Циккол
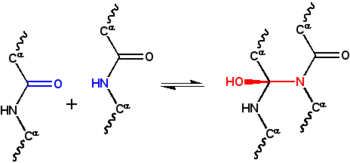
Гипотеза циклола дискредитированная первая структурная модель свернутого — ныне , глобулярного белка сформулированная в 1930-х годах. [ 1 ] Он был основан на циклольной реакции пептидных связей, предложенной физиком Фредериком Франком в 1936 году. [ 2 ] в котором две пептидные группы химически сшиты. Эти поперечные связи являются ковалентными аналогами нековалентных водородных связей между пептидными группами и наблюдались в редких случаях, например, в эргопептидах .
Основываясь на этой реакции, математик Дороти Ринч в конце 1930-х годов в серии из пяти статей выдвинула гипотезу о структурной модели глобулярных белков. Она предположила, что при некоторых условиях аминокислоты спонтанно образуют максимально возможное количество циклольных сшивок, в результате чего образуются молекулы циклола и ткани циклола . Далее она предположила, что глобулярные белки имеют третичную структуру, соответствующую платоновым телам и полуправильным многогранникам, образованным из циклольных тканей без свободных краев. В отличие от самой циклольной реакции, эти гипотетические молекулы, ткани и многогранники экспериментально не наблюдались. Модель имеет несколько последствий, которые делают ее энергетически неправдоподобной, например, стерические столкновения между боковыми цепями белка. В ответ на подобную критику Дж. Д. Бернал предположил, что гидрофобные взаимодействия главным образом ответственны за сворачивание белков . [ 3 ] что действительно подтвердилось.
Исторический контекст
[ редактировать ]К середине 1930-х годов аналитические исследования Теодора Сведберга с помощью ультрацентрифугирования показали, что белки имеют четко определенную химическую структуру и не представляют собой скопления небольших молекул. [ 4 ] Те же исследования показали, что молекулярная масса белков делится на несколько четко определенных классов, связанных целыми числами: [ 5 ] например M w = 2 п 3 д Da , где p и q — целые неотрицательные числа. [ 6 ] Однако определить точную молекулярную массу и количество аминокислот в белке было сложно. Сведберг также показал, что изменение условий растворения может привести к распаду белка на мелкие субъединицы, что теперь известно как изменение четвертичной структуры . [ 7 ]
все еще обсуждалась В то время химическая структура белков . [ 8 ] Наиболее распространенной (и в конечном итоге правильной) гипотезой было то, что белки представляют собой линейные полипептиды , т. е. неразветвленные полимеры аминокислот, связанных пептидными связями . [ 9 ] [ 10 ] Однако типичный белок удивительно длинный — сотни аминокислотных остатков — и некоторые выдающиеся ученые не были уверены, могут ли такие длинные линейные макромолекулы быть стабильными в растворе. [ 11 ] [ 12 ] Дальнейшие сомнения в полипептидной природе белков возникли из-за того, что некоторые ферменты расщепляли белки, но не пептиды, тогда как другие ферменты расщепляли пептиды, но не свернутые белки. [ 13 ] Попытки синтезировать белки в пробирке не увенчались успехом, главным образом из-за хиральности аминокислот; Белки природного происхождения состоят только из левосторонних аминокислот. Следовательно, были рассмотрены альтернативные химические модели белков, такие как дикетопиперазиновая гипотеза Эмиля Абдерхалдена . [ 14 ] [ 15 ] Однако ни одна альтернативная модель еще не объяснила, почему белки при гидролизе и протеолизе образуют только аминокислоты и пептиды. Как пояснил Линдерстрем-Ланг , [ 16 ] эти данные протеолиза показали, что денатурированные белки являются полипептидами, но данных о структуре свернутых белков еще не получено; таким образом, денатурация может включать химическое изменение, которое превращает свернутые белки в полипептиды.
Процесс денатурации белка (в отличие от коагуляции ) был открыт в 1910 году Харриетт Чик и Чарльзом Мартином . [ 17 ] но природа его все еще оставалась загадочной. Тим Энсон и Альфред Мирски показали, что денатурация — это обратимый процесс с двумя состояниями. [ 18 ] в результате многие химические группы становятся доступными для химических реакций, включая расщепление ферментами. [ 19 ] В 1929 году Сянь Ву правильно предположил, что денатурация соответствует разворачиванию белка, чисто конформационному изменению, которое приводит к воздействию растворителя на боковые цепи аминокислот. [ 20 ] Гипотеза Ву была также независимо выдвинута в 1936 году Мирским и Лайнусом Полингом . [ 21 ] Тем не менее ученые-белковики не могли исключить возможность того, что денатурация соответствует химическому изменению структуры белка. [ 19 ] гипотеза, которая считалась (отдаленной) возможностью до 1950-х годов. [ 22 ] [ 23 ]
Рентгеновская кристаллография как дисциплина возникла только в 1911 году и относительно быстро продвинулась от простых кристаллов соли к кристаллам сложных молекул, таких как холестерин . [ 24 ] Однако даже самые маленькие белки содержат более 1000 атомов, что значительно усложняет определение их структуры. [ 24 ] В 1934 году Дороти Кроуфут Ходжкин получила кристаллографические данные о структуре небольшого белка инсулина , хотя структура этого и других белков не была решена до конца 1960-х годов. [ 25 ] Однако первые рентгеновских лучей на волокнах данные по дифракции были собраны в начале 1930-х годов для многих природных волокнистых белков, таких как шерсть и волосы, Уильямом Эстбери , который предположил, что «глобулярные белки в целом могут складываться из элементов, по существу подобных элементам волокнистых белков». ." [ 26 ]
Поскольку в 1930-е годы структура белка была так плохо изучена, физические взаимодействия, ответственные за стабилизацию этой структуры, также были неизвестны. Эстбери предположил, что структура волокнистых белков стабилизируется за счет водородных связей в β-листах. [ 27 ] [ 28 ] Идею о том, что глобулярные белки также стабилизируются водородными связями, предложила Дороти Джордан Ллойд. [ 29 ] [ 30 ] в 1932 году, и позже его поддержали Альфред Мирский и Лайнус Полинг . [ 21 ] На лекции Эстбери в Оксфордском младшем научном обществе в 1933 году физик Фредерик Франк предположил, что волокнистый белок α-кератин может быть стабилизирован с помощью альтернативного механизма, а именно, ковалентного сшивания пептидных связей с помощью описанной выше реакции циклола. [ 31 ] Циклольная сшивка сближает две пептидные группы; Атомы N и C разделены ~1,5 Å они разделены ~3 Å , тогда как в типичной водородной связи . Идея заинтриговала Дж. Д. Бернала , который предложил ее математику Дороти Ринч как возможно полезную для понимания структуры белка. [ нужна ссылка ]
