ВНК1
| ВНК1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | WNK1 , HSAN2, HSN2, KDP, PPP1R167, PRKPSK, p65, протеинкиназа 1 с дефицитом лизина WNK | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 605232 ; МГИ : 2442092 ; Гомологен : 14253 ; Генные карты : WNK1 ; ОМА : WNK1 — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||





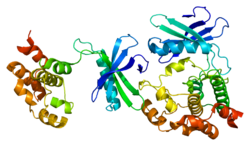
WNK (протеинкиназа 1 с дефицитом лизина) , также известная как WNK1 , представляет собой фермент, кодируемый WNK1 геном . [ 5 ] [ 6 ] [ 7 ] [ 8 ] [ 9 ] WNK1 представляет собой серин-треониновую протеинкиназу и входит в семейство киназ WNK «без лизина/K». [ 5 ] [ 6 ] [ 7 ] [ 9 ] Преобладающая роль WNK1 заключается в регуляции катиона Cl. − котранспортеры (CCC), такие как котранспортер хлорида натрия ( NCC ), базолатеральный симпортер Na-K-Cl ( NKCC1 ) и котранспортер хлорида калия (KCC1), расположенные в почках. [ 5 ] [ 6 ] [ 9 ] CCC опосредуют ионный гомеостаз и модулируют кровяное давление путем транспортировки ионов в клетку и из клетки . [ 5 ] В результате мутации WNK1 были вовлечены в нарушения/заболевания артериального давления; Ярким примером является семейная гиперкалиемическая гипертензия (СГГТ). [ 5 ] [ 6 ] [ 7 ] [ 8 ] [ 9 ]
Структура
[ редактировать ]WNK1 Белок состоит из 2382 аминокислот (молекулярная масса 230 кДа). [ 8 ] Белок N содержит киназный домен, расположенный внутри его короткого -концевого домена , и длинный С-концевой хвост. [ 8 ] Киназный семейством домен имеет некоторое сходство с протеинкиназ MEKK . [ 8 ] Как член семейства WNK, каталитический остаток лизина киназы уникально расположен в бета-цепи 2 глициновой петли . [ 8 ] Чтобы обладать киназной активностью, WNK1 должен аутофосфорилировать остаток серина 382, обнаруженный в его петле активации. [ 8 ] [ 5 ] Кроме того, фосфорилирование в другом сайте (Ser378) увеличивает активность WNK1. [ 5 ] Аутоингибирующий домен расположен внутри C-концевого домена вместе с доменом HQ, который необходим для взаимодействия WNK1 с другими WNK. [ 5 ] [ 6 ] [ 7 ] [ 8 ] Взаимодействия между WNK играют важную роль в функционировании; Мутанты WNK1 , у которых отсутствует домен HQ, также лишены киназной активности.
Функция
[ редактировать ]Ген WNK1 кодирует цитоплазматическую серин-треониновую киназу, экспрессируемую в дистальных отделах нефрона . [ 5 ] [ 6 ] [ 8 ] Исследования показали, что WNK1 может активировать несколько CCC. [ 5 ] [ 6 ] WNK1, однако, не фосфорилирует напрямую сами CCC, а фосфорилирует другие серин-треониновые киназы : родственную Sterile20 пролин-аланин-богатую киназу (SPAK) и киназу 1 реакции окислительного стресса ( OXSR1 ). [ 6 ] [ 5 ] [ 7 ] Фосфорилирование Т-петли SPAK, расположенной в его каталитическом домене, активирует SPAK, что приведет к фосфорилированию N-концевого домена CCC. [ 5 ] [ 6 ] Следовательно, WNK1 косвенно активирует CCC как вышестоящий регулятор SPAK/OSR1. [ 5 ] [ 6 ] [ 7 ]
Реабсорбция натрия
[ редактировать ]

В дистальных извитых канальцах (DCT) WNK1 является мощным активатором NCC, что приводит к увеличению реабсорбции натрия , что приводит к повышению артериального давления. [ 5 ] [ 6 ] [ 7 ] Мутант WNK1, обнаруженный в FHHt, содержит большую делецию в интроне 1, которая вызывает увеличение экспрессии полноразмерного WNK1. [ 5 ] [ 6 ] [ 7 ] [ 8 ] Повышение уровня WNK1 приводит к увеличению активации NCC, что способствует повышению артериального давления/ гипертензии, связанной с FHHt. [ 5 ] [ 6 ] [ 7 ] [ 8 ] WNK1 активирует индуцируемую сывороткой и глюкокортикоидами протеинкиназу SGK1 , что приводит к усилению экспрессии эпителиального натриевого канала (ENaC), что также способствует реабсорбции натрия. [ 6 ]
