Точечная мутация


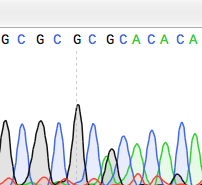
Точечная мутация — это генетическая мутация , при которой одно нуклеотидное основание изменяется, вставляется или удаляется из последовательности ДНК или РНК генома организма. [ 1 ] Точечные мутации оказывают разнообразные эффекты на последующие белковые продукты, причем последствия умеренно предсказуемы в зависимости от особенностей мутации. Эти последствия могут варьироваться от отсутствия эффекта (например, синонимические мутации ) до вредных эффектов (например, мутации сдвига рамки считывания ) в отношении продукции, состава и функции белка.
Причины
[ редактировать ]Точечные мутации обычно происходят во время репликации ДНК . Репликация ДНК происходит, когда одна двухцепочечная молекула ДНК создает две одиночные цепи ДНК, каждая из которых является матрицей для создания комплементарной цепи. Одна точечная мутация может изменить всю последовательность ДНК. Замена одного пурина или пиримидина может привести к изменению аминокислоты, которую нуклеотиды кодируют .
Точечные мутации могут возникать в результате спонтанных мутаций , возникающих во время репликации ДНК . Скорость мутации может быть увеличена мутагенами . Мутагены могут быть физическими, например, излучением УФ-лучей , рентгеновских лучей или сильной жары, или химическими (молекулы, которые перемещают пары оснований неправильно или нарушают спиральную форму ДНК). Мутагены, связанные с раком, часто изучаются, чтобы узнать о раке и его профилактике.
Существует несколько способов возникновения точковых мутаций. Во-первых, ультрафиолетовый (УФ) свет и высокочастотный свет обладают ионизирующей способностью, что, в свою очередь, может влиять на ДНК. Реактивные молекулы кислорода со свободными радикалами, которые являются побочным продуктом клеточного метаболизма, также могут быть очень вредными для ДНК. Эти реагенты могут приводить как к одноцепочечным, так и к двухцепочечным разрывам ДНК. В-третьих, связи в ДНК со временем разрушаются, что создает еще одну проблему для поддержания целостности ДНК на высоком уровне. Также могут возникнуть ошибки репликации, которые приводят к мутациям замены, вставки или делеции.
Категоризация
[ редактировать ]Категоризация перехода/трансверсии
[ редактировать ]
В 1959 году Эрнст Фриз ввел термины «переходы» или «трансверсии» для классификации различных типов точковых мутаций. [ 2 ] [ 3 ] Переходы – это замена пуринового основания другим пуриновым или замена пиримидина другим пиримидином. Трансверсии – это замена пурина на пиримидин или наоборот. Существует систематическая разница в частоте мутаций переходов (Альфа) и трансверсий (Бета). Переходные мутации встречаются примерно в десять раз чаще, чем трансверсии.
Функциональная категоризация
[ редактировать ]К бессмысленным мутациям относятся стоп-прирост и старт-проигрыш. Стоп-гейн — это мутация, которая приводит к преждевременному образованию кодона терминации ( был получен стоп ), что сигнализирует об окончании трансляции. Это прерывание приводит к аномальному укорочению белка. Количество потерянных аминокислот влияет на функциональность белка и на то, будет ли он вообще функционировать. [ 4 ] Стоп-лосс — это мутация исходного кодона терминации ( стоп-кодон потерян ), приводящая к аномальному удлинению карбоксильного конца белка. Start-gain создает стартовый кодон AUG выше исходного стартового сайта. Если новый AUG находится рядом с исходным стартовым сайтом, внутри рамки обрабатываемого транскрипта и ниже сайта связывания рибосом, его можно использовать для инициации трансляции. Вероятным эффектом является добавление дополнительных аминокислот к аминоконцу исходного белка. Мутации со сдвигом рамки также возможны при мутациях «старт-прирост», но обычно не влияют на трансляцию исходного белка. Старт-потеря — это точечная мутация стартового кодона AUG транскрипта, приводящая к снижению или устранению продукции белка.
Миссенс-мутации кодируют другую аминокислоту. Миссенс-мутация изменяет кодон, в результате чего создается другой белок, что является несинонимичным изменением. [ 4 ] Консервативные мутации приводят к замене аминокислот. Однако свойства аминокислоты остаются прежними (например, гидрофобность, гидрофильность и т. д.). Иногда изменение одной аминокислоты в белке не несет вреда для организма в целом. Большинство белков могут противостоять одной или двум точечным мутациям, прежде чем их функция изменится. Неконсервативные мутации приводят к изменению аминокислоты, свойства которой отличаются от свойств дикого типа . Белок может потерять свою функцию, что может привести к заболеванию организма. Например, серповидноклеточная анемия вызывается единственной точечной мутацией (миссенс-мутация) в бета- гемоглобина гене GAG , которая превращает кодон в GUG, который кодирует аминокислоту валин, а не глутаминовую кислоту . Белок также может проявлять «усиление функции» или активироваться, как в случае с мутацией, превращающей валин в глутаминовую кислоту в гене BRAF ; это приводит к активации белка RAF, который вызывает неограниченную пролиферативную передачу сигналов в раковых клетках. [ 5 ] Оба эти примера являются примерами неконсервативной (миссенс) мутации.
Тихие мутации кодируют одну и ту же аминокислоту (« синонимная замена »). Тихая мутация не влияет на функционирование белка . Один нуклеотид может измениться, но новый кодон определяет ту же аминокислоту, в результате чего белок не мутирует. Этот тип изменений называется синонимичным изменением, поскольку старый и новый кодоны кодируют одну и ту же аминокислоту. Это возможно, поскольку 64 кодона определяют только 20 аминокислот. Однако разные кодоны могут приводить к разным уровням экспрессии белков. [ 4 ]
Вставки и делеции одной пары оснований
[ редактировать ]Иногда термин «точечная мутация» используется для описания вставок или делеций одной пары оснований (что оказывает более неблагоприятное воздействие на синтезируемый белок, поскольку нуклеотиды по-прежнему считываются в триплетах, но в разных рамках: мутация, называемая сдвигом рамки считывания) . мутация ). [ 4 ]
Общие последствия
[ редактировать ]Точечные мутации, возникающие в некодирующих последовательностях, чаще всего не имеют последствий, хотя бывают и исключения. Если мутированная пара оснований находится в промоторной последовательности гена, то экспрессия гена может измениться. Кроме того, если мутация происходит в месте сплайсинга интрона , это может помешать правильному сплайсингу транскрибируемой пре-мРНК .
Изменив всего одну аминокислоту, можно изменить весь пептид , тем самым изменив весь белок. Новый белок называется вариантом белка . Если исходный белок участвует в клеточном воспроизводстве, то эта единственная точечная мутация может изменить весь процесс клеточного воспроизводства в этом организме.
Точечные зародышевые мутации могут приводить как к полезным, так и к вредным признакам или заболеваниям. Это приводит к адаптации, основанной на среде, в которой обитает организм. Выгодная мутация может создать преимущество для этого организма и привести к передаче признака из поколения в поколение, улучшая и принося пользу всей популяции. Научная теория эволюции во многом зависит от точечных мутаций в клетках . Теория объясняет разнообразие и историю живых организмов на Земле. Что касается точечных мутаций, в нем говорится, что полезные мутации позволяют организму процветать и размножаться, тем самым передавая положительно затронутые мутировавшие гены следующему поколению. С другой стороны, вредные мутации приводят к гибели организма или снижению его шансов на размножение в результате явления, известного как естественный отбор .