Основная теория
[ редактировать ]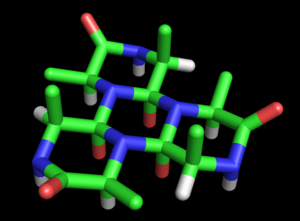
Ринч развил это предположение в полноценную модель структуры белка . Базовая модель циклола была изложена в ее первой статье (1936 г.). [ 32 ] Она отметила возможность того, что полипептиды могут циклизироваться с образованием замкнутых колец ( правда ) и что эти кольца могут образовывать внутренние поперечные связи посредством циклольной реакции (также верно, хотя и редко). Предполагая, что циклольная форма пептидной связи может быть более стабильной, чем амидная форма, Ринч пришел к выводу, что некоторые циклические пептиды естественным образом образуют максимальное количество циклольных связей (например, циклол 6 , рисунок 2). [ 32 ] Такие молекулы циклола имели бы гексагональную симметрию, если бы химические связи считались имеющими одинаковую длину, примерно 1,5 Å ; для сравнения: связи NC и CC имеют длину 1,42 Å и 1,54 Å соответственно. [ 32 ]
Эти кольца можно растягивать до бесконечности, образуя циклоловую ткань (рис. 3). [ 33 ] Такие ткани демонстрируют дальний квазикристаллический порядок, который, по мнению Ринча, скорее всего, был в белках, поскольку они должны плотно упаковывать сотни остатков. Еще одной интересной особенностью таких молекул и тканей является то, что их боковые цепи аминокислот направлены аксиально вверх только с одной стороны; противоположная грань не имеет боковых цепей. Таким образом, одна грань полностью не зависит от первичной последовательности пептида, что, по предположению Ринча, может объяснять независимые от последовательности свойства белков. [ 33 ]
В своей первоначальной статье Ринч ясно заявила, что модель циклола — это всего лишь рабочая гипотеза , потенциально действительная модель белков, которую необходимо будет проверить. [ 32 ] Целью ее этой статьи и ее последующих статей было предложить четко определенную проверяемую модель, проработать последствия ее предположений и сделать прогнозы, которые можно было бы проверить экспериментально. [ 34 ] В этих целях она преуспела; однако в течение нескольких лет эксперименты и дальнейшее моделирование показали, что гипотеза циклола несостоятельна в качестве модели глобулярных белков. [ 35 ] [ 36 ] [ 37 ]
Стабилизирующие энергии
[ редактировать ]
В двух тандемных письмах в редакцию (1936 г.) [ 38 ] [ 39 ] Ринч и Франк обратились к вопросу о том, действительно ли циклоловая форма пептидной группы более стабильна, чем амидная форма. Сравнительно простой расчет показал, что циклольная форма существенно менее стабильна, чем амидная. Следовательно, от циклольной модели придется отказаться, если не будет определен компенсирующий источник энергии. Первоначально Франк предположил, что форма циклола может быть стабилизирована за счет лучшего взаимодействия с окружающим растворителем; позже Ринч и Ирвинг Ленгмюр выдвинули гипотезу, что гидрофобная ассоциация неполярных боковых цепей обеспечивает стабилизирующую энергию для преодоления энергетических затрат циклоловых реакций. [ 40 ] [ 41 ]
Лабильность циклольной связи рассматривалась как преимущество модели, поскольку она давала естественное объяснение свойств денатурации ; возвращение циклольных связей в их более стабильную амидную форму открыло бы структуру и позволило бы атаковать эти связи протеазами , что согласуется с экспериментом. [ 42 ] [ 43 ] Ранние исследования показали, что белки, денатурированные под давлением, часто находятся в другом состоянии, чем те же белки, денатурированные под действием высокой температуры , что было интерпретировано как возможное подтверждение циклоловой модели денатурации. [ 44 ]
Гипотеза Ленгмюра-Ринча о гидрофобной стабилизации привела к падению циклоловой модели, главным образом из-за влияния Лайнуса Полинга , который поддерживал гипотезу о том, что структура белка стабилизируется за счет водородных связей . Прошло еще двадцать лет, прежде чем гидрофобные взаимодействия были признаны главной движущей силой сворачивания белков. [ 45 ]
Стерическая дополнительность
[ редактировать ]В своей третьей статье о циклолах (1936 г.) [ 46 ] Ринч отметил, что многие «физиологически активные» вещества, такие как стероиды, состоят из сросшихся шестиугольных колец атомов углерода и, таким образом, могут быть стерически комплементарны молекулам циклола без боковых цепей аминокислот . Ринч предположил, что стерическая комплементарность является одним из главных факторов, определяющих, будет ли небольшая молекула связываться с белком. [ нужна ссылка ]
Ринч предположил, что белки ответственны за синтез всех биологических молекул. Отметив, что клетки переваривают свои белки только в условиях крайнего голодания, Ринч далее предположил, что жизнь не может существовать без белков. [ нужна ссылка ]
Гибридные модели
[ редактировать ]С самого начала циклольную реакцию рассматривали как ковалентный аналог водородной связи . Поэтому было естественно рассмотреть гибридные модели с обоими типами связей. Это было предметом четвертой статьи Ринча о модели циклола (1936 г.). [ 47 ] написанная совместно с Дороти Джордан Ллойд , которая первой предположила, что глобулярные белки стабилизируются водородными связями. [ 29 ] В 1937 году была написана последующая статья, в которой упоминались другие исследователи водородных связей в белках, такие как Морис Лоял Хаггинс и Лайнус Полинг . [ 48 ]
Ринч также написал статью вместе с Уильямом Эстбери , отметив возможность кето-енольной изомеризации >C. а ЧАС а и амидно-карбонильная группа >C=O, образуя сшивку >C а -С(ОН а )< и снова превращая кислород в гидроксильную группу. [ 49 ] Такие реакции могут давать пятичленные кольца, тогда как классическая гипотеза циклола дает шестичленные кольца. Эта гипотеза сшивки кето-енолов не получила дальнейшего развития. [ 33 ]
Ткани, закрывающие пространство
[ редактировать ]
В своей пятой статье о циклолах (1937 г.) [ 50 ] Ринч определил условия, при которых две плоские ткани из циклола могут быть соединены так, чтобы между их плоскостями образовался угол, соблюдая при этом углы химической связи. Она выявила математическое упрощение, при котором неплоские шестичленные кольца атомов можно представить в виде плоских «срединных шестиугольников», образованных из средних точек химических связей. Это представление «среднего шестиугольника» позволило легко увидеть, что плоскости циклольной ткани могут быть правильно соединены, если двугранный угол между плоскостями равен тетраэдрическому углу связи δ = arccos (-1/3) ≈ 109,47 °. [ нужна ссылка ]
Можно построить большое разнообразие замкнутых многогранников, отвечающих этому критерию, из которых простейшими являются усеченный тетраэдр , усеченный октаэдр и октаэдр , которые представляют собой платоновы тела или полуправильные многогранники . Рассматривая первую серию «замкнутых циклолов» (моделированных по усеченному тетраэдру), Ринч показал, что число аминокислот в них увеличивается квадратично как 72 н. 2 , где n – номер замкнутого циклола C n . Так, циклол С1 циклол С2 имеет 72 остатка, — 288 остатков и т. д. Предварительное экспериментальное подтверждение этого предсказания пришло от Макса Бергмана и Карла Нимана , [ 6 ] чей аминокислотный анализ показал, что белки состоят из целых чисел, кратных 288 аминокислотным остаткам ( n =2). В более общем смысле, циклоловая модель глобулярных белков объясняет ранние ультрацентрифугирования аналитические результаты Теодора Сведберга , которые предполагали, что молекулярные массы белков делятся на несколько классов, связанных целыми числами. [ 4 ] [ 5 ]
Модель циклола соответствовала общим свойствам, приписываемым тогда свернутым белкам. [ 51 ] (1) Исследования центрифугирования показали, что свернутые белки значительно плотнее воды (~1,4 г / мл ) и, следовательно, плотно упакованы; Ринч предположил, что плотная упаковка должна подразумевать регулярную упаковку. (2) Несмотря на свой большой размер, некоторые белки легко кристаллизуются в симметричные кристаллы, что соответствует идее о симметричных гранях, которые совпадают при ассоциации. (3) Белки связывают ионы металлов; поскольку места связывания металлов должны иметь специфическую геометрию связей (например, октаэдрическую), было правдоподобно предположить, что весь белок также имеет аналогичную кристаллическую геометрию. (4) Как описано выше, модель циклола дала простое химическое объяснение денатурации и трудности расщепления свернутых белков протеазами. (5) Считалось, что белки ответственны за синтез всех биологических молекул, включая другие белки. Ринч отметил, что фиксированная, однородная структура была бы полезна для белков при планировании их собственного синтеза, аналогично концепции Уотсона - Фрэнсиса Крика о ДНК, моделирующей собственную репликацию. Учитывая, что многие биологические молекулы, такие как сахара и стерины имеют гексагональную структуру, можно было предположить, что синтезирующие их белки также имеют гексагональную структуру. Ринч обобщила свою модель и подтверждающие экспериментальные данные по молекулярной массе в трех обзорных статьях. [ 52 ]
Предсказанные белковые структуры
[ редактировать ]Предложив модель глобулярных белков, Ринч исследовал, согласуется ли она с имеющимися структурными данными. Она предположила, что бычий туберкулиновый белок (523) представляет собой замкнутый циклол C 1 , состоящий из 72 остатков. [ 53 ] и что пищеварительный фермент пепсин представляет собой замкнутый C 2 циклол из 288 остатков. [ 54 ] [ 55 ] Эти предсказания количества остатков было трудно проверить, поскольку доступные тогда методы измерения массы белков, такие как аналитическое ультрацентрифугирование и химические методы, были неточными. [ нужна ссылка ]
Ринч также предсказал, что инсулин представляет собой замкнутый С2 - циклол, состоящий из 288 остатков. Для инсулина были доступны ограниченные рентгеновские кристаллографические данные, которые Ринч интерпретировала как «подтверждающие» ее модель. [ 56 ] Однако такая интерпретация вызвала довольно резкую критику как преждевременная. [ 57 ] Тщательное изучение диаграмм инсулина Паттерсона, полученных Дороти Кроуфут-Ходжкин, показало, что они примерно соответствуют модели циклола; однако соглашение не было достаточно хорошим, чтобы утверждать, что модель циклола подтвердилась. [ 58 ]
Неправдоподобность модели
[ редактировать ]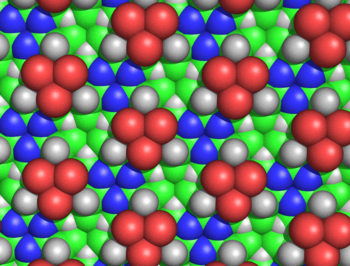
Было показано, что циклоловая ткань неправдоподобна по нескольким причинам. Ганс Нейрат и Генри Булл показали, что плотная упаковка боковых цепей в ткани циклола несовместима с экспериментальной плотностью, наблюдаемой в белковых пленках. [ 59 ] Морис Хаггинс подсчитал, что несколько несвязанных атомов циклольной ткани будут сближаться ближе, чем позволяют их ван-дер-ваальсовые радиусы ; например, внутренний H а и С а атомы лакун будут разделены всего 1,68 Å (рис. 5). [ 35 ] Хауровиц химически показал, что внешняя часть белков не может иметь большого количества гидроксильных групп, что является ключевым предсказанием циклоловой модели. [ 60 ] тогда как Мейер и Хоэнемсер показали, что циклольные конденсации аминокислот не существуют даже в ничтожных количествах в качестве переходного состояния . [ 61 ] Более общие химические аргументы против модели циклола были приведены Бергманном и Ниманном. [ 62 ] и Нойбергера . [ 36 ] [ 37 ] Данные инфракрасной спектроскопии показали, что число карбонильных групп в белке при гидролизе не меняется. [ 63 ] и что интактные, свернутые белки имеют полный набор амидных карбонильных групп; [ 64 ] оба наблюдения противоречат гипотезе циклола о том, что такие карбонилы превращаются в гидроксильные группы в свернутых белках. Наконец, известно, что белки содержат пролин в значительных количествах (обычно 5%); поскольку у пролина отсутствует амидный водород, а его азот уже образует три ковалентные связи, пролин кажется неспособным к циклольной реакции и включению в циклольную ткань. Энциклопедическое резюме химических и структурных доказательств против модели циклола было дано Полингом и Ниманном. [ 65 ] Более того, есть подтверждающее доказательство — результат того, что все белки содержат целое число аминокислотных остатков , кратное 288. [ 6 ] — также было показано, что оно неверно в 1939 году. [ 66 ]
