Островная карликовость

Островная карликовость , форма филетической карликовости . [ 1 ] это процесс и состояние крупных животных, эволюционирующих или имеющих уменьшенный размер тела [ а ] когда ареал их популяции ограничен небольшой средой обитания, в первую очередь островами. Этот естественный процесс отличается от преднамеренного создания карликовых пород, называемого карликовостью . Этот процесс происходил много раз на протяжении всей эволюционной истории, например, различные виды карликовых слонов , которые развились в эпоху плейстоцена , а также более древние примеры, такие как динозавры европазавр и мадьярозавр . Этот процесс и другие артефакты « островной генетики » могут происходить не только на островах, но и в других ситуациях, когда экосистема изолирована от внешних ресурсов и размножения. Это могут быть пещеры , пустынные оазисы , изолированные долины и изолированные горы (« небесные острова »). [ нужна ссылка ] Островная карликовость является одним из аспектов более общего «островного эффекта» или «правила Фостера» , которое утверждает, что, когда материковые животные колонизируют острова, мелкие виды имеют тенденцию развивать более крупные тела ( островной гигантизм ), а крупные виды имеют тенденцию развивать более мелкие тела. Это сам по себе один из аспектов островного синдрома , который описывает различия в морфологии , экологии , физиологии и поведении островных видов по сравнению с их континентальными собратьями.
Возможные причины
[ редактировать ]Есть несколько предложенных объяснений механизма, вызывающего такую карликовость. [ 3 ] [ 4 ]
Один из них представляет собой селективный процесс, при котором выживают только более мелкие животные, оказавшиеся на острове, поскольку количество пищи периодически снижается до пограничного уровня. Меньшим животным нужно меньше ресурсов и меньшие территории, и поэтому они с большей вероятностью преодолеют переломный момент, когда сокращение популяции позволит пополнить источники пищи, достаточные для процветания выживших. Меньший размер также выгоден с репродуктивной точки зрения, поскольку он влечет за собой более короткие периоды беременности и время генерации . [ 3 ]
В тропиках небольшой размер должен облегчить терморегуляцию . [ 3 ]
Среди травоядных крупный размер дает преимущества в борьбе как с конкурентами, так и с хищниками, поэтому сокращение или отсутствие того и другого будет способствовать карликовости; конкуренция представляется более важным фактором. [ 4 ]
Считается, что среди хищников основным фактором является размер и доступность ресурсов добычи, а конкуренция считается менее важной. [ 4 ] У тигровых змей островная карликовость возникает на островах, где доступная добыча ограничена меньшими размерами, чем обычно добывают материковые змеи. Поскольку предпочтения змей по размеру добычи обычно пропорциональны размеру тела, маленькие змеи могут лучше адаптироваться к ловле мелкой добычи. [ 5 ]
Карликовость против гигантизма
[ редактировать ]Обратный процесс, при котором мелкие животные, размножающиеся на изолированных островах, лишенных хищников с больших территорий, могут стать намного крупнее обычных, называется островным гигантизмом . Отличным примером является дронт , предками которого были голуби нормального размера . Есть также несколько видов гигантских крыс , одна из которых сохранилась до сих пор, которые сосуществовали как с Homo floresiensis , так и с карликовыми стегодонтами на Флоресе.
Процесс островной карликовости может происходить относительно быстро по эволюционным меркам. Это контрастирует с увеличением максимального размера тела, которое происходит гораздо более постепенно. При нормализации к длине поколения было обнаружено, что максимальная скорость уменьшения массы тела во время островной карликовости более чем в 30 раз превышает максимальную скорость увеличения массы тела при десятикратном изменении у млекопитающих. [ 6 ] Считается, что это неравенство отражает тот факт, что педоморфизм предлагает относительно простой путь к уменьшению размера тела взрослого человека; с другой стороны, эволюция более крупных максимальных размеров тела, вероятно, будет прервана появлением ряда ограничений, которые необходимо преодолеть с помощью эволюционных инноваций, прежде чем процесс сможет продолжиться. [ 6 ]
Факторы, влияющие на степень карликовости
[ редактировать ]Как для травоядных, так и для хищников размер острова, степень изоляции острова и размер предковых континентальных видов, по-видимому, не имеют прямого значения для степени карликовости. [ 4 ] Однако, если принять во внимание только массы тела современных высших травоядных и плотоядных животных и включить данные как по континентальным, так и по островным участкам суши, было обнаружено, что массы тела крупнейших видов на суше масштабируются до размера суши, при этом уклоны около 0,5 бревна (масса тела/кг) на бревно (площадь земли/км) 2 ). [ 7 ] Существовали отдельные линии регрессии для эндотермических высших хищников, экзотермических высших хищников, эндотермических высших травоядных и (на основании ограниченных данных) экзотермических высших травоядных, так что потребление пищи у высших травоядных было в 7-24 раза выше, чем у высших хищников. и примерно одинаково для эндотерм и эктотермов одного трофического уровня (это приводит к тому, что эктотермы в 5–16 раз тяжелее соответствующих эндотермы). [ 7 ]
Было высказано предположение, что для карликовых слонов конкуренция была важным фактором размера тела: на островах, где есть конкурирующие травоядные, карликовые слоны были значительно крупнее, чем на тех, где конкурирующие травоядные животные отсутствовали. [ 8 ]
Примеры
[ редактировать ]Нептичьи динозавры
[ редактировать ]Признание того, что островная карликовость может распространяться и на динозавров, возникло благодаря работе Ференца Нопчи , аристократа венгерского происхождения, искателя приключений, ученого и палеонтолога. Нопча интенсивно изучал трансильванских динозавров, заметив, что они меньше своих собратьев в других частях света. Например, он обнаружил шестиметровых зауроподов — группу динозавров, длина которых в других местах обычно вырастала до 30 метров и более. Нопча пришел к выводу, что местом, где были найдены останки, был остров Хацег (ныне бассейн Хацег или Хацег в Румынии ) в мезозойскую эпоху. [ 9 ] [ 10 ] Предложение Нопчи о карликовости динозавров на острове Хацег сегодня широко принято после того, как дальнейшие исследования подтвердили, что найденные останки принадлежат не молодым особям. [ 11 ]
Зауроподы
[ редактировать ]| Пример | Разновидность | Диапазон | Временные рамки | Континентальный родственник |
|---|---|---|---|---|
Ампелозавр |
А. атакис | Иберо — Арморикан остров | Поздний мел / Маастрихт |  Немегтозавриды |
 Европазавр |
Э. Хольгери | Нижняя Саксония | Поздняя юра /средний кимеридж |  Брахиозавры |
 Магьярозавр |
М. дакус | Остров Хатег | Поздний мел / Маастрихт |  Рапетозавр |
 Лирайнозавр [ 12 ] |
Л. астибиае | Иберо-Армориканский остров | Поздний мел | |
 Палудититан |
П. налатценсис | Остров Хатег | Поздний мел / Маастрихт |  Эпахтозавр |
Другой
[ редактировать ]| Пример | Разновидность | Диапазон | Временные рамки | Континентальный родственник |
|---|---|---|---|---|
Лангенбергский карьер торвозавр (синий) |
Безымянный | Нижняя Саксония | Поздняя юра /средний кимеридж | Торвозавр |
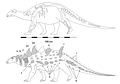 Струтиозавр [ 13 ] |
Святой австрийский С. трансильваникус С. languedocensis |
Иберо-Армориканские, Австрало-Альпийские острова и острова Хатег. | Поздний мел |  Эдмонтония |
 Телматозавр |
Т. транссильваникус | Остров Хатег | Поздний мел | Гадрозавриды |
 Текодонтозавр [ 10 ] |
Т. древний | Южная Англия | Поздний триас / ретий |  Платеозавры |
 Залмокс [ 10 ] (фиолетовый) |
З. робуста г-н Шкиперорум |
Остров Хатег | Поздний мел |  Тенонтозавр |
Кроме того, род Balaur первоначально был описан как велоцираптора размером с дромеозаврид (и, как следствие, как сомнительный пример островной карликовости), но с тех пор был реклассифицирован как вторично нелетающая стволовая птица, более близкая к современным птицам, чем Jeholornis (таким образом, на самом деле это пример островного гигантизма ).
Птицы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
 Гавайские нелетающие ибисы |
Аптерибис гленос | Молокаи | Вымершие (поздний четвертичный период ) |  Американские ибисы |
|
| Вы найдете это коротким | Мауи | ||||
| Косумель-курассо [ 14 ] | Кракс рубра грискоми | Косумель | Неизвестный |  Великий курасов |
|
 Эму с острова Кенгуру [ 15 ] |
Dromaius novaehollandiae baudinianus | Остров Кенгуру , Южная Австралия | Вымершие (ок. 1827 г. н.э.) |  Emu |
|
 Эму с острова Кинг [ 16 ] (черный) |
Dromaius novaehollandiae минор | Остров Кинг , Тасмания | Вымершие (1822 г. н. э.) | ЛР ≈ 0,48 [ б ] | |
| Карликовый желтоглазый пингвин [ 17 ] | Megadyptes antipodes richdalei | Острова Чатем , Новая Зеландия | Вымершие (после 1300 г. н. э.) |  Желтоглазый пингвин |
|
 Косумельский трэшер [ 14 ] |
Токсостома глутатум | Косумель | Находится под угрозой исчезновения |  Другие трэшеры |
Весы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
 Мадагаскарский карликовый хамелеон |
Брукезия минимум | Остров Нуси-Бе , Мадагаскар | Находящийся под угрозой исчезновения |  Мадагаскарские листовые хамелеоны | |
 Нуси Хара хамелеон [ 18 ] |
Брукезия микра | Остров Нуси Хара , Мадагаскар | Уязвимый | ||
| Тигровая змея острова Роксби [ 5 ] | Обратите внимание на щиты | Остров Роксби, Южная Австралия | Неизвестный |  Тигровая змея |
|
| Карликовый бирманский питон | Программирование Python | Ява , Бали , Сумбава и Сулавеси , Индонезия | Неизвестный |  Бирманский питон |
ЛР ≈ 0,44 [ с ] |
| Сетчатый питон Танахджампеа [ 21 ] | Python reticulatus jampeanus | Танахджампеа , между Сулавеси и Флоресом. | Неизвестный |  Сетчатый питон |
LR ≈ 0,41, самцы LR ≈ 0,49, самки [ д ] |
Млекопитающие
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
 Карликовый трехпалый ленивец |
Брадипус пигмей | Остров Верагуас Шилд , Панама | Находится под угрозой исчезновения |  Бурогорлый ленивец |
 Акратокнус |
А. антилленсис А. одонтригонус A. ye |
Куба , Эспаньола и Пуэрто-Рико | Вымершие (ок. 3000 г. до н. э.) |  Континентальные наземные ленивцы |
| Имагокнус | И. зазае | Куба | Вымершие ( ранний миоцен ) | |
 Мегалокн |
М. Роденс М. дней |
Куба и Эспаньола | Вымершие (ок. 2700 г. до н. э.) | |
 Неокнус |
Neocnus виды. | Куба и Эспаньола | Вымершие (ок. 3000 г. до н. э.) |
Хоботки
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
| Сулавесский карликовый слон | Знаменитый слон | Сулавеси | Вымершие (ранний плейстоцен) |  Азиатский слон |
 Кабарруйский карликовый слон |
Элефас Бейери | Лусон | Вымерший | |
 Критский карликовый мамонт |
Критский мамонт | Крит | Вымерший |  Маммут |
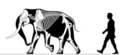 Мамонт с Нормандских островов |
Изгнанный мамонт | Санта-Роза Остров | Вымершие ( поздний плейстоцен ) | Колумбийский мамонт |
| Сардинский мамонт | Маммут ламарморай | Сардиния | Вымершие (поздний плейстоцен) |  Степной мамонт |
| Шерстистый мамонт с острова Сент-Пол [ 24 ] [ 25 ] | Оригинальный мамонт | Остров Сент-Пол , Аляска | Вымершие (ок. 3750 г. до н. э.) | Шерстистый мамонт |
 Сикуло-мальтийские слоны |
Палеолоксодон древний Леонарди П. мнайдриенсис П. мелитенсис П. фальконери |
Сицилия и Мальта | Вымерший |  Слон с прямым бивнем (левый) |
| Критские слоны | Палеолоксодон chaniensis П. Крейцбурги |
Крит | Вымерший | |
 Кипрский карликовый слон |
Палеолоксодон киприотский | Кипр | Вымершие (ок. 9000 г. до н.э.) | |
| Наксосский карликовый слон | Палеолоксодон сп. | Наксос | Вымерший | |
| Карликовый слон Родоса и Тилоса | Палеолоксодон тиленсис | Родос и Тилос | Вымерший | |
| Бумиаю карликовый синомастодонт [ 26 ] | Синомастодон бумиаюенсис | Остров Бумиаю (ныне часть Явы ) | Вымершие (ранний плейстоцен) |  Синомастодон |
 Японский стегодонт [ 27 ] [ 28 ] |
Стегодон миенсис Стегодон протоавроры Стегодон рассвета |
Япония (также Тайвань для S. aurorae ) [ 29 ] | Вымершие ( ранний плейстоцен ) |  Китайский стегодон |
| Карликовый стегодонт Большого Флореса [ 3 ] | Стегодон флоренсис | Флорес | Вымершие (поздний плейстоцен) |  Сундаленд Стегодон |
| Яванские карликовые стегодонты | Стегодон гипсилофус [ 26 ] С. semedoensis [ 30 ] С. сп. [ 26 ] |
Ява | Вымершие ( четвертичный период ) | |
| Карликовый стегодонт Минданао [ 31 ] | Стегодон минданский | Минданао и Сулавеси | Вымершие (средний плейстоцен) | |
| Сулавесский карликовый стегодонт [ 26 ] | Стегодон сомпоенсис | Сулавеси | Вымерший | |
| Карликовый стегодонт Малого Флореса [ 3 ] | Стегодон грешник | Флорес | Вымершие (средний плейстоцен) | |
| Карликовый стегодонт Сумба [ 32 ] | Стегодон сумбаенсис | