Регионы с высоким содержанием питательных веществ и низким содержанием хлорофилла
Районы с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC) — это регионы океана, где численность фитопланктона низкая и довольно постоянная, несмотря на наличие макронутриентов . Фитопланктон нуждается в наборе питательных веществ для клеточного функционирования. Макронутриенты (например, нитраты , фосфаты , кремниевая кислота ) обычно присутствуют в больших количествах в поверхностных водах океана и являются типичными компонентами обычных садовых удобрений. Микронутриенты (например, железо , цинк , кобальт ) обычно доступны в меньших количествах и включают следы металлов . Макронутриенты обычно доступны в миллимолярных концентрациях , тогда как микроэлементы обычно доступны в микро- и наномолярных концентрациях. В целом азот, как правило, является ограничивающим питательным веществом океана , но в регионах HNLC он никогда не истощается значительно. [1] [2] Вместо этого эти регионы, как правило, ограничены низкими концентрациями метаболизируемого железа. [1] Железо является важнейшим микроэлементом фитопланктона, необходимым для ферментативного катализа и транспорта электронов . [3] [4]
Между 1930-ми и 1980-ми годами была выдвинута гипотеза, что железо является лимитирующим микроэлементом океана, но не было достаточных методов надежного обнаружения железа в морской воде, чтобы подтвердить эту гипотезу. [5] В 1989 году были обнаружены высокие концентрации богатых железом отложений в прибрежных водах залива Аляска. [6] Однако морские воды имели более низкую концентрацию железа и более низкую продуктивность, несмотря на доступность макроэлементов для роста фитопланктона. [6] Эта закономерность наблюдалась и в других океанических регионах и привела к названию трех основных зон HNLC: Северная часть Тихого океана , Экваториальная часть Тихого океана и Южный океан . [1] [2]
Открытие регионов HNLC способствовало научным дебатам об этике и эффективности экспериментов по внесению железных удобрений, которые пытаются снизить содержание углекислого газа в атмосфере путем стимулирования фотосинтеза на уровне поверхности. Это также привело к развитию таких гипотез, как контроль выпаса, который предполагает, что регионы HNLC формируются, частично, в результате выпаса фитопланктона (например, динофлагеллят , инфузорий ) более мелкими организмами (например, протистами ).
Первичное производство
[ редактировать ]

Первичное производство — это процесс, при котором автотрофы используют свет для преобразования углерода из водного диоксида углерода в сахар для роста клеток. [7] Свет обеспечивает энергию для процесса фотосинтеза , а питательные вещества включаются в органический материал . Для осуществления фотосинтеза макроэлементы, такие как нитраты и фосфаты, должны быть доступны в достаточных соотношениях и в биодоступных формах для биологического использования. Молекулярное соотношение 106(Углерод):16(Азот):1(Фосфор) было выведено Редфилдом, Кетчемом и Ричардсом (RKR) и известно как соотношение Редфилда . [8] Фотосинтез (прямой) и дыхание (обратный) описываются уравнением:

Фотосинтез может быть ограничен дефицитом определенных макроэлементов. Однако в северной, экваториальной части Тихого океана и Южном океане макронутриенты обнаруживаются в достаточных соотношениях, количествах и биодоступных формах, чтобы поддерживать более высокие уровни первичного производства, чем обнаружены. Наличие макронутриентов в регионах HNLC в сочетании с низкими запасами фитопланктона позволяет предположить, что какой-то другой биогеохимический процесс ограничивает рост фитопланктона. [7]
Поскольку первичная продукция и биомасса фитопланктона в настоящее время не могут быть измерены во всех океанских бассейнах, ученые используют хлорофилл α в качестве показателя первичной продукции. Современные спутниковые наблюдения контролируют и отслеживают глобальное содержание хлорофилла α в океане с помощью дистанционного зондирования . Более высокие концентрации хлорофилла обычно указывают на области повышенного первичного производства, и наоборот, более низкие уровни хлорофилла указывают на низкое первичное производство. Сочетание низкого содержания хлорофилла и высокой доступности макронутриентов является причиной того, что эти регионы считаются «высоким содержанием питательных веществ и низким содержанием хлорофилла».
В дополнение к макроэлементам, необходимым для синтеза органических веществ, фитопланктону необходимы микроэлементы, такие как микроэлементы, для клеточных функций. [7] Доступность микроэлементов может ограничивать первичное производство, поскольку микроэлементы иногда ограничивают питательные вещества. Было установлено, что железо является основным лимитирующим микроэлементом в провинциях HNLC. [5] Недавние исследования показали, что цинк и кобальт могут быть вторичными и/или сопутствующими микроэлементами. [10] [11]
Глобальное распространение
[ редактировать ]Общие характеристики
[ редактировать ]Регионы HNLC покрывают 20% мирового океана и характеризуются различными физическими, химическими и биологическими особенностями. Эти поверхностные воды имеют ежегодно меняющуюся, но относительно высокую концентрацию макронутриентов по сравнению с другими океаническими провинциями. [5] Хотя HNLC в общих чертах описывает биогеохимические тенденции этих крупных океанических регионов, во всех трех зонах наблюдается сезонное цветение фитопланктона в ответ на глобальные атмосферные закономерности. В среднем регионы HNLC, как правило, ограничены ростом железа и, в разной степени, цинка. [11] [12] Это ограничение содержания микроэлементов приводит к образованию сообществ фитопланктона меньшего размера. По сравнению с более продуктивными регионами океана, зоны HNLC имеют более высокое соотношение кремниевой кислоты к нитратам, поскольку более крупные диатомовые водоросли , которым требуется кремниевая кислота для изготовления опаловых кремнеземных оболочек, менее распространены. [10] [11] [12] В отличие от Южного океана и северной части Тихого океана, в экваториальной части Тихого океана наблюдается временное наличие силикатов, что приводит к масштабному сезонному цветению диатомей. [13] [14]
Распределение микроэлементов и относительное содержание макронутриентов отражается на структуре планктонного сообщества. Например, отбор фитопланктона с высоким соотношением площади поверхности к объему приводит к тому, что в регионах HNLC доминируют нано- и пикопланктон. Такое соотношение позволяет оптимально использовать доступные растворенные питательные вещества. Более крупный фитопланктон, такой как диатомовые водоросли, не может энергетически поддерживать себя в этих регионах. Обычный пикопланктон в этих регионах включает такие роды, как прохлорококк (обычно не встречающийся в северной части Тихого океана), синехококк и различные эукариоты . Пасущиеся протисты, вероятно, контролируют численность и распределение этого небольшого фитопланктона. [15] [16]