Ринч ответил на критику циклоловой модели в отношении стерического столкновения, свободной энергии, химического состава и количества остатков. Что касается стерических столкновений, она отметила, что небольшие деформации валентных углов и длин связей позволили бы облегчить эти стерические столкновения или, по крайней мере, уменьшить их до разумного уровня. [ 67 ] Она отметила, что расстояния между несвязанными группами внутри одной молекулы могут быть короче, чем можно было бы ожидать, исходя из их радиусов Ван-дер-Ваальса , например, расстояние 2,93 Å между метильными группами в гексаметилбензоле. Что касается штрафа за свободную энергию для реакции циклола, Ринч не согласился с расчетами Полинга и заявил, что о внутримолекулярных энергиях известно слишком мало, чтобы исключить модель циклола только на этом основании. [ 67 ] В ответ на химическую критику Ринч предположил, что изученные модельные соединения и простые бимолекулярные реакции не обязательно относятся к модели циклола и что стерические препятствия могли препятствовать реакции поверхностных гидроксильных групп. [ 34 ] Что касается критики числа остатков, Ринч расширила свою модель, включив в нее другое количество остатков. В частности, она произвела «минимальный» закрытый циклол, состоящий всего из 48 остатков. [ 68 ] и на этом (неверном) основании он, возможно, был первым, кто предположил, что мономер инсулина имел молекулярную массу примерно 6000 Да . [ 69 ] [ 70 ]
Поэтому она утверждала, что циклоловая модель глобулярных белков все еще потенциально жизнеспособна. [ 71 ] [ 72 ] и даже предложил циклоловую ткань как компонент цитоскелета . [ 73 ] Однако большинство ученых-белковиков перестали в это верить, и Ринч обратила свое научное внимание на математические проблемы рентгеновской кристаллографии , в решение которых она внесла значительный вклад. [ 74 ] Единственным исключением была физик Глэдис Анслоу , коллега Ринч в Смит-колледже , которая изучала ультрафиолетовые спектры поглощения белков и пептидов в 1940-х годах и допустила возможность использования циклолов при интерпретации своих результатов. [ 75 ] [ 76 ] Когда последовательность введения инсулина начал определять Фредерик Сэнгер , [ 25 ] Анслоу опубликовал трехмерную модель циклола с боковыми цепями. [ 77 ] основан на основе модели «минимального циклола» Ринча 1948 года. [ 68 ]
Частичное погашение
[ редактировать ]
Крах общей модели циклола в целом привел к отказу от ее элементов; Одним из заметных исключений было Дж. Д. Берналом недолгое принятие гипотезы Ленгмюра-Ринча о том, что сворачивание белков обусловлено гидрофобной ассоциацией. [ 79 ] Тем не менее, циклольные связи были идентифицированы в небольших природных циклических пептидах в 1950-х годах. [ нужна ссылка ]
Уточнение современной терминологии является целесообразным. Классическая циклольная реакция представляет собой присоединение NH-амина одной пептидной группы к карбонильной группе C=O другой; полученное соединение теперь называется азациклолом . По аналогии, оксациклол образуется при присоединении гидроксильной группы OH к пептидильной карбонильной группе. Аналогичным образом тиациклол образуется путем добавления тиольного фрагмента SH к пептидильной карбонильной группе. [ 80 ]
оксациклола Алкалоид эрготамин из гриба Claviceps purpurea был первым идентифицированным циклолом. [ 81 ] Циклический депсипептид серратамолид также образуется в результате оксациклоловой реакции. [ 82 ] Получены также химически аналогичные циклические тиациклолы. [ 83 ] Классические азациклолы наблюдались в небольших молекулах. [ 84 ] и трипептиды. [ 85 ] Пептиды естественным образом производятся путем реверсии азацилолов. [ 86 ] ключевое предсказание циклоловой модели. В настоящее время идентифицированы сотни молекул циклола, несмотря на расчет Лайнуса Полинга , согласно которому такие молекулы не должны существовать из-за их неблагоприятно высокой энергии . [ 65 ]
После долгого перерыва, во время которого она работала в основном над математикой рентгеновской кристаллографии , Ринч отреагировала на эти открытия с новым энтузиазмом по поводу модели циклола и ее значимости в биохимии. [ 87 ] Она также опубликовала две книги, описывающие теорию циклола и малые пептиды в целом. [ 88 ] [ 89 ]
Ссылки
[ редактировать ]- ^ Тиселиус А (1939). «Химия белков и аминокислот». Ежегодный обзор биохимии . 8 : 155–184. дои : 10.1146/annurev.bi.08.070139.001103 .
- ^ Франк, ФК (1936). «Энергия образования молекул циклола» . Природа . 138 (3484): 242. Бибкод : 1936Natur.138..242F . дои : 10.1038/138242a0 . S2CID 4065283 .
- ^ Бернал Дж.Д. (1939). «Строение белков». Природа . 143 (3625): 663–667. Бибкод : 1939Natur.143..663B . дои : 10.1038/143663a0 . S2CID 46327591 .
- ^ Jump up to: а б Сведберг Т (1929). «Масса и размеры белковых молекул» . Природа . 123 (3110): 871. Бибкод : 1929Natur.123..871S . дои : 10.1038/123871a0 . S2CID 4068088 .
- ^ Jump up to: а б Сведберг Т (1934). «Седиментация молекул в центробежных полях». Химические обзоры . 14 : 1–15. дои : 10.1021/cr60047a001 .
- ^ Jump up to: а б с Бергманн М, Ниманн С (1937). «О строении белков: гемоглобин крупного рогатого скота, яичный альбумин, фибрин крупного рогатого скота и желатин» . Журнал биологической химии . 118 : 301–314. дои : 10.1016/S0021-9258(18)74540-7 .
- ^ Сведберг Т (1930). «Области стабильности pH белков». Труды Фарадеевского общества . 26 : 741–744. дои : 10.1039/TF9302600737 .
- ^ Фрутон Дж.С. (май 1979 г.). «Ранние теории структуры белка». Анналы Нью-Йоркской академии наук . 325 (1): xiv, 1-xiv, 18. Бибкод : 1979NYASA.325....1F . дои : 10.1111/j.1749-6632.1979.tb14125.x . ПМИД 378063 . S2CID 39125170 .
- ^ Хофмейстер Ф (1902). «О строении и группировке белковых тел». Результаты физиологии . 1 :759-802. дои : 10.1007/BF02323641 . S2CID 101988911 .
