Биомолекулярная структура
Эта статья может сбивать с толку или быть непонятной читателям . ( февраль 2016 г. ) |
 |  |
Биомолекулярная структура — это сложная сложенная трехмерная форма, образованная молекулой , ДНК белка или РНК , и это важно для ее функции. Структуру этих молекул можно рассматривать на любом из нескольких масштабов: от уровня отдельных атомов до взаимоотношений между целыми белковыми субъединицами . Это полезное различие между шкалами часто выражается как разложение молекулярной структуры на четыре уровня: первичный, вторичный, третичный и четверичный. Каркас для этой многомасштабной организации молекулы возникает на вторичном уровне, где фундаментальными структурными элементами являются различные водородные связи молекулы . Это приводит к появлению нескольких узнаваемых доменов структуры белка и структуры нуклеиновой кислоты , включая такие особенности вторичной структуры, как альфа-спирали и бета-листы для белков, а также шпильки , выпуклости и внутренние петли для нуклеиновых кислот.Термины первичная , вторичная , третичная и четверичная структура были введены Кай Ульрик Линдерстрем-Ланг на своих медицинских лекциях в Стэнфордском университете в 1951 году .
Первичная структура
[ редактировать ]Первичная структура биополимера — это точная спецификация его атомного состава и химических связей, соединяющих эти атомы (включая стереохимию ). Для типичного неразветвленного, несшитого биополимера (такого как молекула типичного внутриклеточного белка , ДНК или РНК ) первичная структура эквивалентна указанию последовательности его мономерных субъединиц, таких как аминокислоты или нуклеотиды .
О первичной структуре белка сообщается, начиная с амино -N-конца до карбоксильного С-конца , тогда как первичная структура молекулы ДНК или РНК известна как последовательность нуклеиновой кислоты, сообщаемая от 5'-конца к 3'-концу .Последовательность нуклеиновой кислоты относится к точной последовательности нуклеотидов, составляющих всю молекулу. Часто первичная структура кодирует мотивы последовательности , имеющие функциональное значение. Некоторые примеры таких мотивов: C/D [1] и коробки H/ACA [2] мякРНК , , сайт связывания LSm обнаруженный в сплайсосомальных РНК, таких как U1 , U2 , U4 , U5 , U6 , U12 и U3 , последовательность Шайна-Дальгарно , [3] Козака консенсусная последовательность [4] и терминатор РНК-полимеразы III . [5]
Вторичная структура
[ редактировать ]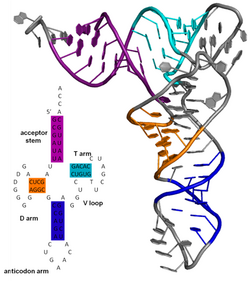
Вторичная структура белка представляет собой структуру водородных связей в биополимере. Они определяют общую трехмерную форму локальных сегментов биополимеров, но не описывают глобальную структуру конкретных атомных позиций в трехмерном пространстве, которые считаются третичной структурой . Вторичная структура формально определяется водородными связями биополимера, как это наблюдается в структуре с атомным разрешением. В белках вторичная структура определяется характером водородных связей между амином основной цепи и карбоксильными группами (водородные связи боковая цепь-основная цепь и боковая цепь-боковая цепь не имеют значения), где DSSP используется определение водородной связи .
Вторичная структура нуклеиновой кислоты определяется водородными связями между азотистыми основаниями.
Однако в белках водородная связь коррелирует с другими структурными особенностями, что привело к менее формальным определениям вторичной структуры. Например, спирали могут принимать основные двугранные углы в некоторых областях графика Рамачандрана ; таким образом, участок остатков с такими двугранными углами часто называют спиралью , независимо от того, имеет ли он правильные водородные связи. Было предложено множество других, менее формальных определений, часто с применением концепций дифференциальной геометрии кривых, таких как кривизна и кручение . Структурные биологи, решающие новую структуру с атомным разрешением, иногда на глаз определяют ее вторичную структуру и записывают свои назначения в соответствующий файл банка данных белков (PDB).
Вторичная структура молекулы нуклеиновой кислоты относится к взаимодействиям пар оснований внутри одной молекулы или набора взаимодействующих молекул. Вторичную структуру биологических РНК часто можно однозначно разложить на стебли и петли. Часто эти элементы или их комбинации можно дополнительно классифицировать, например, тетрапетли , псевдоузлы и стеблевые петли . Существует множество элементов вторичной структуры, имеющих функциональное значение для биологической РНК. Известные примеры включают Rho-независимые петли терминатора и клеверный лист транспортной РНК (тРНК). Существует небольшая отрасль исследователей, пытающихся определить вторичную структуру молекул РНК. Подходы включают как экспериментальные , так и вычислительные методы (см. также Список программного обеспечения для прогнозирования структуры РНК ).