Существуют различные краткосрочные и долгосрочные последствия, которые могут возникнуть в результате мутаций. Меньшие из них будут означать остановку клеточного цикла во многих точках. Это означает, что кодон, кодирующий аминокислоту глицин, может быть заменен на стоп-кодон, в результате чего белки, которые должны были быть произведены, деформируются и не могут выполнять свои намеченные задачи. Поскольку мутации могут повлиять на ДНК и, следовательно, на хроматин , они могут препятствовать возникновению митоза из-за отсутствия полной хромосомы. Проблемы также могут возникнуть во время процессов транскрипции и репликации ДНК. Все это препятствует размножению клетки и, таким образом, приводит к ее гибели. Долгосрочные последствия могут заключаться в необратимом изменении хромосомы, которое может привести к мутации. Эти мутации могут быть как полезными, так и вредными. Рак является примером того, насколько они могут быть вредными. [ 6 ]
Другие эффекты точечных мутаций или однонуклеотидного полиморфизма в ДНК зависят от местоположения мутации внутри гена. Например, если мутация происходит в области гена, отвечающего за кодирование, аминокислотная последовательность кодируемого белка может быть изменена, вызывая изменение функции, локализации белка, стабильности белка или белкового комплекса. Было предложено множество методов для прогнозирования влияния миссенс-мутаций на белки. Алгоритмы машинного обучения обучают свои модели, чтобы отличать известные заболевания, связанные с нейтральными мутациями, тогда как другие методы не обучают свои модели явно, но почти все методы используют эволюционную консервацию, предполагая, что изменения в консервативных позициях имеют тенденцию быть более вредными. Хотя большинство методов обеспечивают бинарную классификацию эффектов мутаций на повреждающие и доброкачественные, необходим новый уровень аннотации, чтобы предложить объяснение того, почему и как эти мутации повреждают белки. [ 7 ]
Более того, если мутация происходит в области гена, где транскрипционный аппарат связывается с белком, мутация может повлиять на связывание факторов транскрипции, поскольку короткие нуклеотидные последовательности, распознаваемые факторами транскрипции, будут изменены. Мутации в этой области могут влиять на эффективность транскрипции генов, что, в свою очередь, может изменять уровни мРНК и, следовательно, уровни белка в целом.
Точечные мутации могут оказывать различное влияние на поведение и воспроизводство белка в зависимости от того, где происходит мутация в аминокислотной последовательности белка. Если мутация происходит в области гена, отвечающего за кодирование белка, аминокислота может быть изменена. Это небольшое изменение в последовательности аминокислот может вызвать изменение функции, активацию белка, то есть то, как он связывается с данным ферментом, где белок будет расположен внутри клетки, или количество свободной энергии, запасенной внутри белка. .
Если мутация происходит в области гена, где транскрипционный аппарат связывается с белком, мутация может повлиять на способ связывания транскрипционных факторов с белком. Механизмы транскрипции связываются с белком посредством узнавания коротких нуклеотидных последовательностей. Мутация в этой области может изменить эти последовательности и, таким образом, изменить способ связывания факторов транскрипции с белком. Мутации в этой области могут влиять на эффективность транскрипции генов, которая контролирует как уровни мРНК, так и общий уровень белка. [ 8 ]
Специфические заболевания, вызванные точковыми мутациями
[ редактировать ]Рак
[ редактировать ]Точечные мутации в множественных белках-супрессорах опухолей вызывают рак . Например, точковые мутации аденоматозного полипоза Coli способствуют онкогенезу. [ 9 ] Новый анализ, быстрый параллельный протеолиз (FASTpp) , может помочь быстро выявить специфические дефекты стабильности у отдельных больных раком. [ 10 ]
Нейрофиброматоз
[ редактировать ]Нейрофиброматоз вызывается точечными мутациями нейрофибромина 1. [ 11 ] [ 12 ] или ген нейрофибромина 2 . [ 13 ]
Серповидноклеточная анемия
[ редактировать ]Серповидноклеточная анемия вызвана точечной мутацией в β-глобиновой цепи гемоглобина, приводящей к замене гидрофильной аминокислоты глутаминовой кислоты на гидрофобную аминокислоту валин в шестом положении.
Ген β-глобина находится на коротком плече хромосомы 11. Ассоциация двух субъединиц α-глобина дикого типа с двумя мутантными субъединицами β-глобина образует гемоглобин S (HbS). В условиях низкого содержания кислорода (например, на большой высоте) отсутствие полярной аминокислоты в шестом положении цепи β-глобина способствует нековалентной полимеризации (агрегации) гемоглобина, которая искажает эритроциты в серповидную форму и снижает их эластичность. [ 14 ]
Гемоглобин — это белок, содержащийся в красных кровяных тельцах, который отвечает за транспортировку кислорода по организму. [ 15 ] В состав белка гемоглобина входят две субъединицы: бета-глобины и альфа-глобины . [ 16 ] Бета-гемоглобин создается на основе генетической информации о гене HBB, или «гемоглобин бета», обнаруженном на хромосоме 11p15.5. [ 17 ] Единственная точечная мутация в этой полипептидной цепи длиной 147 аминокислот приводит к заболеванию, известному как серповидноклеточная анемия. [ 18 ] Серповидноклеточная анемия — это аутосомно-рецессивное заболевание, которым страдает 1 из 500 афроамериканцев, и одно из наиболее распространенных заболеваний крови в Соединенных Штатах. [ 17 ] Единственная замена шестой аминокислоты в бета-глобине, глутаминовой кислоты, на валин приводит к деформации эритроцитов. Эти серповидные клетки не могут переносить почти столько же кислорода, как нормальные эритроциты, и они легче захватываются капиллярами, перекрывая кровоснабжение жизненно важных органов. Изменение одного нуклеотида в бета-глобине означает, что даже малейшее усилие со стороны носителя приводит к сильной боли и даже сердечному приступу. Ниже приведена таблица, изображающая первые тринадцать аминокислот в нормальной и аномальной полипептидной цепи серповидноклеточных клеток . [ 18 ]
| АВГ | ГУГ | САС | КУГ | АКУ | CCU | Г А Г | кляп | ААГ | КОНЧИК | GCC | ГУУ | АКУ |
| НАЧИНАТЬ | Вал | Его | Лео | чр | Про | Глу | Глу | Свет | Быть | Земля | Вал | чр |
| АВГ | ГУГ | САС | КУГ | АКУ | CCU | Г У Г | кляп | ААГ | КОНЧИК | GCC | ГУУ | АКУ |
| НАЧИНАТЬ | Вал | Его | Лео | чр | Про | Вал | Глу | Свет | Быть | Земля | Вал | чр |
Болезнь Тея-Сакса
[ редактировать ]Причиной болезни Тея-Сакса является генетический дефект, передающийся от родителей к ребенку. Этот генетический дефект локализован в гене HEXA, который находится на 15-й хромосоме.
Ген HEXA является частью фермента под названием бета-гексозаминидаза А, который играет решающую роль в нервной системе. Этот фермент помогает расщеплять жирное вещество под названием ганглиозид GM2 в нервных клетках. Мутации гена HEXA нарушают активность бета-гексозаминидазы А, предотвращая расщепление жировых веществ. В результате жировые вещества накапливаются до смертельно опасного уровня в головном и спинном мозге. Накопление ганглиозида GM2 вызывает прогрессирующее повреждение нервных клеток. Это является причиной признаков и симптомов болезни Тея-Сакса. [ 19 ]
Повторно-индуцированная точечная мутация
[ редактировать ]В молекулярной биологии точечная мутация , индуцированная повторами , или RIP, представляет собой процесс, посредством которого ДНК накапливает от G : C к A : T мутации перехода . Геномные данные указывают на то, что RIP встречается или возникал у различных грибов. [ 20 ] хотя экспериментальные данные показывают, что RIP активен в Neurospora crassa , [ 21 ] Podospora anserina , [ 22 ] Магнапорте гризеа , [ 23 ] Лептосфаерия пятнистая , [ 24 ] Гибберелла Зеа [ 25 ] и Nectria haematococca . [ 26 ] У Neurospora crassa последовательности, мутированные с помощью RIP, часто метилируются de novo . [ 21 ]
RIP происходит во время половой стадии в гаплоидных ядрах после оплодотворения, но до мейотической репликации ДНК . [ 21 ] У Neurospora crassa повторяющиеся последовательности длиной не менее 400 пар оснований уязвимы для RIP. всего 80% Повторы с идентичностью нуклеотидов также могут подвергаться RIP. Хотя точный механизм распознавания повторов и мутагенеза плохо изучен, RIP приводит к тому, что повторяющиеся последовательности подвергаются множественным переходным мутациям .
Мутации RIP, по-видимому, не ограничиваются повторяющимися последовательностями. Действительно, например, у фитопатогенного гриба L. maculans мутации RIP обнаруживаются в единичных копийных участках, прилегающих к повторяющимся элементам. Эти области представляют собой либо некодирующие области, либо гены, кодирующие небольшие секретируемые белки, включая гены авирулентности. Степень RIP в этих единичных регионах была пропорциональна их близости к повторяющимся элементам. [ 27 ]
Реп и Кистлер предположили, что присутствие высокоповторяющихся областей, содержащих транспозоны, может способствовать мутации резидентных эффекторных генов. [ 28 ] Таким образом, предполагается, что присутствие эффекторных генов в таких регионах способствует их адаптации и диверсификации при воздействии сильного давления отбора. [ 29 ]
Поскольку традиционно считается, что мутация RIP ограничивается повторяющимися областями, а не единичными копиями, Fudal et al. [ 30 ] предположили, что утечка мутации RIP может происходить на относительно небольшом расстоянии от повтора, затронутого RIP. Действительно, об этом сообщалось у N. crassa , при этом утечка RIP была обнаружена в одиночных копий последовательностей на расстоянии по меньшей мере 930 п.н. от границы соседних дублированных последовательностей. [ 31 ] Выяснение механизма обнаружения повторяющихся последовательностей, приводящих к RIP, может позволить понять, как также могут быть затронуты фланкирующие последовательности.
Механизм
[ редактировать ]RIP вызывает G : C в A : T мутации перехода внутри повторов, однако механизм, который обнаруживает повторяющиеся последовательности, неизвестен. RID — единственный известный белок, необходимый для RIP. Это ДНК-метилтрансферазоподобный белок, мутация или нокаут которого приводит к потере RIP. [ 32 ] Делеция гомолога рида у Aspergillus nidulans , dmtA , приводит к потере фертильности. [ 33 ] в то время как делеция гомолога рида у Ascobolus immersens , masc1 , приводит к дефектам фертильности и потере метилирования, индуцированного премейотически (MIP) . [ 34 ]
Последствия
[ редактировать ]Считается, что RIP развился как защитный механизм против мобильных элементов , которые напоминают паразитов , проникая в геном и размножаясь в нем. RIP создает множественные миссенс- и нонсенс-мутации в кодирующей последовательности. Эта гипермутация GC в AT в повторяющихся последовательностях устраняет функциональные генные продукты последовательности (если они вообще были). Кроме того, многие С-несущие нуклеотиды метилируются , что снижает транскрипцию.
Использование в молекулярной биологии
[ редактировать ]Поскольку RIP настолько эффективен при обнаружении и мутации повторов, биологи-грибки часто используют его как инструмент мутагенеза . Вторая копия однокопийного гена сначала трансформируется в геном . Затем гриб должен спариться и пройти половой цикл, чтобы активировать механизм RIP. Даже в результате одного события оплодотворения возникает множество различных мутаций внутри дуплицированного гена, так что инактивированные аллели, обычно из-за нонсенс-мутаций , а также аллели, содержащие миссенс-мутации . могут быть получены [ 35 ]
История
[ редактировать ]Процесс клеточного воспроизводства мейоза был открыт Оскаром Гертвигом в 1876 году. Митоз был открыт несколькими годами позже, в 1882 году, Вальтером Флеммингом .
Хертвиг изучал морских ежей и заметил, что каждая яйцеклетка содержала одно ядро до оплодотворения и два ядра после. Это открытие доказало, что один сперматозоид может оплодотворить яйцеклетку, а значит, доказало процесс мейоза. Герман Фоль продолжил исследования Хертвига, проверив эффекты инъекции нескольких сперматозоидов в яйцеклетку, и обнаружил, что этот процесс не работает более чем с одним сперматозоидом. [ 36 ]
Флемминг начал свои исследования деления клеток в 1868 году. В этот период изучение клеток становилось все более популярной темой. К 1873 году Шнайдер уже начал описывать этапы деления клеток. Флемминг развил это описание в 1874 и 1875 годах, более подробно объяснив эти этапы. Он также опроверг выводы Шнайдера о том, что ядро разделилось на стержнеобразные структуры, предположив, что ядро на самом деле разделилось на нити, которые, в свою очередь, разделились. Флемминг пришел к выводу, что клетки реплицируются посредством клеточного деления, а точнее митоза. [ 37 ]
Мэтью Мезельсону и Франклину Сталю приписывают открытие репликации ДНК . Уотсон и Крик признали, что структура ДНК действительно указывает на наличие той или иной формы процесса репликации. Однако до Уотсона и Крика по этому аспекту ДНК не проводилось много исследований. Люди рассмотрели все возможные методы определения процесса репликации ДНК, но ни один из них не оказался успешным до Мезельсона и Сталя. Мезельсон и Шталь ввели тяжелый изотоп в ДНК и проследили его распространение. Благодаря этому эксперименту Мезельсон и Шталь смогли доказать, что ДНК размножается полуконсервативно. [ 38 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Точечная мутация» . Биологический словарь . 22 ноября 2016 года . Проверено 17 мая 2019 г.
- ^ Фриз, Эрнст (апрель 1959 г.). «Разница между спонтанными мутациями фага Т4 и мутациями, индуцированными базовым аналогом» . Учеб. Натл. акад. наук. США . 45 (4): 622–33. Бибкод : 1959PNAS...45..622F . дои : 10.1073/pnas.45.4.622 . ПМК 222607 . ПМИД 16590424 .
- ^ Фриз, Эрнст (1959). «Специфическое мутагенное действие базовых аналогов на фаг Т4». Дж. Мол. Биол . 1 (2): 87–105. дои : 10.1016/S0022-2836(59)80038-3 .
- ^ Перейти обратно: а б с д «Генетический учебник» . Архивировано из оригинала 11 апреля 2005 года.
- ^ Дэвис Х., Бигнелл Г.Р., Кокс С. и др. (июнь 2002 г.). «Мутации гена BRAF при раке человека» (PDF) . Природа . 417 (6892): 949–54. Бибкод : 2002Natur.417..949D . дои : 10.1038/nature00766 . ПМИД 12068308 . S2CID 3071547 .
- ^ Хоймейкерс Дж. Х. (май 2001 г.). «Механизмы поддержания генома для предотвращения рака». Природа . 411 (6835): 366–74. Бибкод : 2001Natur.411..366H . дои : 10.1038/35077232 . ПМИД 11357144 . S2CID 4337913 .
- ^ Ли, Минхуэй; Гончаренко, Александр; Панченко, Анна Р. (2017). «Аннотирование мутационных эффектов на белки и белковые взаимодействия: разработка новых и пересмотр существующих протоколов». Протеомика . Методы молекулярной биологии. Том. 1550. стр. 235–260. дои : 10.1007/978-1-4939-6747-6_17 . ISBN 978-1-4939-6745-2 . ISSN 1940-6029 . ПМК 5388446 . ПМИД 28188534 .
- ^ «Краткий путь к персонализированной медицине» . Новости генной инженерии и биотехнологии. 18 июня 2008 г.
- ^ Минде Д.П., Анвариан З., Рюдигер С.Г., Морис М.М. (2011). «Беспорядок: как миссенс-мутации в белке-супрессоре опухолей APC приводят к раку?» . Мол. Рак . 10 :101. дои : 10.1186/1476-4598-10-101 . ПМК 3170638 . ПМИД 21859464 .
- ^ Минде Д.П., Морис М.М., Рюдигер С.Г. (2012). «Определение биофизической стабильности белков в лизатах с помощью анализа быстрого протеолиза, FASTpp» . ПЛОС ОДИН . 7 (10): е46147. Бибкод : 2012PLoSO...746147M . дои : 10.1371/journal.pone.0046147 . ПМЦ 3463568 . ПМИД 23056252 .
- ^ Серра, Э; Арс, Э; Равелла, А; Санчес, А; Пуиг, С; Розенбаум, Т; Эстивилл, X; Лазаро, К. (2001). «Мутационный спектр соматического NF1 в доброкачественных нейрофибромах: дефекты сплайсинга МРНК распространены среди точечных мутаций». Генетика человека . 108 (5): 416–29. дои : 10.1007/s004390100514 . ПМИД 11409870 . S2CID 2136834 .
- ^ Уист, В; Эйзенбарт, я; Шмегнер, К; Крона, В; Предположим, Г (2003). «Спектры соматических мутаций NF1 в семье с нейрофиброматозом 1 типа: к теории генетических модификаторов» . Человеческая мутация . 22 (6): 423–7. дои : 10.1002/humu.10272 . ПМИД 14635100 . S2CID 22140210 .