- ^ Фишер Х.Э. (1902). «О гидролизе белковых веществ». Газета «Химик» . 26 :939-940.
- ^ Фишер Х.Э. (1913). «Синтез депсидов, лишайников и дубильных веществ» . Отчеты Немецкого химического общества . 46 (3): 3253–3289. дои : 10.1002/cber.191304603109 .
- ^ Соренсен С.П. (1930). «Состав растворимых белков как обратимо диссоциируемых компонентных систем». Отчеты о работе лаборатории Carlsberg . 18 :1–124.
- ^ Фрутон Дж.С. (1999). Белки, ферменты, гены: взаимодействие химии и биологии . Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 0-585-35980-6 .
- ^ Абдерхальден Э (1924). «Дикетопиперазины». естественные науки . 12 (36): 716–720. Бибкод : 1924NW.....12..716A . дои : 10.1007/BF01504819 . S2CID 29012795 .
- ^ Абдерхальден Э , Комм Э (1924). «Об ангидридном строении белков». Журнал физиологической химии . 139 (3–4): 181–204. дои : 10.1515/bchm2.1924.139.3-4.181 .
- ^ Линдерстрем-Ланг К. , Хочкисс Р.Д., Йохансен Г. (1938). «Пептидные связи в глобулярных белках» . Природа . 142 (3605): 996. Бибкод : 1938Natur.142..996L . дои : 10.1038/142996a0 . S2CID 4086716 .
- ^ Чик Х. (июль 1910 г.). «О «тепловой коагуляции» белков» . Журнал физиологии . 40 (5): 404–430. дои : 10.1113/jphysicalol.1910.sp001378 . ПМК 1533708 . ПМИД 16993016 .
Чик Х (сентябрь 1911 г.). «О «тепловой коагуляции» белков: Часть II. Действие горячей воды на яичный белок и влияние кислоты и солей на скорость реакции» . Журнал физиологии . 43 (1): 1–27. дои : 10.1113/jphysicalol.1911.sp001456 . ПМЦ 1512746 . ПМИД 16993081 .
Чик Х. (август 1912 г.). «О «тепловой коагуляции» белков: Часть III. Влияние щелочи на скорость реакции» . Журнал физиологии . 45 (1–2): 61–69. дои : 10.1113/jphysicalol.1912.sp001535 . ПМК 1512881 . ПМИД 16993182 .
Чик Х. (октябрь 1912 г.). «О «тепловой коагуляции» белков: Часть IV. Условия, контролирующие агглютинацию белков, уже подвергнутых воздействию горячей воды» . Журнал физиологии . 45 (4): 261–295. дои : 10.1113/jphysicalol.1912.sp001551 . ПМЦ 1512885 . ПМИД 16993156 . - ^ Ансон М.Л. , Мирский А.Е. (ноябрь 1929 г.). «Получение полностью коагулированного гемоглобина» . Журнал общей физиологии . 13 (2): 121–132. дои : 10.1085/jgp.13.2.121 . ПМК 2141032 . ПМИД 19872511 .
- ^ Jump up to: а б Энсон М.Л. (1945). «Денатурация белка и свойства белковых групп». Достижения в области химии белков . 2 : 361–386. дои : 10.1016/S0065-3233(08)60629-4 . ISBN 978-0-12-034202-0 .
- ^ У Х (1931). «Исследования по денатурации белков. XIII. Теория денатурации». Китайский журнал физиологии . 5 : 321–344. Предварительные отчеты были представлены перед XIII Международным конгрессом физиологов в Бостоне (19–24 августа 1929 г.) и в октябрьском номере « Американского журнала физиологии» за 1929 г.
- ^ Jump up to: а б Мирский А.Е. , Полинг Л. (июль 1936 г.). «О строении нативных, денатурированных и коагулированных белков» . Труды Национальной академии наук Соединенных Штатов Америки . 22 (7): 439–447. Бибкод : 1936ПНАС...22..439М . дои : 10.1073/pnas.22.7.439 . ПМЦ 1076802 . ПМИД 16577722 .
- ^ Нейрат Х. , Гринштейн Дж. П., Патнэм Ф. В., Эриксон Дж. О. (1944). «Химия денатурации белка». Химические обзоры . 34 (2): 157–265. дои : 10.1021/cr60108a003 .
- ^ Патнэм Ф (1953). «Денатурация белка». В Нейрате Х., Бейли К. (ред.). Белки . Том. 1Б. стр. 807–892.
- ^ Jump up to: а б Джерузалми Д (2007). «Первый анализ макромолекулярных кристаллов: биохимия и дифракция рентгеновских лучей». В Дубле S (ред.). Протоколы макромолекулярной кристаллографии, Том 2 . Методы молекулярной биологии. Том. 364. Клифтон, Нью-Джерси, стр. 43–62. дои : 10.1385/1-59745-266-1:43 . ISBN 978-1-59745-266-3 . ПМИД 17172760 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Jump up to: а б Сэнгер Ф., Таппи Х. (сентябрь 1951 г.). «Аминокислотная последовательность в фенилаланильной цепи инсулина. I. Идентификация низших пептидов из частичных гидролизатов» . Биохимический журнал . 49 (4): 463–481. дои : 10.1042/bj0490463 . ПМК 1197535 . ПМИД 14886310 . ; Сэнгер Ф., Таппи Х. (сентябрь 1951 г.). «Аминокислотная последовательность фенилаланильной цепи инсулина. 2. Исследование пептидов из ферментных гидролизатов» . Биохимический журнал . 49 (4): 481–490. дои : 10.1042/bj0490481 . ПМК 1197536 . ПМИД 14886311 . ; Сэнгер Ф., Томпсон Э.О. (февраль 1953 г.). «Аминокислотная последовательность в глицильной цепи инсулина. I. Идентификация низших пептидов из частичных гидролизатов» . Биохимический журнал . 53 (3): 353–366. дои : 10.1042/bj0530353 . ПМК 1198157 . ПМИД 13032078 . ; Сэнгер Ф., Томпсон Э.О. (февраль 1953 г.). «Аминокислотная последовательность в глицильной цепи инсулина. II. Исследование пептидов из ферментных гидролизатов» . Биохимический журнал . 53 (3): 366–374. дои : 10.1042/bj0530366 . ПМК 1198158 . ПМИД 13032079 .