Как правило, более низкая чистая первичная продукция в зонах HNLC приводит к более низкому биологическому сокращению атмосферного углекислого газа, и поэтому эти регионы обычно считаются чистым источником углекислого газа в атмосферу. [14] Области HNLC представляют интерес для геоинженеров и некоторых представителей научного сообщества, которые считают, что удобрение больших участков этих вод железом потенциально может снизить растворенный углекислый газ и компенсировать увеличение антропогенных выбросов углерода. [6] Анализ данных кернов антарктического льда за последний миллион лет показывает корреляцию между высоким уровнем пыли и низкой температурой, указывая на то, что добавление диффузной богатой железом пыли в море было естественным усилителем похолодания климата. [17]
Северная часть Тихого океана
[ редактировать ]
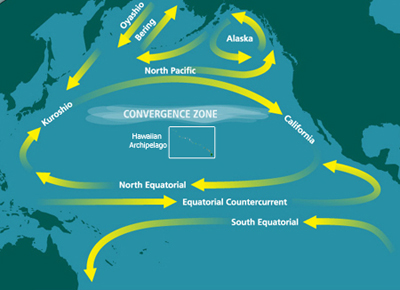
Открытие и наименование первого региона HNLC, Северная часть Тихого океана , было официально оформлено в основополагающей статье, опубликованной в 1988 году. [6] Исследование пришло к выводу, что в поверхностных водах восточной части северной части Тихого океана обычно преобладает пикопланктон, несмотря на относительное изобилие макронутриентов. [6] Другими словами, более крупный фитопланктон, такой как диатомовые водоросли, которые процветают в богатых питательными веществами водах, не был обнаружен. Вместо этого поверхностные воды были изобилуют более мелким пико- и нанопланктоном. [6] На основании лабораторных экспериментов с питательными веществами была выдвинута гипотеза, что железо является ключевым лимитирующим микроэлементом. [6]
– Тихий океан самый крупный и древний водоем на Земле. Северная часть Тихого океана характеризуется общим вращением северо-тихоокеанского круговорота по часовой стрелке , которое приводится в движение пассатными ветрами . Пространственные вариации пассатов приводят к более низким температурам воздуха в западной части северной части Тихого океана и к более умеренным температурам воздуха в восточной части северной части Тихого океана (т. е. в субарктической части Тихого океана). [18] Железо доставляется в северную часть Тихого океана пыльными бурями, возникающими в Азии и на Аляске, а также богатыми железом водами, переносимыми с окраины континента, иногда вихрями, такими как Хайда Вихри . [19] [20]
Однако концентрации железа меняются в течение года. Океанские течения обусловлены сезонными атмосферными явлениями, которые переносят железо с Курило-Камчатской окраины в западную субарктическую часть Тихого океана. Такое введение железа обеспечивает подземный запас микроэлементов, которые могут быть использованы первичными продуцентами при подъеме более глубоких вод на поверхность. [21] Глубина морского дна также может стимулировать цветение фитопланктона в регионах HNLC, поскольку железо диффундирует с морского дна и снижает дефицит железа на мелководье. [22] Исследования, проведенные в заливе Аляска, показали, что в районах с мелководьем, таких как южный шельф Аляски, цветение фитопланктона более интенсивное, чем в прибрежных водах. [22] Вулканический пепел от извержения вулкана Касаточи в августе 2008 года стал примером естественного обогащения железом в северо-восточной части Тихого океана. [23] Этот регион был удобрен дождями из вулканической пыли, содержащей растворимое железо. В последующие дни цветение фитопланктона было видно из космоса. [23]
Ограничения в концентрации микроэлементов в северной части Тихого океана ограничивают цветение диатомей в течение всего года. [24] Несмотря на то, что северная часть Тихого океана является регионом HNLC, она производит и экспортирует во внутреннюю часть океана относительно большое количество твердых частиц биогенного кремнезема по сравнению с Северной Атлантикой, что способствует значительному росту диатомовых водорослей. [24]
Экваториальная часть Тихого океана
[ редактировать ]Экваториальная часть Тихого океана — океаническая провинция, характеризующаяся почти круглогодичным апвеллингом из-за сближения пассатов с северо-востока и юго-востока в зоне внутритропической конвергенции . Экваториальная часть Тихого океана занимает почти половину окружности Земли и играет важную роль в глобальной морской первичной продукции . [25] Новое производство — это термин, используемый в биологической океанографии для описания способа переработки азота в океане. [18] В регионах повышенной новой продукции нитраты из афотической зоны попадают в поверхностные воды, пополняя запасы поверхностных нитратов. Несмотря на наличие азота в водах экваториальной части Тихого океана, первичная продукция и наблюдаемая поверхностная биомасса океана значительно ниже по сравнению с другими крупными районами апвеллинга океана. [26] Таким образом, экваториальная часть Тихого океана считается одним из трех основных регионов HNLC.
Как и другие крупные провинции HNLC, экваториальная часть Тихого океана считается ограниченной по питательным веществам из-за отсутствия микроэлементов, таких как железо. Экваториальная часть Тихого океана получает примерно в 7-10 раз больше железа в результате апвеллинга Экваториального подводного течения (EUC), чем из-за оседания атмосферной пыли. [27] Реконструкция климата ледниковых периодов с использованием косвенных данных отложений показала, что экваториальная часть Тихого океана могла быть в 2,5 раза более продуктивной, чем современный экваториальный океан. [27] В эти ледниковые периоды экваториальная часть Тихого океана увеличила экспорт новой морской продукции, [27] таким образом обеспечивая сток углекислого газа из атмосферы. Наука палеоокеанография пытается понять взаимодействие ледниковых циклов с динамикой океана. Палеоокеанографы в настоящее время оспаривают гипотезу эоловой пыли, которая предполагает, что атмосферный перенос богатой железом пыли из Центральной и Южной Америки контролирует интенсивность первичной продукции в экваториальной части Тихого океана. [27] Одно исследование предполагает, что, поскольку апвеллинг EUC обеспечивает большую часть биодоступного железа экваториальным поверхностным водам, единственным способом обратить вспять условия HNLC является усиление апвеллинга. [27] [28] Другими словами, усиление регионального апвеллинга, а не отложение богатой железом атмосферной пыли, может объяснить, почему в этом регионе наблюдается более высокая первичная продуктивность во время ледниковых периодов.