- ^ Мохьюддин, А; Нири, WJ; Уоллес, А; Ву, CL; Перселл, С; Рид, Х; Рамсден, RT; Читай, А; Черный, Г; Эванс, генеральный директор (2002). «Молекулярно-генетический анализ гена NF2 у молодых пациентов с односторонними вестибулярными шванномами» . Журнал медицинской генетики . 39 (5): 315–22. дои : 10.1136/jmg.39.5.315 . ПМК 1735110 . ПМИД 12011146 .
- ^ Гены и болезни . Национальный центр биотехнологической информации (США). 29 сентября 1998 г. - через PubMed.
- ^ Ся CC (январь 1998 г.). «Дыхательная функция гемоглобина». Н. англ. Дж. Мед . 338 (4): 239–47. дои : 10.1056/NEJM199801223380407 . ПМИД 9435331 .
- ^ «ГББ — Гемоглобин Бета» . Домашний справочник по генетике . Национальная медицинская библиотека.
- ^ Перейти обратно: а б «Анемия, серповидноклеточная» . Гены и болезни . Bethesda MD: Национальный центр биотехнологической информации. 1998. НБК22183.
- ^ Перейти обратно: а б Клэнси С. (2008). «Генетическая мутация» . Природное образование . 1 (1): 187.
- ^ эМедТВ. «Причины Тея-Сакса» . Архивировано из оригинала 6 августа 2020 года . Проверено 28 декабря 2011 г.
- ^ Клаттербак Эй Джей (2011). «Геномные доказательства точковой мутации, индуцированной повторами (RIP) у нитчатых аскомицетов». Грибковая генетика биол . 48 (3): 306–26. дои : 10.1016/j.fgb.2010.09.002 . ПМИД 20854921 .
- ^ Перейти обратно: а б с Селкер ЕС, Камбарери Э.Б., Йенсен Б.К., Хаак К.Р. (декабрь 1987 г.). «Перестройка дублированной ДНК в специализированных клетках Neurospora». Клетка . 51 (5): 741–752. дои : 10.1016/0092-8674(87)90097-3 . ПМИД 2960455 . S2CID 23036409 .
- ^ Грая Ф, Леспине О, Рембо Б, Декар-Шаблат М, Коппин Э, Пикард М (май 2001 г.). «Контроль качества генома: RIP (повторно-индуцированная точечная мутация) приходит к подоспорам» . Мол Микробиол . 40 (3): 586–595. дои : 10.1046/j.1365-2958.2001.02367.x . ПМИД 11359565 . S2CID 25096512 .
- ^ Икеда К., Накаяшики Х., Катаока Т., Тамба Х., Хасимото Ю., Тоса Ю., Маяма С. (сентябрь 2002 г.). «Повторно-индуцированная точечная мутация (RIP) у Magnaporthe grisea : последствия для ее полового цикла в контексте естественного поля» . Мол Микробиол . 45 (5): 1355–1364. дои : 10.1046/j.1365-2958.2002.03101.x . ПМИД 12207702 .
- ^ Иднурм А., Хоулетт Б.Дж. (июнь 2003 г.). могут возникать повторные точковые мутации «Анализ мутантов с потерей патогенности показывает, что у Dothideomycete Leptosphaeria maculans ». Грибковая генетика биол . 39 (1): 31–37. дои : 10.1016/S1087-1845(02)00588-1 . ПМИД 12742061 .
- ^ Куомо Калифорния, Гюльденер Ю, Сюй Дж.Р., Трейл Ф, Терджен Б.Г., Ди Пьетро А., Уолтон Дж.Д., Ма Л.Дж. и др. (сентябрь 2007 г.). « Геном Fusarium graminearum обнаруживает связь между локализованным полиморфизмом и специализацией патогена». Наука . 317 (5843): 1400–2. Бибкод : 2007Sci...317.1400C . дои : 10.1126/science.1143708 . ПМИД 17823352 . S2CID 11080216 .
- ^ Коулман Дж.Дж., Раунсли С.Д., Родригес-Каррес М., Куо А., Васманн К.С., Гримвуд Дж., Шмутц Дж. и др. (август 2009 г.). «Геном Nectria haematococca : вклад дополнительных хромосом в расширение генов» . ПЛОС Генет . 5 (8): e1000618. дои : 10.1371/journal.pgen.1000618 . ПМЦ 2725324 . ПМИД 19714214 .