- ^ Чарльз Танфорд и Жаклин Рейнольдс (2001) Роботы природы: история белков , страницы с 80 по 83, Oxford University Press ISBN 019 850466 7
- ^ Эстбери WT , Вудс HJ (1931). «Молекулярная масса белков». Природа . 127 (3209): 663–665. Бибкод : 1931Natur.127..663A . дои : 10.1038/127663b0 . S2CID 4133226 .
- ^ Эстбери WT (1933). «Некоторые проблемы рентгеновского анализа структуры волос животных и других белковых волокон». Труды Фарадеевского общества . 29 (140): 193–211. дои : 10.1039/tf9332900193 .
- ^ Jump up to: а б Джордан Ллойд Д. (1932). «Коллоидная структура и ее биологическое значение». Биологические обзоры . 7 (3): 254–273. дои : 10.1111/j.1469-185x.1962.tb01043.x . S2CID 84286671 .
- ^ Ллойд DJ , Marriott RH (1933). «Название неизвестно». Труды Фарадеевского общества . 29 : 1228. дои : 10.1039/tf9332901228 .
- ^ Эстбери WT (1936). «Неизвестное название». Журнал Текстильного института . 27 : 282–?.
- ^ Jump up to: а б с д Ринч ДМ (1936). «Узор белков». Природа . 137 (3462): 411–412. Бибкод : 1936Natur.137..411W . дои : 10.1038/137411a0 . S2CID 4140591 .
- ^ Jump up to: а б с Ключ ДМ . «Тканевая теория структуры белка». Философский журнал . 30 : 64–67.
- ^ Jump up to: а б Ринч ДМ (1940). «Гипотеза циклола». Природа . 145 (3678): 669–670. Бибкод : 1940Natur.145..669W . дои : 10.1038/145669a0 . S2CID 4065545 .
- ^ Jump up to: а б Хаггинс М. (1939). «Строение белков». Журнал Американского химического общества . 61 (3): 755. дои : 10.1021/ja01872a512 .
- ^ Jump up to: а б Нойбергер А (1939). «Химическая критика циклоловой и частотной гипотезы структуры белка». Труды Королевского общества . 170 : 64–65.
- ^ Jump up to: а б Нойбергер А (1939). «Химические аспекты гипотезы циклола». Природа . 143 (3620): 473. Бибкод : 1939Natur.143..473N . дои : 10.1038/143473a0 . S2CID 4102966 .
- ^ Ринч ДМ (1936). «Энергия образования молекул циклола». Природа . 138 (3484): 241–242. Бибкод : 1936Natur.138..241W . дои : 10.1038/138241a0 . S2CID 4103892 .
- ^ Франк ФК (1936). «Энергия образования молекул циклола» . Природа . 138 (3484): 242. Бибкод : 1936Natur.138..242F . дои : 10.1038/138242a0 . S2CID 4065283 .
- ^ Ленгмюр I , Ринч Д.М. (1939). «Природа циклоловой связи». Природа . 143 (3611): 49–52. Бибкод : 1939Natur.143...49L . дои : 10.1038/143049a0 . S2CID 4056966 .
- ^ Ленгмюр I (1939). «Строение белков». Труды Физического общества . 51 (4): 592–612. Бибкод : 1939PPS....51..592L . дои : 10.1088/0959-5309/51/4/305 .
- ^ Ринч ДМ (1938). «О гидратации и денатурации белков». Философский журнал . 25 : 705–739.
- ^ Ринч ДМ (1936). «Гидратация и денатурация белков» . Природа . 142 (3588): 260. Бибкод : 1938Natur.142..259. . дои : 10.1038/142259a0 .
- ^ Доу Р.Б., Мэтьюз-младший Дж.Э., Торп В.Т. (1940). «Влияние обработки высоким давлением на физиологическую активность инсулина». Американский журнал физиологии . 131 (2): 382–387. дои : 10.1152/ajplegacy.1940.131.2.382 .
- ^ Каузманн В. (1959). «Некоторые факторы интерпретации денатурации белка». Достижения в области химии белков . 14 :1–63. дои : 10.1016/S0065-3233(08)60608-7 . ISBN 978-0-12-034214-3 . ПМИД 14404936 .
- ^ Ринч ДМ (1936). «Строение белков и некоторых физиологически активных соединений» . Природа . 138 (3493): 651–652. Бибкод : 1936Natur.138..651W . дои : 10.1038/138651a0 . S2CID 4108696 .
- ^ Ринч Д.М. , Джордан Ллойд Д. (1936). «Водородная связь и структура белков». Природа . 138 (3496): 758–759. Бибкод : 1936Natur.138..758W . дои : 10.1038/138758a0 . S2CID 4096438 .
- ^ Ринч ДМ (1937). «Природа связи в белках» . Природа . 139 (3521): 718. Бибкод : 1937Natur.139..718W . дои : 10.1038/139718a0 . S2CID 4116000 .
- ^ Эстбери В.Т. , Ринч Д.М. (1937). «Внутримолекулярное сворачивание белков путем кето-енолового обмена» . Природа . 139 (3523): 798. Бибкод : 1937Natur.139..798A . дои : 10.1038/139798a0 . S2CID 41311699 .
- ^ Ринч ДМ (1937). «Теория циклола и «глобулярные» белки» . Природа . 139 (3527): 972–973. Бибкод : 1937Natur.139..972W . дои : 10.1038/139972a0 . S2CID 4066210 .
- ^ Ринч Д. (июль 1947 г.). «Нативный белок». Наука . 106 (2743): 73–76. Бибкод : 1947Sci...106...73W . дои : 10.1126/science.106.2743.73 . ПМИД 17808858 .
- ^ Ринч ДМ (1937). «О структуре белков». Труды Королевского общества . А160 : 59–86.
Ринч ДМ (1937). «Гипотеза циклола и «глобулярные» белки». Труды Королевского общества . А161 : 505–524.
Ринч ДМ (1938). «О молекулярной массе глобулярных белков». Философский журнал . 26 : 313–332. - ^ Ринч ДМ (1939). «Туберкулиновый белок TBU-бычий (523)» . Природа . 144 (3636): 77. Бибкод : 1939Natur.144...77W . дои : 10.1038/144077a0 . S2CID 4113033 .
- ^ Ринч ДМ (1937). «О строении пепсина». Философский журнал . 24 :940.
- ^ Ринч ДМ (1938). «Структура пепсина» . Природа . 142 (3587): 217. Бибкод : 1938Natur.142..215. . дои : 10.1038/142215a0 .
- ^ Ринч ДМ (июнь 1937 г.). «О строении инсулина». Наука . 85 (2215): 566–567. Бибкод : 1937Sci....85..566W . дои : 10.1126/science.85.2215.566 . ПМИД 17769864 .