По сравнению с северной частью Тихого океана и Южным океаном, воды экваториальной части Тихого океана имеют относительно низкий уровень биогенного кремнезема и, следовательно, не поддерживают значительные постоянные запасы диатомовых водорослей. [14] Пикопланктон является наиболее многочисленным первичным морским продуцентом в этих регионах, главным образом, благодаря его способности ассимилировать низкие концентрации микроэлементов. [14] Различные сообщества фитопланктона в экваториальной части Тихого океана выпасаются с той же скоростью, что и скорость их роста, что еще больше ограничивает первичное производство. [28]
В настоящее время нет единого мнения относительно того, какая из двух основных гипотез (выпас или микроэлементы) контролирует производство в этих экваториальных водах. Вполне вероятно, что ограничения по следам металлов отдают предпочтение мелкоклеточным организмам, что тем самым увеличивает нагрузку на выпас простейших. [28] Хотя экваториальная часть Тихого океана сохраняет характеристики HNLC, продуктивность временами может быть высокой. Продуктивность приводит к обилию морских птиц, таких как буревестники, вблизи места схождения субтропических вод и экваториального «холодного языка». В экваториальной части Тихого океана находится крупнейший в мире промысел желтоперого тунца. [18] и является домом для пятнистого дельфина .
Южный океан
[ редактировать ]
Южный океан является крупнейшим регионом HNLC в мировом океане. Поверхностные воды Южного океана широко известны как богатые макроэлементами, несмотря на низкие запасы фитопланктона. [29] [30] [31] Железо, отложившееся в Северной Атлантике, попадает в глубоководные воды Северной Атлантики и переносится в Южный океан посредством термохалинной циркуляции . [32] В конечном итоге смешиваясь с циркумполярными водами Антарктики , апвеллинг доставляет железо и макроэлементы в поверхностные воды Южного океана. Таким образом, поступления железа и первичное производство в Южном океане чувствительны к богатой железом сахарской пыли, отложившейся над Атлантикой. Из-за низкого поступления атмосферной пыли непосредственно в поверхностные воды Южного океана, [33] [34] Концентрация хлорофилла α низкая. Доступность света в Южном океане резко меняется в зависимости от сезона, но, похоже, это не является существенным ограничением для роста фитопланктона. [3]
Макронутриенты, присутствующие в поверхностных водах Южного океана, поступают из поднятых глубинных вод. Хотя такие микроэлементы, как цинк и кобальт, возможно, могут совместно ограничивать рост фитопланктона в Южном океане, железо, по-видимому, является критическим лимитирующим микроэлементом. [4] В некоторых регионах Южного океана наблюдается достаточная концентрация биодоступного железа и макронутриентов, однако рост фитопланктона ограничен. Гидрографические исследования [35] [36] и исследования региона Южного пролива Дрейка [37] наблюдали это явление вокруг островов Крозе , Кергелен , Южной Георгии и Южных Сандвичевых островов . [37] [38] Эти районы примыкают к шельфовым районам Антарктиды и островам Южного океана. Считается, что микроэлементы, необходимые для роста водорослей, поставляются с самих полок. [37] За исключением районов, близких к антарктическому шельфу, дефицит микроэлементов серьезно ограничивает продуктивность в Южном океане.
Доступность железа – не единственный регулятор продуктивности и биомассы фитопланктона. [39] [40] Считается, что в Южном океане преобладающие низкие температуры оказывают негативное влияние на темпы роста фитопланктона. [40] Скорость роста фитопланктона очень интенсивна и недолговечна на открытых территориях, окруженных морским льдом и в зонах постоянного морского льда . выпас травоядных животных, таких как криль, копеподы и сальпы, Считается, что подавляет запасы фитопланктона. В отличие от открытых вод Южного океана, выпас вдоль окраин континентального шельфа невелик, поэтому большая часть не потребляемого фитопланктона опускается на морское дно, обеспечивая питательными веществами донные организмы. [39]
Гипотезы
[ редактировать ]Учитывая удаленность районов HNLC, ученые объединили данные моделирования и наблюдений, чтобы изучить ограничения на первичное производство. Объединение этих двух источников данных позволяет сравнивать северную часть Тихого океана, экваториальную часть Тихого океана и Южный океан. Двумя текущими объяснениями глобальных регионов HNLC являются ограничения роста из-за доступности железа и контроля выпаса фитопланктона.
Железная гипотеза
[ редактировать ]В 1988 году Джон Мартин подтвердил гипотезу о том, что железо ограничивает цветение и скорость роста фитопланктона в северной части Тихого океана. Его работа была экстраполирована на другие регионы HNLC благодаря доказательствам, связывающим низкую концентрацию железа на поверхности с низким содержанием хлорофилла. [6] В ответ на эксперименты по внесению железных удобрений серьезные реакции фитопланктона, такие как снижение концентрации питательных веществ на поверхности и повышение биологической активности . (IronEx, SOIREE, SEEDS и т. д.) в районах HNLC наблюдались [41] [42] [43] [44] [45]
Исследования по оплодотворению железом, проводимые с повторяющимися интервалами в течение недели, дали больший биологический ответ, чем однократное оплодотворение. [42] [44] [46] Размер биологической реакции обычно зависит от биологических, химических и физических характеристик участка. Считается, что в экваториальной и северной части Тихого океана кремнезем ограничивает дополнительное производство после внесения железных удобрений, в то время как свет ограничивает дополнительное производство в Южном океане. [42] Местный, более мелкий фитопланктон первым отреагировал на повышенное содержание железа, но его быстро вытеснил более крупный прибрежный фитопланктон, такой как диатомовые водоросли. [44] [47] [48] [49] Масштабная реакция цветения и смена сообществ привели к экологическим опасениям по поводу внесения удобрений на больших участках регионов HNLC. Одно исследование предполагает, что диатомеи растут преимущественно во время экспериментов по оплодотворению. Некоторые диатомовые водоросли, такие как псевдоницсия , выделяют нейротоксин домоевую кислоту , отравляющую пасущуюся рыбу. [48] Если диатомовые водоросли растут преимущественно во время экспериментов по оплодотворению железом, длительное внесение удобрений может усилить отравление домоевой кислотой в морской пищевой сети вблизи удобренных участков. [48]

Эолова пыль
