4262-0571-2 . - ^ Перейти обратно: а б с Роджерс, Лесли Дж.; Каплан, Гизела Т. (2000). Песни, рёв и ритуалы: Общение птиц, млекопитающих и других животных . Бостон, Массачусетс: Издательство Гарвардского университета. стр. 20, 83, 155. ISBN. 978-0-674-00827-4 .
- ^ Перейти обратно: а б с Жувентен, Пьер; Нолан, Пол М.; Орнборг, Йонас; Добсон, Ф. Стивен (февраль 2005 г.). «Ультрафиолетовые пятна у королевских и императорских пингвинов» . Кондор . 113 (3): 144–150. дои : 10.1650/7512 . S2CID 85776106 .
- ^ Кэмпбелл, Бернард Грант, изд. (1972). Половой отбор и происхождение человека: дарвиновский стержень . Нью-Брансуик, Нью-Джерси: Издатели транзакций. п. 186 . ISBN 978-0-202-02005-1 .
- ^ Томпсон, Билл; Блом, Эйрик А.Т.; Гордон, Джеффри А. (2005). Идентифицируйте себя: 50 наиболее распространенных проблем с идентификацией птиц . Нью-Йорк, штат Нью-Йорк: Houghton Mifflin Harcourt. п. 128 . ISBN 978-0-618-51469-4 .
- ^ О'Брайен, Майкл; Кроссли, Ричард; Карлсон, Кевин (2006). Путеводитель по прибрежным птицам . Нью-Йорк, штат Нью-Йорк: Хоутон Миффлин. п. 76. ИСБН 978-0-618-43294-3 .
- ^ Хауэлл (2007) п. 21
- ^ Паркс, AS; Эмменс, CW (1944). «Влияние андрогенов и эстрогенов на птиц». В Харрисе, Ричард С.; Тиманн, Кеннет Вивиан (ред.). Витамины и гормоны . Том. 2. Нью-Йорк, штат Нью-Йорк: Академик Пресс. п. 371. ИСБН 978-0-12-709802-9 .
- ^ Бхуллар, Бхарт-Анджан С.; Моррис, Закари С.; Сефтон, Элизабет М.; Ток, Аталай; Токита, Масаёси; Намкун, Бумджин; Камачо, Жасмин; Бернэм, Дэвид А.; Абжанов, Архат (июль 2015 г.). «Молекулярный механизм возникновения ключевой эволюционной инновации — птичьего клюва и неба, выявленный с помощью интегративного подхода к основным изменениям в истории позвоночных: МЕХАНИЗМ РАЗВИТИЯ ПРОИСХОЖДЕНИЯ ПТИЦЕВОГО КЛЮВА» . Эволюция . 69 (7): 1665–1677. дои : 10.1111/evo.12684 . ПМИД 25964090 . S2CID 205124061 .
- ^ Перейти обратно: а б Абжанов, Архат; Протас, Мередит; Грант, Б. Розмари; Грант, Питер Р.; Табин, Клиффорд Дж. (3 сентября 2004 г.). «Bmp4 и морфологическая изменчивость клювов у зябликов Дарвина» . Наука . 305 (5689): 1462–1465. Бибкод : 2004Sci...305.1462A . дои : 10.1126/science.1098095 . ISSN 0036-8075 . ПМИД 15353802 . S2CID 17226774 .
- ^ Перейти обратно: а б Абжанов, Архат; Куо, Уинстон П.; Хартманн, Кристина; Грант, Б. Розмари; Грант, Питер Р.; Табин, Клиффорд Дж. (август 2006 г.). «Путь кальмодулина и эволюция удлиненной морфологии клюва у зябликов Дарвина» . Природа . 442 (7102): 563–567. Бибкод : 2006Natur.442..563A . дои : 10.1038/nature04843 . ISSN 0028-0836 . ПМИД 16885984 . S2CID 2416057 .
- ^ Ито, Ёсихиро; Йе, Джэ Ён; Хитил, Анна; Хан, Джун; Брингас, Пабло; Накадзима, Акира; Шулер, Чарльз Ф.; Моисей, Гарольд Л.; Чай, Ян (1 ноября 2003 г.). «Условная инактивация Tgfbr2 в краниальном нервном гребне вызывает расщелину неба и дефекты черепа» . Разработка . 130 (21): 5269–5280. дои : 10.1242/dev.00708 . ISSN 1477-9129 . ПМИД 12975342 . S2CID 10925294 .
- ^ Малларино, Р.; Грант, PR; Грант, БР; Херрел, А.; Куо, В.П.; Абжанов, А. (08 марта 2011 г.). «Два модуля развития создают трехмерные вариации формы клюва дарвиновских вьюрков» . Труды Национальной академии наук . 108 (10): 4057–4062. Бибкод : 2011PNAS..108.4057M . дои : 10.1073/pnas.1011480108 . ISSN 0027-8424 . ПМК 3053969 . ПМИД 21368127 .
- ^ Самур (2000) с. 7
- ^ Патель, Мира (осень 2007 г.). «Электрорецепция Утконоса» . Биология 342: Поведение животных. Портленд, Орегон: Рид-Колледж .
- ^ «Утконос» . ЖиваяНаука . 4 августа 2014 г. 27572.
- ^ Перейти обратно: а б с д
Клейтон, [ неполная цитата ] ; Ли, [ неполная цитата ] ; Томпкинс, [ цитата неполная ] ; Броди, [ неполная цитата ] (сентябрь 1999 г.). «Взаимный естественный отбор по фенотипам хозяин-паразит» (PDF) . Американский натуралист . 154 (3): 261–270. дои : 10.1086/303237 . hdl : 10536/DRO/DU:30056229 . ISSN 1537-5323 . ПМИД 10506542 . S2CID 4369897 . Архивировано из оригинала (PDF) 12 сентября 2020 г. Проверено 5 сентября 2019 г.{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) [ нужна полная цитата ] - ^ Помрой, Делавэр (февраль 1962 г.). «Птицы с ненормальными клювами» (PDF) . Британские птицы . 55 (2): 49–72.
- ^
Бойд, [ неполная цитата ] (1951). «Обзор паразитизма Сталинга Sturnus vulgaris L. в Северной Америке». Журнал паразитологии . 37 (1): 56–84. дои : 10.2307/3273522 . JSTOR 3273522 . ПМИД 14825028 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) [ нужна полная цитата ] - ^
Стоит, [ неполная цитата ] (1940). «Записка о распространении маллофага ». Кольцевание птиц . 11:23 , 24.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) [ нужна полная цитата ] - ^
Эш, [ цитата неполная ] (1960). «Исследование маллофагов птиц с особым упором на их экологию». Ибис . 102 : 93–110. дои : 10.1111/j.1474-919X.1960.tb05095.x .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) CS1 maint: числовые имена: список авторов ( ссылка ) [ нужна полная цитата ] - ^ Клейтон, Дейл Х.; Мойер, Бретт Р.; Буш, Сара Э.; Джонс, Тони Г.; Гардинер, Дэвид В.; Роудс, Барри Б.; Голлер, Франц (22 апреля 2005 г.). «Адаптационное значение морфологии клюва птиц для борьбы с эктопаразитами» . Труды Лондонского королевского общества B: Биологические науки . 272 (1565): 811–817. дои : 10.1098/rspb.2004.3036 . ISSN 0962-8452 . ПМК 1599863 . ПМИД 15888414 .
- ^ Мойер, Бретт Р.; Петерсон, А. Таунсенд; Клейтон, Дейл Х. (2002). «Влияние формы клюва на нагрузку эктопаразитов у западных кустарниковых сойок» (PDF) . Кондор . 104 (3): 675–678. doi : 10.1650/0010-5422(2002)104[0675:iobsoe]2.0.co;2 . hdl : 1808/16618 . ISSN 0010-5422 . S2CID 32708877 .
- ^ Клейтон, Д.Х.; Вальтер, бакалавр (1 сентября 2001 г.). «Влияние экологии и морфологии хозяина на разнообразие неотропических птичьих вшей». Ойкос . 94 (3): 455–467. дои : 10.1034/j.1600-0706.2001.940308.x . ISSN 1600-0706 .
- ^ Таттерсолл, Гленн Дж.; Андраде, Денис В.; Абэ, Аугусто С. (24 июля 2009 г.). «Теплообмен тукана Билла свидетельствует о наличии управляемого сосудистого теплового излучателя». Наука . 325 (5949): 468–470. Бибкод : 2009Sci...325..468T . дои : 10.1126/science.1175553 . ПМИД 19628866 . S2CID 42756257 .
- ^ Гринберт, Рассел; Даннер, Раймонд; Олсен, Брайан; Лютер, Дэвид (14 июля 2011 г.). «Высокая летняя температура объясняет изменение размера клюва солончаковых воробьев». Экография . сначала онлайн (2): 146–152. дои : 10.1111/j.1600-0587.2011.07002.x .
- ^ Филлипс, Полли К.; Сэнборн, Аллен Ф. (декабрь 1994 г.). «Инфракрасное термографическое исследование температуры поверхности у трех бескилевых: страуса, эму и двубородого казуара». Журнал термической биологии . 19 (6): 423–430. дои : 10.1016/0306-4565(94)90042-6 .
- ^ «Эволюция птичьих счетов: птицы сокращают свои «счета за отопление» в холодном климате» . Наука Дейли . 23 июня 2010 г. Проверено 12 марта 2012 г.
- ^ Бирма, Натан (12 августа 2004 г.). «Добавьте это в список жизни: «Птицы» вдохновили на множество слов» . Чикаго Трибьюн . Проверено 6 июня 2011 г.
- ^ Террес, Джон К. (1980). Энциклопедия птиц Северной Америки Общества Одюбона Нью-Йорк, штат Нью-Йорк: Альфред А. Кнопф. ISBN 978-0-394-46651-4 .
- ^ Шрайбер, Элизабет Энн; Бургер, Джоанна, ред. (2002). Биология морских птиц . Бока-Ратон, Флорида: CRC Press. п. 325 . ISBN 978-0-8493-9882-7 .
- ^ Армстронг (1965) п. 7
- ^ Уилсон, Эдвард О. (1980). Социобиология . Бостон, Массачусетс: Издательство Гарвардского университета. п. 227. ИСБН 978-0-674-81624-4 .
- ^ Амерсон, А. Биньон (май 1967 г.). «Заболеваемость и перенос Rhinonyssidae (Acarina: Mesostigmata) у сажистых крачек ( Sterna fuscata )». Журнал медицинской энтомологии . 4 (2): 197–9. дои : 10.1093/jmedent/4.2.197 . ПМИД 6052126 .
- ^ Парк, Ф.Дж. (март 2011 г.). «Птичий трихомониаз: исследование поражений и относительной распространенности у различных видов птиц, живущих в неволе и на свободе, как это видно из австралийской птичьей практики». Журнал Австралийской ветеринарной ассоциации . 89 (3): 82–88. дои : 10.1111/j.1751-0813.2010.00681.x . ПМИД 21323655 .
- ^ Партридж, Эрик (2001). Непристойность Шекспира (4-е изд.). Лондон, Великобритания: Routledge Classics 2001. с. 82 . ISBN 978-0-415-25553-0 .
- ^ Бертон, Морис; Бертон, Роберт (1980). Международная энциклопедия дикой природы . Том. 12. Нью-Йорк, штат Нью-Йорк: Marshall Cavendish Corp., с. 1680.
- ^ Перейти обратно: а б Грандин, Темпл (2010). Улучшение благополучия животных: практический подход . Оксфордшир, Великобритания: CABI. п. 110. ИСБН 978-1-84593-541-2 .
- ^ Рэйс Фостер; Марти Смит. «Птичьи клювы: анатомия, уход и болезни» . Воспитание домашних животных . Архивировано из оригинала 4 июня 2012 года . Проверено 16 апреля 2012 г. [ самостоятельно опубликованный источник? ]
- ^ Эш, Лидия (2020) [2004]. «Справиться со своим Раптором» . Современный ученик . Архивировано из оригинала 6 апреля 2005 г. Проверено 16 апреля 2012 г.
- ^ Каннингем, Сьюзен Дж.; Элли, MR; Кастро, И.; Поттер, Массачусетс; Каннингем, М.; Пайн, MJ (2010). «Морфология клюва, или ибисы, предполагает наличие дистанционно-тактильной сенсорной системы для обнаружения добычи» . Аук . 127 (2): 308–316. дои : 10.1525/аук.2009.09117 . S2CID 85254980 .
- ^ Демери, Зои П.; Чаппелл, Дж.; Мартин, GR (2011). «Зрение, осязание и манипулирование объектами у сенегальских попугаев Poicephalus senegalus » . Труды Королевского общества Б. 278 (1725): 3687–3693. дои : 10.1098/rspb.2011.0374 . ПМК 3203496 . ПМИД 21525059 .
Библиография
[ редактировать ]- Армстронг, Эдвард Олверти (1965). Демонстрация и поведение птиц: введение в изучение психологии птиц . Нью-Йорк, штат Нью-Йорк: Dover Publications. LCCN 64013457 .
- Кэмпбелл, Брюс; Лэк, Элизабет, ред. (1985). Словарь птиц . Карлтон, Англия: Т. и А.Д. Пойзер. ISBN 978-0-85661-039-4 .
- Куэс, Эллиотт (1890). Справочник по полевой и общей орнитологии . Лондон, Великобритания: Macmillan and Co. p. 1 . OCLC 263166207 .
- Гилбертсон, Лэнс (1999). Руководство зоологической лаборатории (4-е изд.). Нью-Йорк, штат Нью-Йорк: Компании McGraw Hill. ISBN 978-0-07-237716-3 .
- Гилл, Фрэнк Б. (1995). Орнитология (2-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-2415-5 .
- Гирлинг, Саймон (2003). Ветеринарный уход за экзотическими домашними животными . Оксфорд, Великобритания: Blackwell Publishing. ISBN 978-1-4051-0747-1 .
- Хилл, Джеффри Э. (2010). Окраска птиц National Geographic . Вашингтон, округ Колумбия: National Geographic. ISBN 978-1-4262-0571-2 .
- Хауэлл, Стив Н.Г. (2007). Чайки Америки . Нью-Йорк, штат Нью-Йорк: Компания Houghton Mifflin. ISBN 978-0-618-72641-7 .
- Кинг, Энтони Стюарт; Маклелланд, Джон, ред. (1985). Форма и функции у птиц . Том. 3. Лондон, Великобритания: Академик Пресс. ISBN 978-0-12-407503-0 .
- Малларни, Киллиан; Свенссон, Ларс; Зеттерстрем, Дэн; Грант, Питер Дж. (1999). Путеводитель Коллинза по птицам: наиболее полный путеводитель по птицам Британии и Европы . Лондон, Великобритания: Харпер Коллинз. ISBN 978-0-00-711332-3 .
- Проктор, Ноубл С.; Линч, Патрик Дж. (1998). Руководство по орнитологии: строение и функции птиц . Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 978-0-300-07619-6 .
- Роджерс, Лесли Дж.; Каплан, Гизела Т. (2000). Песни, рёв и ритуалы: Общение птиц, млекопитающих и других животных . Бостон, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-00827-4 .
- Самур, Хайме, изд. (2000). Птичья медицина . Лондон, Великобритания: Мосби. ISBN 978-0-7234-2960-9 .
| Базы данных органов управления : Национальные |
|---|