Ринч ДМ (1937). «О строении инсулина». Труды Фарадеевского общества . 33 : 1368–1380. дои : 10.1039/tf9373301368 .
Ринч ДМ (1938). «Строение молекулы инсулина». Журнал Американского химического общества . 60 (8): 2005–2006. дои : 10.1021/ja01275a514 . S2CID 45312422 .
Ринч ДМ (август 1938 г.). «Строение молекулы инсулина». Наука . 88 (2276): 148–149. Бибкод : 1938Sci....88..148W . дои : 10.1126/science.88.2276.148-a . ПМИД 17751525 .
Ринч Д.М. , Ленгмюр I (1938). «Строение молекулы инсулина». Журнал Американского химического общества . 60 (9): 2247–2255. дои : 10.1021/ja01276a062 .
Ленгмюр I , Ринч Д.М. (1939). «Заметка о структуре инсулина». Труды Физического общества . 51 (4): 613–624. Бибкод : 1939PPS....51..613L . дои : 10.1088/0959-5309/51/4/306 . - ^ Брэгг В.Л. (1939). «Диаграммы Паттерсона в кристаллическом анализе». Природа . 143 (3611): 73–74. Бибкод : 1939Natur.143...73B . дои : 10.1038/143073a0 . S2CID 4063833 .
Бернал Дж.Д. (1939). «Векторные карты и гипотеза циклола». Природа . 143 (3611): 74–75. Бибкод : 1939Natur.143...74B . дои : 10.1038/143074a0 . S2CID 4108005 .
Робертсон Дж. М. (1939). «Векторные карты и тяжелые атомы в кристаллическом анализе и структуре инсулина». Природа . 143 (3611): 75–76. Бибкод : 1939Natur.143...75R . дои : 10.1038/143075a0 . S2CID 4053119 . - ^ Райли Д. П., Фанкучен I (1939). «Производный анализ по Паттерсону скелета молекулы циклола C 2 ». Природа . 143 (3624): 648–649. Бибкод : 1939Natur.143..648R . дои : 10.1038/143648a0 . S2CID 4086672 .
Ринч ДМ (1940). «Проекция Паттерсона скелетов структуры, предложенной для молекулы инсулина» . Природа . 145 (3687): 1018. Бибкод : 1940Natur.145.1018W . дои : 10.1038/1451018a0 . S2CID 4119581 .
Райли Д. (1940). «Анализ Паттерсона, полученный на основе скелета циклола C 2 » . Природа . 146 (3694): 231. Бибкод : 1940Natur.146..231R . дои : 10.1038/146231a0 . S2CID 4111977 . - ^ Нейрат Х. , Булл Х.Б. (1938). «Поверхностная активность белков». Химические обзоры . 23 (3): 391–435. дои : 10.1021/cr60076a001 .
- ^ Хауровиц Ф (1938). «Расположение пептидных цепей в молекулах сферопротеинов». Журнал физиологической химии Хоппе-Зейлера . 256 :28–32. дои : 10.1515/bchm2.1938.256.1.28 .
- ^ Мейер К.Х., Хоэнемсер В. (1938). «Возможность образования циклолов из простых пептидов». Природа . 141 (3582): 1138–1139. Бибкод : 1938Natur.141.1138M . дои : 10.1038/1411138b0 . S2CID 4097115 .
- ^ Бергманн М, Ниманн С (1938). «Химия аминокислот и белков» . Ежегодный обзор биохимии . 7 (2): 99–124. дои : 10.1146/annurev.bi.07.070138.000531 . ПМК 537431 .
- ^ Хауровиц Ф., Аструп Т. (1939). «Ультрафиолетовое поглощение настоящего и гидролизованного белка». Природа . 143 (3612): 118–119. Бибкод : 1939Natur.143..118H . дои : 10.1038/143118b0 . S2CID 4078416 .
- ^ Клотц И.М., Грисволд П. (март 1949 г.). «Инфракрасные спектры и амидная связь в нативном глобулярном белке». Наука . 109 (2830): 309–310. Бибкод : 1949Sci...109..309K . дои : 10.1126/science.109.2830.309 . ПМИД 17782718 .
- ^ Jump up to: а б Полинг Л. , Ниманн К. (1939). «Строение белков». Журнал Американского химического общества . 61 (7): 1860–1867. дои : 10.1021/ja01876a065 .
- ^ Гочкис Р.Д. (1939). «Определение пептидных связей в кристаллическом лактоглобулине» . Журнал биологической химии . 131 : 387–395. дои : 10.1016/S0021-9258(18)73511-4 .
- ^ Jump up to: а б Ринч ДМ (1941). «Геометрическая атака на структуру белка». Журнал Американского химического общества . 63 (2): 330–33. дои : 10.1021/ja01847a004 .
- ^ Jump up to: а б Ринч Д. (апрель 1948 г.). «Нативные белки как поликонденсации аминокислот». Наука . 107 (2783): 445–446. Бибкод : 1948Sci...107R.445W . дои : 10.1126/science.107.2783.445-a . ПМИД 17844448 . S2CID 10206302 .
- ^ Ринч Д. (март 1952 г.). «Скелетные единицы в белковых кристаллах». Наука . 115 (2987): 356–357. Бибкод : 1952Sci...115..356W . дои : 10.1126/science.115.2987.356 . ПМИД 17748855 .
- ^ Ринч Д. (ноябрь 1952 г.). «Молекулы строения инсулина». Наука . 116 (3021): 562–564. Бибкод : 1952Sci...116..562W . дои : 10.1126/science.116.3021.562 . ПМИД 13015111 .
- ^ Ринч ДМ (1939). «Строение глобулярных белков». Природа . 143 (3620): 482–483. Бибкод : 1939Natur.143..482W . дои : 10.1038/143482a0 . S2CID 5362977 .
- ^ Ринч ДМ (1939). «Теория циклола и структура инсулина». Природа . 143 (3627): 763–764. Бибкод : 1939Natur.143..763W . дои : 10.1038/143763a0 . S2CID 4063795 .
- ^ Ринч ДМ (1939). «Нативные белки, гибкие каркасы и цитоплазматическая организация». Природа . 150 (3800): 270–271. Бибкод : 1942Natur.150..270W . дои : 10.1038/150270a0 . S2CID 4085657 .
- ^ Сенешаль М (2012). Я умер за красоту: Дороти Ринч и научная культура . Оксфорд: Издательство Оксфордского университета, США. ISBN 978-0-19-987579-5 . OCLC 818851574 .