[ редактировать ]Железо попадает в отдаленные регионы HNLC двумя основными способами: подъемом богатой питательными веществами воды и осаждением атмосферной пыли . Железо необходимо пополнять часто и в биодоступных формах из-за его нерастворимости , быстрого усвоения биологическими системами и сродства связывания с лигандами . [50] [51] Отложение пыли может не привести к цветению фитопланктона, если оседающая пыль не будет иметь правильную биодоступную форму железа. Кроме того, железо должно откладываться в течение продуктивных сезонов и соответствовать соответствующему соотношению RKR поверхностных питательных веществ. [19] [52] Эоловая пыль оказывает большее влияние на регионы HNLC Северного полушария, поскольку большая часть суши способствует большему осаждению пыли. [53] Из-за изоляции Южного океана от суши апвеллинг, связанный с вихревой диффузией, обеспечивает железо в регионы HNLC. [54]
Гипотеза контроля выпаса скота
[ редактировать ]Гипотеза выпаса, сформулированная Джоном Уолшем в 1976 году, утверждает, что выпас гетеротрофов подавляет первичную продуктивность в районах с высокой концентрацией питательных веществ. [41] [55] Хищничество микрозоопланктона в первую очередь приводит к гибели фитопланктона в регионах HNLC. Выпас более крупного зоопланктона и адвективное перемешивание также несут ответственность за небольшую долю потерь фитопланктонных сообществ. [6] [56] [57] Постоянный выпас ограничивает фитопланктон низким, постоянным запасом. Некоторые ученые полагают, что без такого давления на выпас скота мелкий фитопланктон будет вызывать цветение, несмотря на истощение микроэлементов, поскольку более мелкий фитопланктон обычно требует меньше железа и может поглощать питательные вещества медленнее. [50] [56]
Современный вид
[ редактировать ]Текущий научный консенсус согласен с тем, что в районах HNLC отсутствует высокая продуктивность из-за сочетания железа и физиологических ограничений, давления выпаса и физических воздействий. [2] [6] [43] [49] [56] [58] Степень, в которой каждый фактор способствует снижению производства, может различаться в каждом регионе HNLC. Ограничение железа позволяет более мелкому и более экономному по железу фитопланктону расти быстрыми темпами, в то время как выпас микрозоопланктона поддерживает стабильные запасы этого более мелкого фитопланктона. [6] [51] [56] Как только микроэлементы станут доступными, выпас может ограничить размер цветения. [41] [43] [44] [46] [49] Дополнительные ограничения на микроэлементы, такие как микроэлементы, такие как цинк или кобальт, могут подавлять цветение фитопланктона. [12] Турбулентное перемешивание в регионах HNLC более высоких широт (северная часть Тихого океана и Южный океан) может смешивать фитопланктон ниже критической глубины, необходимой для роста сообщества. [41]
Геоинженерия регионов HNLC
[ редактировать ]Теория
[ редактировать ]
Поскольку прошлые эксперименты по удобрению железом привели к массовому цветению фитопланктона, некоторые полагают, что следует провести крупномасштабные эксперименты по удобрению океана для поглощения неорганического антропогенного углекислого газа в форме твердых частиц органического углерода . Внесение удобрений будет стимулировать биологическую продуктивность, что приведет к уменьшению количества неорганического поверхностного диоксида углерода на удобренном участке. Затем цветение заглохнет и, предположительно, опустится в глубины океана, унося большую часть поглощенного углекислого газа на морское дно и изолируя его от кратковременного углеродного цикла в глубинах океана или океанских отложениях. [46] [59] [60] [61] [62]
Эффективность и действенность
[ редактировать ]Чтобы эффективно удалить антропогенный углерод из атмосферы, внесение железа должно привести к значительному удалению твердых частиц углерода с поверхности океана и транспортировке его в глубины океана. [42] [43] [60] [61] По оценкам различных исследований, менее 7-10% углерода, поглощенного во время цветения, будет секвестрировано. [63] и только снижение содержания углекислого газа в атмосфере на 15-25 частей на миллион приведет к устойчивому глобальному удобрению железом. [7] [60] Количество удаленного углекислого газа может быть компенсировано затратами на топливо для приобретения, транспортировки и выброса значительных количеств железа в отдаленные регионы HNLC. [61]
Существует множество экологических проблем, связанных с крупномасштабным внесением железных удобрений. Хотя цветение можно изучать и отслеживать, ученые до сих пор не знают, попадает ли дополнительная продукция в пищевую цепочку или попадает в глубокий океан после отмирания цветения. [42] [43] Даже если углерод экспортируется на глубину, органические вещества, выпадающие в результате дождя, могут вдыхаться, что потенциально создает бескислородные зоны в середине столба или вызывает подкисление глубоководных океанских вод. [61] [64] Выраженный сдвиг сообщества в сторону диатомовых водорослей наблюдался во время оплодотворения, и до сих пор неясно, имеет ли изменение видового состава какие-либо долгосрочные последствия для окружающей среды. [48] [61]
Энергетические ресурсы
[ редактировать ]Нижеследующее является полностью теоретическим. Тестирование потребуется для определения осуществимости, оптимальной концентрации железа на единицу площади, секвестрации углерода по площади с течением времени, потребности в других микроэлементах, количества энергии, необходимой для поддержания такой системы, и потенциального количества энергии, производимой системой. Эта система учитывает экономическую целесообразность (рентабельность биотопливных продуктов и углеродных кредитов) и управление рисками.
Рост
[ редактировать ]Выпас приводит к тому, что водоросли поедаются микрозоопланктоном. В результате этого хищничества на дно океана попадает менее 7-10% углерода. Выращивание водорослей на плавучих фермах может позволить этим районам HNLC выращивать водоросли для сбора урожая, не опасаясь нападения хищников. Водоросли, выращенные на плавучих фермах, будут перерабатываться путем выпаса в случае катастрофического отказа плавучей фермы, что ограничит любой ущерб окружающей среде.
Использование
[ редактировать ]Водоросли, выращенные на плавучих фермах, можно собирать и использовать в пищу или топливо. Вся биологическая жизнь состоит из липидов, углеводов, аминокислот и нуклеиновых кислот. Целые водоросли можно превратить в корм для животных, удобрения или биоуголь . Отделение липидов от водорослей также может привести к созданию биодизельного топлива из содержания липидов и биоугля из остального. Конечно, водоросли можно было бы перекачивать на дно океана, ниже любого давления выпаса, для секвестрации.