Третичная структура
[ редактировать ]Третичная структура белка или любой другой макромолекулы — это ее трехмерная структура, определяемая координатами атомов. [6] Белки и нуклеиновые кислоты складываются в сложные трехмерные структуры, которые определяют функции молекул. Хотя такие структуры разнообразны и сложны, они часто состоят из повторяющихся, узнаваемых мотивов третичной структуры и доменов, которые служат молекулярными строительными блоками. биомолекулы Считается, что третичная структура во многом определяется первичной структурой (ее последовательностью аминокислот или нуклеотидов ).
Четвертичная структура
[ редактировать ]белка Четвертичная структура [а] относится к количеству и расположению нескольких белковых молекул в многосубъединичном комплексе.
Для нуклеиновых кислот этот термин менее распространен, но может относиться к организации ДНК более высокого уровня в хроматине . [7] включая его взаимодействие с гистонами или взаимодействия между отдельными единицами РНК в рибосоме. [8] [9] или сплайсосома .
Определение структуры
[ редактировать ]Зондирование структуры — это процесс, с помощью которого биохимические методы используются для определения биомолекулярной структуры. [10] Этот анализ можно использовать для определения закономерностей, которые можно использовать для вывода о молекулярной структуре, экспериментального анализа молекулярной структуры и функций, а также для дальнейшего понимания разработки более мелких молекул для дальнейших биологических исследований. [11] Анализ структурного зондирования можно проводить с помощью множества различных методов, включая химическое зондирование, зондирование гидроксильных радикалов, интерференционное картирование аналогов нуклеотидов (NAIM) и поточное зондирование. [10]
Структуры белков и нуклеиновых кислот можно определить с помощью спектроскопии ядерного магнитного резонанса ( ЯМР ), рентгеновской кристаллографии или одночастичной криоэлектронной микроскопии ( криоЭМ ). В первых опубликованных отчетах о ДНК ( Розалиндой Франклин и Рэймондом Гослингом А-ДНК в 1953 году) о рентгенограммах , а также B-ДНК, использовался анализ, основанный на преобразованиях функций Паттерсона , которые предоставили лишь ограниченное количество структурной информации для ориентированных волокна ДНК, выделенные из тимуса теленка . [12] [13] Альтернативный анализ был затем предложен Wilkins et al. в 1953 году для рентгеновской дифракции B-ДНК и картин рассеяния гидратированных, бактериально-ориентированных волокон ДНК и головок сперматозоидов форели в терминах квадратов функций Бесселя . [14] Хотя форма B-ДНК наиболее распространена в условиях, встречающихся в клетках, [15] это не четко определенная конформация, а семейство или нечеткий набор конформаций ДНК, которые возникают при высоких уровнях гидратации, присутствующих в самых разных живых клетках. [16] Соответствующие им рентгенограммы и картины рассеяния характерны для молекулярных паракристаллов со значительной степенью беспорядка (более 20%), [17] [18] и структуру невозможно определить, используя только стандартный анализ.
Напротив, стандартный анализ, включающий только преобразования Фурье Бесселя функций [19] ДНК и молекулярные модели до сих пор обычно используются для анализа рентгенограмм A-ДНК и Z-ДНК. [20]
Прогнозирование структуры
[ редактировать ]
Предсказание биомолекулярной структуры — это предсказание трехмерной структуры белка по его аминокислотной последовательности или нуклеиновой кислоты по его последовательности нуклеиновых оснований. Другими словами, это предсказание вторичной и третичной структуры на основе ее первичной структуры. Предсказание структуры является обратным по отношению к биомолекулярному дизайну, как в рациональном дизайне , дизайне белка , дизайне нуклеиновых кислот и биомолекулярной инженерии .
Предсказание структуры белка — одна из важнейших задач биоинформатики и теоретической химии . Прогнозирование структуры белка имеет большое значение в медицине (например, при разработке лекарств ) и биотехнологии (например, при разработке новых ферментов ). Каждые два года эффективность текущих методов оценивается в эксперименте «Критическая оценка прогнозирования структуры белка » ( CASP ).
Также было проведено значительное количество биоинформатических исследований, направленных на проблему предсказания структуры РНК. Общей проблемой исследователей, работающих с РНК, является определение трехмерной структуры молекулы, зная только последовательность нуклеиновой кислоты. Однако в случае РНК большая часть окончательной структуры определяется вторичной структурой или внутримолекулярными взаимодействиями спаривания оснований молекулы. Об этом свидетельствует высокая консервативность пар оснований у разных видов.
Вторичная структура небольших молекул нуклеиновой кислоты определяется в основном сильными локальными взаимодействиями, такими как водородные связи и укладка оснований . Суммирование свободной энергии таких взаимодействий, обычно с использованием метода ближайших соседей , дает аппроксимацию стабильности данной структуры. [21] Самый простой способ найти структуру с наименьшей свободной энергией — это сгенерировать все возможные структуры и вычислить для них свободную энергию, но количество возможных структур для последовательности увеличивается экспоненциально с длиной молекулы. [22] Для более длинных молекул количество возможных вторичных структур огромно. [21]
Методы ковариации последовательностей основаны на существовании набора данных, состоящего из множества гомологичных последовательностей РНК со родственными, но несходными последовательностями. Эти методы анализируют ковариацию отдельных базовых сайтов в эволюции ; сохранение в двух широко разделенных сайтах пары нуклеотидов , спаривающих основания , указывает на наличие структурно необходимой водородной связи между этими положениями. Было показано, что общая проблема прогнозирования псевдоузлов является NP-полной . [23]
Дизайн
[ редактировать ]Биомолекулярный дизайн можно считать обратным предсказанию структуры. При предсказании структуры структура определяется на основе известной последовательности, тогда как при проектировании белка или нуклеиновой кислоты генерируется последовательность, которая будет формировать желаемую структуру.
Другие биомолекулы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( апрель 2010 г. ) |
Другие биомолекулы, такие как полисахариды , полифенолы и липиды , также могут иметь структуру более высокого порядка биологического значения.
См. также
[ редактировать ]- Биомолекулярный
- Сравнение программного обеспечения для моделирования нуклеиновых кислот
- Генная структура
- Список программного обеспечения для прогнозирования структуры РНК
- Некодирующая РНК
Примечания
[ редактировать ]- ^ Здесь четвертичность означает « четвертого уровня структуру », а не « четырехстороннее взаимодействие». Этимологически четвертичная система правильна: четвертичная система происходит от латинских распределительных чисел и следует за двоичной и троичной системой ; в то время как четвертичный происходит от латинских порядковых чисел и следует за вторичным и третичным . Однако четвертичность является стандартной в биологии.
Ссылки
[ редактировать ]- ^ Самарский Д.А., Фурнье М.Ж., Сингер Р.Х., Бертран Э. (июль 1998 г.). «Мотив C/D-бокса мякРНК управляет ядрышковым нацеливанием, а также связывает синтез и локализацию мякРНК» . Журнал ЭМБО . 17 (13): 3747–57. дои : 10.1093/emboj/17.13.3747 . ПМК 1170710 . ПМИД 9649444 .
- ^ Гано П., Кайзерг-Феррер М., Кисс Т. (апрель 1997 г.). «Семейство малых ядрышковых РНК бокса ACA определяется эволюционно консервативной вторичной структурой и повсеместными элементами последовательности, необходимыми для накопления РНК» . Гены и развитие . 11 (7): 941–56. дои : 10.1101/gad.11.7.941 . ПМИД 9106664 .
- ^ Шайн Дж. , Далгарно Л. (март 1975 г.). «Определитель цистронной специфичности бактериальных рибосом». Природа . 254 (5495): 34–38. Бибкод : 1975Natur.254...34S . дои : 10.1038/254034a0 . ПМИД 803646 . S2CID 4162567 .
- ^ Козак М. (октябрь 1987 г.). «Анализ 5'-некодирующих последовательностей 699 информационных РНК позвоночных» . Исследования нуклеиновых кислот . 15 (20): 8125–48. дои : 10.1093/нар/15.20.8125 . ПМК 306349 . ПМИД 3313277 .
- ^ Богенхаген Д.Ф., Браун Д.Д. (апрель 1981 г.). «Нуклеотидные последовательности в ДНК Xenopus 5S, необходимые для терминации транскрипции». Клетка . 24 (1): 261–70. дои : 10.1016/0092-8674(81)90522-5 . ПМИД 6263489 . S2CID 9982829 .
- ^ ИЮПАК , Сборник химической терминологии , 2-е изд. («Золотая книга») (1997). Исправленная онлайн-версия: (2006–) « Третичная структура ». два : 10.1351/goldbook.T06282
- ^ Сипски М.Л., Вагнер Т.Э. (март 1977 г.). «Изучение четвертичной упорядоченности ДНК с помощью спектроскопии кругового дихроизма: исследование хромосомных волокон спермы лошадей». Биополимеры . 16 (3): 573–82. дои : 10.1002/bip.1977.360160308 . ПМИД 843604 . S2CID 35930758 .
- ^ Ноллер Х.Ф. (1984). «Структура рибосомальной РНК». Ежегодный обзор биохимии . 53 : 119–62. дои : 10.1146/annurev.bi.53.070184.001003 . ПМИД 6206780 .
- ^ Ниссен П., Ипполито Дж.А., Бан Н., Мур П.Б., Стейц Т.А. (апрель 2001 г.). «Третичные взаимодействия РНК в большой субъединице рибосомы: А-минорный мотив» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (9): 4899–903. Бибкод : 2001PNAS...98.4899N . дои : 10.1073/pnas.081082398 . ПМК 33135 . ПМИД 11296253 .
- ^ Jump up to: а б Теуниссен, AWM (1979). Исследование структуры РНК: анализ биохимической структуры молекул РНК, связанных с аутоиммунными заболеваниями . стр. 1–27. ISBN 978-90-901323-4-1 .
- ^ Пейс Н.Р., Томас Б.К., Вёзе Ч.Р. (1999). Исследование структуры, функции и истории РНК путем сравнительного анализа . Лабораторный пресс Колд-Спринг-Харбор. стр. 113–17. ISBN 978-0-87969-589-7 .
- ^ Франклин Р.Э. , Гослинг Р.Г. (6 марта 1953 г.). «Структура волокон тимонуклеата натрия (I. Влияние содержания воды и II. Цилиндрически симметричная функция Паттерсона)» (PDF) . Акта Кристаллогр . 6 (8): 673–78. дои : 10.1107/s0365110x53001939 .
- ^ Франклин Р.Э., Гослинг Р.Г. (апрель 1953 г.). «Молекулярная конфигурация тимонуклеата натрия». Природа . 171 (4356): 740–41. Бибкод : 1953Natur.171..740F . дои : 10.1038/171740a0 . ПМИД 13054694 . S2CID 4268222 .
- ^ Уилкинс М.Х., Стоукс А.Р., Уилсон Х.Р. (апрель 1953 г.). «Молекулярная структура дезоксипентозных нуклеиновых кислот». Природа . 171 (4356): 738–40. Бибкод : 1953Natur.171..738W . дои : 10.1038/171738a0 . ПМИД 13054693 . S2CID 4280080 .
- ^ Лесли А.Г., Арнотт С., Чандрасекаран Р., Рэтлифф Р.Л. (октябрь 1980 г.). «Полиморфизм двойных спиралей ДНК». Журнал молекулярной биологии . 143 (1): 49–72. дои : 10.1016/0022-2836(80)90124-2 . ПМИД 7441761 .
- ^ Баяну, IC (1980). «Структурный порядок и частичный беспорядок в биологических системах». Бык. Математика. Биол . 42 (1): 137–41. дои : 10.1007/BF02462372 . S2CID 189888972 .
- ^ Хоземанн Р., Багчи Р.Н. (1962). Прямой анализ дифракции на веществе . Амстердам/Нью-Йорк: Северная Голландия.
- ^ Баяну И.С. (1978). «Рассеяние рентгеновских лучей частично неупорядоченными мембранными системами». Акта Кристаллогр. А. 34 (5): 751–53. Бибкод : 1978AcCrA..34..751B . дои : 10.1107/s0567739478001540 .
- ^ «Функции Бесселя и дифракция на спиральных структурах» . PlanetPhysics.org . [ постоянная мертвая ссылка ]
- ^ «Рентгенограммы кристаллов двуспиральной дезоксирибонуклеиновой кислоты (ДНК)» . PlanetPhysics.org . Архивировано из оригинала 24 июля 2009 года.
- ^ Jump up to: а б Мэтьюз Д.Х. (июнь 2006 г.). «Революции в предсказании вторичной структуры РНК». Журнал молекулярной биологии . 359 (3): 526–32. дои : 10.1016/j.jmb.2006.01.067 . ПМИД 16500677 .
- ^ Цукер М., Санкофф Д. (1984). «Вторичные структуры РНК и их предсказание». Бык. Математика. Биол . 46 (4): 591–621. дои : 10.1007/BF02459506 . S2CID 189885784 .
- ^ Люнгсё Р.Б., Педерсен К.Н. (2000). «Предсказание псевдоузла РНК в энергетических моделях». Журнал вычислительной биологии . 7 (3–4): 409–27. CiteSeerX 10.1.1.34.4044 . дои : 10.1089/106652700750050862 . ПМИД 11108471 .