- ^ Анслоу Г.А. (1942). «Энергия связи в некоторых белковых тканях и боковых цепях». Физический обзор . 61 (7–8): 547. Бибкод : 1942PhRv...61..541. . дои : 10.1103/PhysRev.61.541 .
- ^ Анслоу Г.А. (1945). «Ультрафиолетовые спектры биологически важных молекул». Журнал прикладной физики . 16 (1): 41–49. Бибкод : 1945JAP....16...41A . дои : 10.1063/1.1707499 .
- ^ Анслоу Г.А. (1953). «Участки аминокислотных остатков на циклоловой модели инсулина» . Журнал химической физики . 21 (11): 2083–2084. Бибкод : 1953ЖЧФ..21.2083А . дои : 10.1063/1.1698765 .
- ^ Гедес Т., Нуньес А., Тинео Э., Нуньес О. (2002). «Эффект конфигурации размера кольца и трансаннулярные внутренние скорости в макроциклах бислактама». Журнал Химического общества, Perkin Transactions 2 . 2002 (12): 2078–2082. дои : 10.1039/b207233e .
- ^ Бернал Дж.Д. (1939). «Строение белков». Природа . 143 (3625): 663–667. Бибкод : 1939Natur.143..663B . дои : 10.1038/143663a0 . S2CID 46327591 .
- ^ Виланд Т. и Бодански М., Мир пептидов , Springer Verlag, стр. 193–198. ISBN 0-387-52830-X
- ^ Хофманн А., Отт Х., Гриот Р., Стадлер П.А., Фрей А.Дж. (1963). «Синтез фон Эрготамина». Helvetica Chimica Acta . 46 : 2306–2336. дои : 10.1002/hlca.19630460650 .
- ^ Шемякин М.М., Антонов В.К., Шкроб А.М. (1963). «Активация амидной группы путем ацилирования». Пептиды, учеб. 6-я Европа. Пепт. Симп., Афины : 319–328.
- ^ Занотти Г, Пиннен Ф, Лусенте Г, Черрини С, Федели В, Мацца Ф (1984). «Пептидные тиациклолы. Синтез и структурные исследования». Журнал Химического общества, Perkin Transactions 1 : 1153–1157. дои : 10.1039/p19840001153 .
- ^ Гриот Р.Г., Фрей Эй.Дж. (1963). «Образование циклолов из N-гидроксиациллактамов». Тетраэдр . 19 (11): 1661–1673. дои : 10.1016/S0040-4020(01)99239-7 .
- ^ Люсенте Дж, Ромео А (1971). «Синтез циклолов из небольших пептидов посредством амид-амидной реакции». хим. Коммун . ? : 1605–1607. дои : 10.1039/c29710001605 .
Роте М., Шиндлер В., Пудилл Р., Костшева У., Тейсон Р., Стейнбергер Р. (1971). К проблеме синтеза циклолтрипептидов . Пептиды, учеб. 11-е место в Европе Пепт. Симп. (на немецком языке). Вена. стр. 388–399.
Роте М., Розер К.Л. (1988). Конформационная гибкость циклических трипептидов . 20-я Европа. Пепт. Симп. Тюбинген. п. 36. - ^ Виланд Т., Мор Х (1956). «Диациламиды как богатые энергией соединения. Диглицилимид». Анна Либиха. Хим. (на немецком языке). 599 : 222-232. дои : 10.1002/jlac.19565990306 .
Виланд Т., Урбах Х. (1958). «Другие диаминоацилимиды и их внутримолекулярные перегруппировки». Анна Либиха. Хим. (на немецком языке). 613 : 84–95. дои : 10.1002/jlac.19586130109 .
Бреннер М. (1958). Уолстенхолм Дж.Э., О'Коннор К.М. (ред.). «Аминоацильная вставка». Симпозиум Фонда Ciba по аминокислотам и пептидам с антиметаболической активностью . - ^ Ринч ДМ (1957). «Структура бацитрацина А». Природа . 179 (4558): 536–537. Бибкод : 1957Natur.179..536W . дои : 10.1038/179536a0 . S2CID 4154444 .
Ринч ДМ (1957). «Подход к синтезу полициклических пептидов». Природа . 180 (4584): 502–503. Бибкод : 1957Natur.180..502W . дои : 10.1038/180502b0 . S2CID 4289278 .
Ринч Д. (январь 1962 г.). «Некоторые вопросы молекулярной биологии и последние достижения органической химии малых пептидов». Природа . 193 (4812): 245–247. Бибкод : 1962Natur.193..245W . дои : 10.1038/193245a0 . ПМИД 14008494 . S2CID 4252124 .
Ринч ДМ (1963). «Последние достижения в химии циклола». Природа . 199 (4893): 564–566. Бибкод : 1963Natur.199..564W . дои : 10.1038/199564a0 . S2CID 4177095 .
Ринч Д. (май 1965 г.). «Современная картина химических аспектов строения полипептидных цепей и некоторые проблемы молекулярной биологии». Природа . 206 (983): 459–461. Бибкод : 1965Natur.206..459W . дои : 10.1038/206459a0 . ПМИД 5319104 . S2CID 4190760 . - ^ Ринч ДМ (1960). Химические аспекты структуры малых пептидов: Введение . Копенгаген: Мунксгаард.
- ^ Ринч ДМ (1965). Химические аспекты структуры полипептидных цепей и теория циклола . Нью-Йорк: Пленум Пресс.
Дальнейшее чтение
[ редактировать ]- «Белковые единицы, помещенные в графическую «клетку» » . Нью-Йорк Таймс . 19 апреля 1940 г. с. 14.
- «Теория белков вафельницы» . Нью-Йорк Таймс . 2 февраля 1947 г. с. Е9.
- Сенешаль М , изд. (28–30 сентября 1977 г.). Структуры материи и закономерности в науке: вдохновленные работой и жизнью Дороти Ринч, 1894–1976 гг . Материалы симпозиума, состоявшегося в Смит-колледже. Нортгемптон, Массачусетс: Издательская компания Schenkman.
Избранные статьи Дороти Ринч из собрания Софии Смит. Издательская компания Шенкман
- Сенешаль, Марджори (1980). «Избранные статьи Дороти Ринч из коллекции Софии Смит». Структуры материи и закономерности в науке . Издательство Шенкман. ISBN 978-0-87073-908-8 .
- Сенешаль М (2013). Я умер за красоту: Дороти Ринч и научная культура . Издательство Оксфордского университета. ISBN 978-0-19-973259-3 .