секвестрация
[ редактировать ]На контролируемой плавучей ферме можно отобрать образцы урожая, чтобы записать количество водорослей на единицу объема, что будет указывать на количество улавливаемого углерода. Если этот углерод будет локализован на дне океана, эту цифру можно будет использовать для точного расчета углеродных кредитов. Связывание углекислого газа на дне океана может разрушить неизученную экосистему и привести к вымиранию неизведанных форм жизни.
Связывание углерода на суше происходит с помощью высушенных водорослей. Без достаточных источников воды бактериям и другим живым существам будет трудно переваривать изолированные водоросли. Биотопливо, которое не продается и не используется в качестве возобновляемого топлива, может храниться в заброшенных нефтяных скважинах и угольных шахтах. Объем биодизельного топлива и масса биоугля дадут точную цифру для производства (при секвестрации) и продажи (при удалении из скважин или шахт) углеродных кредитов.
См. также
[ редактировать ]- Антарктический криль
- Железные удобрения
- Железная гипотеза
- Джон Мартин (океанограф)
- Морской снег
- Первичное производство
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Лалли, СМ; Парсонс, Т.Р. (2004) Биологическая океанография: Введение (2-е изд.) Эльзевир Баттерворт Хайнеманн, Берлингтон, Массачусетс, с. 55.
- ^ Перейти обратно: а б с Питчфорд, JW; Бриндли, Дж. (1999). «Ограничение железа, пастбищное давление и океанические регионы с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC)». Журнал исследований планктона . 21 (3): 525–547. дои : 10.1093/планкт/21.3.525 .
- ^ Перейти обратно: а б Венейблс, Х. и К.М. Мур (2010), Фитопланктон и ограничение света в Южном океане: уроки из районов с высоким содержанием питательных веществ и хлорофилла, J. Geophys. Рез., 115 , C02015, doi:10.1029/2009JC005361
- ^ Перейти обратно: а б Хасслер, CS; Синуар, М.; Клементсон, Луизиана; Батлер, ECV (2012). «Изучение связи между микроэлементами и фитопланктоном в Южном океане австралийским летом 2007 года» . Границы микробиологии . 3 : 202. дои : 10.3389/fmicb.2012.00202 . ПМК 3392650 . ПМИД 22787456 .
- ^ Перейти обратно: а б с Мартин, Джон (1992). Первичная продуктивность и биогеохимические циклы моря . Спрингер США. стр. 122–137.
- ^ Перейти обратно: а б с д и ж г час я дж к Мартин, Джон; Гордон, Майкл; Фицуотер, Стив; Броенков, Уильям В. (1989). «VERTEX: исследования фитопланктона и железа в заливе Аляски». Глубоководные исследования. Часть A: Документы океанографических исследований . 35 (6): 649–680. Бибкод : 1989DSRA...36..649M . дои : 10.1016/0198-0149(89)90144-1 .
- ^ Перейти обратно: а б с д Миллер, Чарльз Б.; Уиллер, Патрисия А. (2012). Биологическая океанография (2-е изд.). Чичестер, Западный Суссекс: John Wiley and Sons, Ltd., стр. 49–62. ISBN 9781444333015 . OCLC 794619582 .
- ^ Редфилд, AC; Кетчум, GH; Ричардс, ФА (1963). «Влияние организмов на состав морской воды». В Хилле, Миннесота (ред.). Море . Нью-Йорк: Wiley-Interscience. стр. 26–77.
- ^ Вернер, Штумм (2013). Водная химия: химическое равновесие и скорость в природных водах . Морган, Джеймс Дж. (3-е изд.). Хобокен: Уайли. ISBN 9780471673033 . OCLC 863203908 .
- ^ Перейти обратно: а б «Влияние железа и цинка на кинетику поглощения кремниевой кислоты и нитратов в трех регионах с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC) (доступна загрузка в формате PDF)» . Исследовательские ворота . Проверено 3 ноября 2017 г.
- ^ Перейти обратно: а б с Андерсон, Массачусетс; Морель, ФММ; Гиллард, RRL (2 ноября 1978 г.). «Ограничение роста прибрежной диатомеи из-за низкой активности ионов цинка». Природа . 276 (5683): 70–71. Бибкод : 1978Natur.276...70A . дои : 10.1038/276070a0 . S2CID 4350372 .
- ^ Перейти обратно: а б с Де Ла Роча, Кристина Л.; Хатчинс, Дэвид А.; Бжезинский, Марк А.; Чжан, Яохун (2000). «Влияние дефицита железа и цинка на элементный состав и производство кремнезема диатомовыми водорослями» (PDF) . Серия «Прогресс в области морской экологии» . 195 : 71–79. Бибкод : 2000MEPS..195...71D . дои : 10.3354/meps195071 . JSTOR 24855011 .
- ^ Дагдейл, Ричард С.; Вилкерсон, Фрэнсис П.; Минас, Ханс Дж. (1 мая 1995 г.). «Роль силикатного насоса в стимулировании нового производства». Глубоководные исследования. Часть I: Статьи океанографических исследований . 42 (5): 697–719. Бибкод : 1995DSRI...42..697D . дои : 10.1016/0967-0637(95)00015-X .
- ^ Перейти обратно: а б с д Дагдейл, Ричард С.; Вилкерсон, Фрэнсис П. (15 января 1998 г.). «Силикатное регулирование новой продукции в экваториальном тихоокеанском апвеллинге». Природа . 391 (6664): 270–273. Бибкод : 1998Natur.391..270D . дои : 10.1038/34630 . S2CID 4394149 .
- ^ Каллен, Джон Дж.; Льюис, Марлон Р.; Дэвис, Кертисс О.; Барбер, Ричард Т. (15 января 1992 г.). «Фотосинтетические характеристики и предполагаемые темпы роста указывают на то, что выпас является непосредственным контролем первичной продукции в экваториальной части Тихого океана» (PDF) . Журнал геофизических исследований: Океаны . 97 (С1): 639–654. Бибкод : 1992JGR....97..639C . дои : 10.1029/91JC01320 . ISSN 2156-2202 .
- ^ Лэндри, MR; Константину, Дж.; Латаса, М.; Браун, СЛ; Бидигаре, Р.Р.; Ондрусек, Мэн (9 августа 2000 г.). «Биологический ответ на внесение железа в восточной экваториальной части Тихого океана (IronEx II). III. Динамика роста фитопланктона и выпаса микрозоопланктона» . Серия «Прогресс в области морской экологии» . 201 : 57–72. Бибкод : 2000MEPS..201...57L . дои : 10.3354/meps201057 . hdl : 10261/184104 .
- ^ стр.15 Оэсте, Франц Дитрих; Рихтер, Рено де; Мин, Тинчжэнь; Кайоль, Сильвен (2017): Климатическая инженерия путем имитации климат-контроля природной пыли. Аэрозольный метод с солями железа. В: Система Земли. Динам. 8 (1), С. 1–54. DOI: 10.5194/esd-8-1-2017
- ^ Перейти обратно: а б с «Уайли: Биологическая океанография, 2-е издание - Чарльз Б. Миллер, Патрисия А. Уиллер» . www.wiley.com . Проверено 11 ноября 2017 г.
- ^ Перейти обратно: а б Бойд, П.В.; Маки, Д.С.; Хантер, Калифорния (2010). «Аэрозольное осаждение железа на поверхность океана. Способы поступления железа и биологические реакции». Морская химия . 120 (1–4): 128–143. Бибкод : 2010Март.120..128B . дои : 10.1016/j.marchem.2009.01.008 .
- ^ Хансард, СП; Лендинг, ВМ; Меры, CI; Фёлкер, Б.М. (2009). «Растворенное железо (II) в Тихом океане: измерения в ходе повторных гидрографических экспедиций PO2 и P16N CLIVAR/CO 2» . Глубоководные исследования. Часть I: Статьи океанографических исследований . 56 (7): 1117–1129. Бибкод : 2009DSRI...56.1117H . дои : 10.1016/j.dsr.2009.03.006 .
- ^ Лам, Фиби Дж.; Епископ, Джеймс КБ (01 апреля 2008 г.). «Окраина континента является ключевым источником железа в северной части Тихого океана HNLC» (PDF) . Письма о геофизических исследованиях . 35 (7): L07608. Бибкод : 2008GeoRL..35.7608L . дои : 10.1029/2008gl033294 . hdl : 1912/3359 . ISSN 1944-8007 .
- ^ Перейти обратно: а б Тиррелл, Т.; Мерико, А.; Ванек, Джей-Джей; Вонг, CS; Мецль, Н.; Уитни, Ф. (1 декабря 2005 г.). «Влияние глубины морского дна на цветение фитопланктона в регионах с высоким содержанием нитратов и низким содержанием хлорофилла (HNLC)» (PDF) . Журнал геофизических исследований: Биогеонауки . 110 (Г2): G02007. Бибкод : 2005JGRG..110.2007T . дои : 10.1029/2005jg000041 . ISSN 2156-2202 . S2CID 129590962 .
- ^ Перейти обратно: а б Лангманн, Б.; Закшек, К.; Хорт, М.; Дагген, С. (2010). «Вулканический пепел как удобрение для поверхности океана» . Химия и физика атмосферы . 10 (8): 3891–3899. Бибкод : 2010ACP....10.3891L . дои : 10.5194/acp-10-3891-2010 . S2CID 55653078 .
- ^ Перейти обратно: а б Пондавен, П.; Руис-Пино, Д.; Дрюон, JN; Фравало, К.; Трегер, П. (1999). «Факторы, контролирующие биогеохимические циклы кремния и азота в системах с высоким содержанием питательных веществ и низким содержанием хлорофилла (Южный океан и северная часть Тихого океана): сравнение с мезотрофной системой (Северная Атлантика)». Глубоководные исследования. Часть I: Статьи океанографических исследований . 46 (11): 1923–1968. Бибкод : 1999DSRI...46.1923P . дои : 10.1016/s0967-0637(99)00033-3 .
- ^ Чавес, Ф. П. и Дж. Р. Тоггвейлер, 1995: Физические оценки новой глобальной продукции: вклад апвеллинга на семинаре в Далеме по апвеллингу в океане: современные процессы и древние записи , Чичестер, Великобритания, John Wiley & Sons, 313-320 .
- ^ Эппли, RW; Ренгер, Э.Х. (15 января 1992 г.). «Утилизация нитратов планктоном в экваториальной части Тихого океана, март 1988 г., вдоль 150 ° з.д.». Журнал геофизических исследований: Океаны . 97 (С1): 663–668. Бибкод : 1992JGR....97..663E . дои : 10.1029/91JC01271 . ISSN 2156-2202 .
- ^ Перейти обратно: а б с д и Винклер, Гизела; Андерсон, Роберт Ф.; Жаккар, Сэмюэл Л.; Маркантонио, Франко (31 мая 2016 г.). «В течение последних 500 000 лет продуктивность экваториальной части Тихого океана контролировала динамика океана, а не пыль» . Труды Национальной академии наук . 113 (22): 6119–6124. Бибкод : 2016PNAS..113.6119W . дои : 10.1073/pnas.1600616113 . ISSN 0027-8424 . ПМЦ 4896667 . ПМИД 27185933 .
- ^ Перейти обратно: а б с Лэндри, Майкл Р.; Селф, Карен Э.; Тейлор, Эндрю Г.; Десима, Мойра; Балч, Уильям М.; Бидигаре, Роберт Р. (1 февраля 2011 г.). «Рост фитопланктона, выпас скота и баланс производства в экваториальной части Тихого океана HNLC». Глубоководные исследования, часть II: Актуальные исследования в океанографии . Динамика планктона и круговорот углерода в экваториальной части Тихого океана. 58 (3): 524–535. Бибкод : 2011DSRII..58..524L . дои : 10.1016/j.dsr2.2010.08.011 .
- ^ Чисхолм, SW, и Морель, FM (1991). Что контролирует производство фитопланктона в богатых питательными веществами районах открытого моря?
- ^ Поллард, Раймонд; Трегер, Поль; Прочтите, Джейн (2006). «Количественная оценка поступления питательных веществ в Южный океан» . Журнал геофизических исследований . 111 (С5): C05011. Бибкод : 2006JGRC..111.5011P . дои : 10.1029/2005JC003076 .
- ^ Моррисон, Адель К.; Фрелихер, Томас Л.; Сармьенто, Хорхе Л. (2015). «Апвеллинг в Южном океане» . Физика сегодня . 68 (1): 27–32. Бибкод : 2015ФТ....68а..27М . дои : 10.1063/PT.3.2654 .
- ^ Санудо-Вильхельми, С., и А.Р. Флегал (2003), Потенциальное влияние пыли Сахары на химический состав Южного океана, Geochem. Геофиз. Геосист., 4, 1063, doi:10.1029/2003GC000507, 7
- ^ де Баар, Хейн Дж.В.; Бойд, Филип В.; Коул, Кеннет Х.; Лэндри, Майкл Р.; Цуда, Ацуши; Эссми, Филипп; Баккер, Дороти CE; Божец, Янн; Барбер, Ричард Т. (1 сентября 2005 г.). «Эксперименты по синтезу железных удобрений: от железного века до эпохи Просвещения» (PDF) . Журнал геофизических исследований: Океаны . 110 (С9): C09S16. Бибкод : 2005JGRC..110.9S16D . дои : 10.1029/2004jc002601 . hdl : 1912/3541 . ISSN 2156-2202 .
- ^ Мартин, Джон Х.; Гордон, Р. Майкл; Фицуотер, Стив Э. (10 мая 1990 г.). «Железо в водах Антарктики». Природа . 345 (6271): 156–158. Бибкод : 1990Natur.345..156M . дои : 10.1038/345156a0 . ISSN 1476-4687 . S2CID 25799856 .
- ^ Поллард, Р., Сандерс, Р., Лукас, М., Стэтхэм, П., 2007. Эксперимент Крозе по цветению и экспорту природного железа (CROZEX). Глубоководные ресурсы. II: Верх. Стад. Океаногр. 54 (18–20), 1905–1914 гг.
- ^ Блен, С., Кегинер, Б., Трулл, Т.В., 2008. Эксперимент KEOPS по естественному удобрению железом (исследование по сравнению океана и плато Кергелен): обзор. Глубоководные ресурсы. II 55, 559–565
- ^ Перейти обратно: а б с Шаретт, Мэтт; Сандерс, Ричард; Чжоу, Мэн (2011). «Природное железное удобрение Южного океана» (PDF) . Глубоководные исследования. Часть II . 90 : 283.
- ^ Венейблс, Хью; Мур, К. Марк (01 февраля 2010 г.). «Фитопланктон и ограничение света в Южном океане: уроки из районов с высоким содержанием питательных веществ и хлорофилла» (PDF) . Журнал геофизических исследований: Океаны . 115 (С2): C02015. Бибкод : 2010JGRC..115.2015V . дои : 10.1029/2009JC005361 . ISSN 2156-2202 .
- ^ Перейти обратно: а б Лиггетт, Даниэла, Брайан Стори, Ивонн Кук и Вероника Медуна, ред. Исследование последнего континента: знакомство с Антарктидой . Спрингер, 2015.
- ^ Перейти обратно: а б Шаретт, Мэтт; Сандерс, Ричард; Чжоу, Мэн (2011). «Природное железное удобрение Южного океана» (PDF) . Глубоководные исследования. Часть II . 90 : 283.
- ^ Перейти обратно: а б с д Мартин, Дж. Х.; Коул, КХ; Джонсон, Канзас; Фицуотер, SE; Гордон, РМ; Таннер, С.Дж.; Хантер, Китай; Элрод, Вирджиния; Новицкий, Дж. Л. (сентябрь 1994 г.). «Проверка гипотезы железа в экосистеме экваториальной части Тихого океана». Природа . 371 (6493): 123–129. Бибкод : 1994Natur.371..123M . дои : 10.1038/371123a0 . hdl : 10945/43402 . S2CID 4369303 .
- ^ Перейти обратно: а б с д и Фуджи, Масахико; Йоши, Наоки; Яманака, Ясухиро; Чай, Фей (2005). «Смоделированные биогеохимические реакции на обогащение железом в трех регионах с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC)». Прогресс в океанографии . 64 (2–4): 307–324. Бибкод : 2005Proce..64..307F . дои : 10.1016/j.pocean.2005.02.017 .
- ^ Перейти обратно: а б с д и Эдвардс, Эндрю М.; Платт, Тревор; Сатьендранатх, Шубха (2004). «Режим океана с высоким содержанием питательных веществ и низким содержанием хлорофилла: ограничения на биомассу и нитраты до и после обогащения железом». Экологическое моделирование . 171 (1–2): 103–125. Бибкод : 2004EcMod.171..103E . doi : 10.1016/j.ecolmodel.2003.06.001 .
- ^ Перейти обратно: а б с д Беренфельд, Майкл Дж. (10 октября 1996 г.). «Подтверждение ограничения фотосинтеза фитопланктона железом в экваториальной части Тихого океана». Природа . 383 (6600): 508–511. Бибкод : 1996Natur.383..508B . дои : 10.1038/383508a0 .
- ^ Мартин, Джон Х.; Фицуотер, Стив Э.; Гордон, Р. Майкл (1 марта 1990 г.). «Дефицит железа ограничивает рост фитопланктона в водах Антарктики». Глобальные биогеохимические циклы . 4 (1): 5–12. Бибкод : 1990GBioC...4....5M . дои : 10.1029/gb004i001p00005 . ISSN 1944-9224 .
- ^ Перейти обратно: а б с «МГЭИК – Межправительственная группа экспертов по изменению климата» . www.ipcc.ch. Проверено 3 ноября 2017 г.
- ^ Лам, Фиби Дж.; Тортелл, Филипп Д.; Морель, Франсуа М.М. (март 2001 г.). «Дифференциальное влияние добавок железа на производство органического и неорганического углерода фитопланктоном». Лимнология и океанография . 46 (5): 1199–1202. Бибкод : 2001LimOc..46.1199L . дои : 10.4319/lo.2001.46.5.1199 . S2CID 94868090 .
- ^ Перейти обратно: а б с д Трик, Чарльз Г.; Билл, Брайан Д.; Кочлан, Уильям П.; Уэллс, Марк Л.; Тренер Вера Л.; Пикелл, Лиза Д. (30 марта 2010 г.). «Обогащение железом стимулирует производство токсичных диатомей в районах с высоким содержанием нитратов и низким содержанием хлорофилла» . Труды Национальной академии наук . 107 (13): 5887–5892. Бибкод : 2010PNAS..107.5887T . дои : 10.1073/pnas.0910579107 . ISSN 0027-8424 . ПМЦ 2851856 . ПМИД 20231473 .