- ^ Ван де Воу А.П., Козинсен А.Дж., Хане Дж.К. и др. (2010). «На эволюцию связанных эффекторов авирулентности у Leptosphaeria maculans влияет геномная среда и воздействие генов устойчивости в растениях-хозяевах» . ПЛОС Патог . 6 (11): е1001180. дои : 10.1371/journal.ppat.1001180 . ПМЦ 2973834 . ПМИД 21079787 .
- ^ Представитель М, Кистлер ХК (август 2010 г.). «Геномная организация патогенности растений видов Fusarium» . Курс. Мнение. Растительная биол . 13 (4): 420–6. Бибкод : 2010COPB...13..420R . дои : 10.1016/j.pbi.2010.04.004 . ПМИД 20471307 . Архивировано из оригинала 2 июня 2020 года . Проверено 29 декабря 2018 г.
- ^ Фарман М.Л. (август 2007 г.). «Теломеры в рисовом грибке Magnaporthe oryzae : мир конца, каким мы его знаем» . ФЭМС Микробиол. Летт . 273 (2): 125–32. дои : 10.1111/j.1574-6968.2007.00812.x . ПМИД 17610516 .
- ^ Фудал И., Росс С., Брун Х. и др. (август 2009 г.). «Повторно-индуцированная точечная мутация (RIP) как альтернативный механизм эволюции вирулентности у Leptosphaeria maculans » . Мол. Взаимодействие с растительными микробами . 22 (8): 932–41. дои : 10.1094/MPMI-22-8-0932 . ПМИД 19589069 .
- ^ Ирелан Дж.Т., Хагеманн А.Т., Селькер ЕС (декабрь 1994 г.). «Высокочастотная точечная мутация, индуцированная повторами (RIP), не связана с эффективной рекомбинацией у Neurospora» . Генетика . 138 (4): 1093–103. дои : 10.1093/генетика/138.4.1093 . ПМК 1206250 . ПМИД 7896093 .
- ^ Фрайтаг М., Уильямс Р.Л., Коте ГО, Селкер ЕС (2002). «Гомолог цитозинметилтрансферазы необходим для повторной точковой мутации у Neurospora crassa » . Proc Natl Acad Sci США . 99 (13): 8802–7. Бибкод : 2002PNAS...99.8802F . дои : 10.1073/pnas.132212899 . ПМЦ 124379 . ПМИД 12072568 .
- ^ Ли Д.В., Фрайтаг М., Селкер ЕС, Арамайо Р. (2008). «Гомолог цитозинметилтрансферазы необходим для полового развития Aspergillus nidulans» . ПЛОС ОДИН . 3 (6): e2531. Бибкод : 2008PLoSO...3.2531L . дои : 10.1371/journal.pone.0002531 . ПМК 2432034 . ПМИД 18575630 .
- ^ Маланьяк Ф., Вендель Б., Гойон С., Фожерон Г., Циклер Д., Россиньол Дж.Л. и др. (1997). «Ген, необходимый для метилирования и развития de novo у Ascobolus, открывает новый тип структуры эукариотической ДНК-метилтрансферазы» . Клетка . 91 (2): 281–90. дои : 10.1016/S0092-8674(00)80410-9 . ПМИД 9346245 . S2CID 14143830 .
- ^ Селькер ЕС (1990). «Премейотическая нестабильность повторяющихся последовательностей Neurospora crassa». Анну преподобный Жене . 24 : 579–613. дои : 10.1146/annurev.ge.24.120190.003051 . ПМИД 2150906 .
- ^ Барбьери, Марчелло (2003). «Проблема поколения» . Органические коды: введение в семантическую биологию . Издательство Кембриджского университета. п. 13. ISBN 978-0-521-53100-9 .
- ^ Павелец Н. (январь 2001 г.). «Вальтер Флемминг: пионер исследования митоза». Нат. Преподобный мол. Клеточная Биол . 2 (1): 72–5. дои : 10.1038/35048077 . ПМИД 11413469 . S2CID 205011982 .
- ^ Холмс, Фредерик Лоуренс (2001). Мезельсон, Шталь и репликация ДНК: история «самого красивого эксперимента в биологии» . Издательство Йельского университета. ISBN 978-0-300-08540-2 .
Внешние ссылки
[ редактировать ]- Точка + мутация Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)