Секреция калия
[ редактировать ]WNK1 регулирует калиевые каналы, обнаруженные в кортикальных собирательных трубочках (CCD) и соединительных канальцах (CNT). [ 6 ] Почечный наружный мозговой калий 1 ( ROMK1 ) и кальций-активируемый калиевый канал большой проводимости l (BKCa) являются двумя основными каналами секреции калия. [ 6 ] WNK1 косвенно стимулирует клатрин-зависимый эндоцитоз ROMK1 за счет потенциального взаимодействия с интерсектином (ITSN1); таким образом, киназная активность не требуется. [ 6 ] Другим возможным механизмом регуляции ROMK1 является аутосомно-рецессивная гиперхолестеринемия (ACH), которая представляет собой адапторную молекулу клатрина. [ 6 ] Фосфорилирование ACH с помощью WNK1 способствует транслокации ROMK1 в покрытые клатрином ямки, запуская эндоцитоз . [ 6 ] WNK1 может косвенно активировать BKCa путем ингибирования действия киназ, регулируемых внеклеточными сигналами (ERK1 и ERK2), которые приводят к лисомальной деградации. [ 6 ]
Регулирование объема клеток
[ редактировать ]Котранспортеры NKCC1/2 регулируются внутриклеточным Cl. − концентрация. [ 9 ] Исследования указывают на WNK1 как на ключевой эффектор, связывающий Cl − концентрация до функции NKCC1/2. [ 5 ] [ 9 ] При гипертоническом (высокий внеклеточный Cl − ) условия, которые вызывают сокращение клеток, неизвестный механизм усиливает экспрессию WNK1, чтобы противодействовать потере объема. [ 5 ] Увеличение WNK1 приводит к активации SPAK/OSR1, которые активируют NKCC1/2 посредством последующего фосфорилирования. [ 5 ] [ 9 ] NKCC1/2 будет способствовать притоку Na + , К + и Cl − ионы проникают в клетку, тем самым вызывая приток воды в клетку. [ 5 ] В обратных обстоятельствах, когда гипотонический (низкий внеклеточный Cl − ) условия вызывают набухание клеток, WNK1 ингибируется. [ 5 ] Другой котранспортер, KCC, неактивен при фосфорилировании; без активированного WNK1 KCC не подвергается фосфорилированию и может активироваться. [ 5 ] Котранспортер будет способствовать оттоку K + и Cl − ионы и вызывают отток воды из клетки для борьбы с отеком. [ 5 ]
WNK1 в мозге
[ редактировать ]В зрелом мозге нейротрансмиттер ГАМК представляет собой основной тормозной сигнал, используемый в передаче сигналов нейронов. [ 5 ] ГАМК активирует ГАМК А рецептор , который представляет собой Cl. − ионный канал. [ 5 ] кл − ионы проникнут в нейрон, вызывая гиперполяризацию и ингибирование передачи сигналов. [ 5 ] Однако во время развития мозга активация ГАМК А позволит Cl − ионы покидают нейрон, вызывая деполяризацию нейрона. [ 5 ] Таким образом, ГАМК является возбуждающим нейромедиатором во время развития. [ 5 ] WNK1 участвует в переключении в процессе развития с возбуждающей на ингибирующую передачу сигналов ГАМК посредством взаимодействия с NKCC1 и KCC. [ 5 ] WNK1 фосфорилирует SPAK/OSR1, который затем фосфорилирует KCC2, ингибируя поток Cl. − ионы выходят из клетки в процессе развития. [ 5 ]

Регуляция WNK1
[ редактировать ]Концентрации Cl − ионы и К + ион играет важную роль в регуляции активности WNK1. [ 5 ] [ 9 ] В ДКП концентрация K в плазме + Считается, что ион влияет на концентрацию Cl − ионы внутри нефрона. [ 5 ] [ 9 ] Высокий плазменный K + Снижение концентрации регулирует активность WNK1 и предотвращает Cl − ион попадает в нефрон через NCC. [ 5 ] [ 9 ] Обратное происходит, когда плазма K + концентрация низкая; повышенная активность WNK1 повышает активность NCC, способствуя реабсорбции Cl − ионы. [ 5 ] [ 9 ] Когда имеется избыток Cl − ионов внутри нефрона , активность WNK1 ингибируется связыванием Cl − ион к каталитическому домену WNK1. [ 5 ] [ 9 ]
Более того, WNK1 и WNK4 могут взаимодействовать с образованием гетеродимеров, которые ингибируют функцию WNK1. [ 7 ] [ 6 ] Высвобождение WNK4 из гетеродимера позволяет мономеру WNK1 связываться с другим мономером WNK1, способствуя активации. [ 6 ] [ 7 ] Функция WNK1 также может быть ингибирована, если WNK1 деградирует. За убиквитинирование WNK1 ответственны два фермента: кельч-подобный 3 (KLHL3) и кулин-3 (CUL3). [ 7 ] [ 6 ] [ 10 ] KLHL3 служит адаптерным белком, который способствует взаимодействию между WNK1 и Cullin3, который находится в комплексе, содержащем убиквитинлигазу E3, которая присоединяет молекулы убиквитина к WNK1. [ 7 ] Убиквитинированный WNK1 впоследствии подвергается протеасомной деградации. [ 7 ] [ 6 ] [ 10 ]
Клиническое значение
[ редактировать ]WNK1 имеет мутации, связанные с синдромом гиперкалиемии-гипертензии Гордона ( псевдогипоальдостеронизм типа II, характеризующийся гипертонией, также называемой семейной гиперкалиемической гипертензией (FHHt)). [ 5 ] [ 7 ] [ 8 ] и врожденная сенсорная нейропатия ( HSAN характеризующаяся потерей восприятия боли типа II , , прикосновения и тепла из-за потери периферических сенсорных нервов ). [ 5 ] [ 11 ]
Сравнительная геномика
[ редактировать ]Ген принадлежит к группе из четырех родственных протеинкиназ (WNK1, WNK2 , WNK3 , WNK4 ). [ 5 ] [ 7 ] [ 8 ]
Гомологи этого белка были обнаружены у Arabidopsis thaliana , C. elegans , Chlamydomonas reinhardtii и Vitis vinifera , а также у позвоночных, включая Danio rerio и Taeniopygia Guttata . [ 7 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000060237 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: Ensembl, выпуск 89: ENSMUSG00000045962 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль Шекараби М., Чжан Дж., Ханна А.Р., Эллисон Д.Х., Дельпир Э., Кале К.Т. (февраль 2017 г.). «Передача сигналов WNK-киназы в ионном гомеостазе и заболеваниях человека» . Клеточный метаболизм . 25 (2): 285–299. дои : 10.1016/j.cmet.2017.01.007 . hdl : 10871/33390 . ПМИД 28178566 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х Хачуэль Дж., Эллисон Д.Х., Гамба Дж. (2016). «Регуляция почечного транспорта электролитов киназами WNK и SPAK-OSR1». Ежегодный обзор физиологии . 78 : 367–89. doi : 10.1146/annurev-physical-021115-105431 . ПМИД 26863326 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Базуа-Валенти С., Гамба Г. (май 2015 г.). «Возврат к регуляции котранспортера NaCl с помощью киназ без лизина» . Американский журнал физиологии. Клеточная физиология . 308 (10): C779-91. doi : 10.1152/ajpcell.00065.2015 . ПМЦ 4436992 . ПМИД 25788573 .
- ^ Jump up to: а б с д и ж г час я дж к л м Сюй Б.Э., Ли Б.Х., Мин X, Ленертц Л., Хейзе С.Дж., Стиппек С., Голдсмит Э.Дж., Кобб М.Х. (январь 2005 г.). «WNK1: анализ структуры протеинкиназы, последующих мишеней и потенциальной роли в гипертонии» . Клеточные исследования . 15 (1): 6–10. дои : 10.1038/sj.cr.7290256 . ПМИД 15686619 . S2CID 22087578 .
- ^ Jump up to: а б с д и ж г час я дж к л Хуан CL, Ченг CJ (ноябрь 2015 г.). «Объединяющий механизм регуляции киназы WNK котранспортера хлорида натрия» . Архив Пфлюгерса . 467 (11): 2235–41. дои : 10.1007/s00424-015-1708-2 . ПМК 4601926 . ПМИД 25904388 .
- ^ Jump up to: а б Алесси Д.Р., Чжан Дж., Ханна А., Хохдёрфер Т., Шан Ю., Кале К.Т. (июль 2014 г.). «Путь WNK-SPAK/OSR1: главный регулятор катион-хлорид-котранспортеров». Научная сигнализация . 7 (334): Лe3. дои : 10.1126/scisignal.2005365 . hdl : 10871/33417 . ПМИД 25028718 . S2CID 206672635 .
- ^ Тан Б.Л. (июль 2016 г.). «(WNK) в момент смерти: безлизиновые (Wnk) киназы при невропатиях и выживании нейронов». Бюллетень исследований мозга . 125 : 92–8. дои : 10.1016/j.brainresbull.2016.04.017 . ПМИД 27131446 . S2CID 3938880 .