- ^ Перейти обратно: а б с Цуда, Ацуши; Такеда; Сайто; Нисиока (июль 2007 г.). «Доказательства гипотезы выпаса: выпас снижает реакцию фитопланктона экосистемы HNLC на обогащение железом в западной субарктической части Тихого океана (SEEDS II)». Журнал океанографии . 63 (6): 983–994. Бибкод : 2007JOce...63..983T . дои : 10.1007/s10872-007-0082-x . S2CID 56097872 .
- ^ Перейти обратно: а б Миони, Сесиль Э.; Хэнди, Сара М.; Эллвуд, Майкл Дж.; Твисс, Майкл Р.; Маккей, Р. Майкл Л.; Бойд, Филип В.; Вильгельм, Стивен В. (1 декабря 2005 г.). «Отслеживание изменений биодоступного железа в океанических водах с высоким содержанием нитратов и низким содержанием хлорофилла: первая оценка с использованием гетеротрофного бактериального биорепортера» . Глобальные биогеохимические циклы . 19 (4): GB4S25. Бибкод : 2005GBioC..19.4S25M . дои : 10.1029/2005gb002476 . hdl : 1885/28454 . ISSN 1944-9224 . S2CID 14401749 .
- ^ Перейти обратно: а б Питчфорд, Джонатан Уильям; Бриндли, Джон (1999). «Ограничение железа, пастбищное давление и океанические регионы с высоким содержанием питательных веществ и низким содержанием хлорофилла (HNLC)». Журнал исследований планктона . 21 (3): 525–547. дои : 10.1093/планкт/21.3.525 .
- ^ Джикеллс, Т.Д.; Ан, ЗС; Андерсен, К.К.; Бейкер, Арканзас; Бергаметти, Дж.; Брукс, Н.; Цао, Джей-Джей; Бойд, П.В.; Дуче, РА (1 апреля 2005 г.). «Глобальные железные связи между пылью пустыни, биогеохимией океана и климатом». Наука . 308 (5718): 67–71. Бибкод : 2005Sci...308...67J . CiteSeerX 10.1.1.686.1063 . дои : 10.1126/science.1105959 . ISSN 0036-8075 . ПМИД 15802595 . S2CID 16985005 .
- ^ Беренфельд, Майкл Дж.; Кольбер, Збигнев С. (5 февраля 1999 г.). «Широко распространенное ограничение фитопланктона железом в южной части Тихого океана». Наука . 283 (5403): 840–843. Бибкод : 1999Sci...283..840B . дои : 10.1126/science.283.5403.840 . ПМИД 9933166 .
- ^ Вагенер, Тибо; Гье, Сесиль; Лосно, Реми; Бонне, Софи; Маховальд, Натали (1 июня 2008 г.). «Возвращение к экспорту атмосферной пыли в океан Южного полушария: биогеохимические последствия» (PDF) . Глобальные биогеохимические циклы . 22 (2): GB2006. Бибкод : 2008GBioC..22.2006W . дои : 10.1029/2007gb002984 . ISSN 1944-9224 . S2CID 55663098 .
- ^ Уолш, Джон Дж. (1 января 1976 г.). «Травоядность как фактор использования питательных веществ в море1» . Лимнология и океанография . 21 (1): 1–13. Бибкод : 1976LimOc..21....1W . дои : 10.4319/lo.1976.21.1.0001 . ISSN 1939-5590 .
- ^ Перейти обратно: а б с д Фрост, Брюс В. (1991). «Роль выпаса скота в богатых питательными веществами районах открытого моря» . Лимнология и океанография . 38 (8): 1616–1630. Бибкод : 1991LimOc..36.1616F . дои : 10.4319/lo.1991.36.8.1616 .
- ^ Минас, Ханс Иоахим; Минас, Моник (1992). «Чистая продукция сообщества в водах тропического и Антарктического океанов с высоким содержанием питательных веществ и низким содержанием хлорофилла: гипотеза выпаса против железа». Океанологика Акта . 15 (2): 145–162.
- ^ Фрост, BW (27 июля 1987 г.). «Контроль выпаса запасов фитопланктона в открытой субарктической части Тихого океана: модель, оценивающая роль мезозоопланктона, особенно крупных каланоидных копепод Neocalanus spp» . Серия «Прогресс в области морской экологии» . 39 : 49–68. Бибкод : 1987MEPS...39...49F . дои : 10.3354/meps039049 .
- ^ Беренфельд, Майкл Дж.; Бэйл, Энтони Дж.; Кольбер, Збигнев С.; Эйкен, Джеймс; Фальковски, Пол Г. (октябрь 1996 г.). «Подтверждение ограничения фотосинтеза фитопланктона железом в экваториальной части Тихого океана». Природа . 383 (6600): 508–511. Бибкод : 1996Natur.383..508B . дои : 10.1038/383508a0 .
- ^ Перейти обратно: а б с Зибе, RE; Арчер, Д. (1 мая 2005 г.). «Возможность удобрения океана и его влияние на будущие уровни CO2 в атмосфере». Письма о геофизических исследованиях . 32 (9): L09703. Бибкод : 2005GeoRL..32.9703Z . дои : 10.1029/2005gl022449 . ISSN 1944-8007 .
- ^ Перейти обратно: а б с д и «Вклад Рабочей группы III в Четвертый оценочный отчет Межправительственной группы экспертов по изменению климата, 2007 г.» . www.ipcc.ch. Проверено 3 ноября 2017 г.
- ^ Уоллер, Риан (18 октября 2012 г.). «Железные удобрения: решение проблемы изменения климата или сброс отходов в океан?» . Нэшнл Географик . Проверено 20 октября 2017 г.
- ^ Бойд, Филип В.; Закон, Клифф С.; Вонг, CS; Нодзири, Юкихиро; Цуда, Ацуши; Левассер, Морис; Такеда, Сигэнобу; Ривкин, Ричард; Харрисон, Пол Дж. (17 марта 2004 г.). «Упадок и судьба вызванного железом цветения субарктического фитопланктона». Природа . 428 (6982): 549–553. Бибкод : 2004Natur.428..549B . дои : 10.1038/nature02437 . ISSN 1476-4687 . ПМИД 15058302 . S2CID 4384093 .
- ^ Цао, Лонг; Калдейра, Кен (01 марта 2010 г.). «Может ли удобрение океана железом смягчить закисление океана?». Климатические изменения . 99 (1–2): 303–311. Бибкод : 2010ClCh...99..303C . дои : 10.1007/s10584-010-9799-4 . ISSN 0165-0009 . S2CID 153613458 .
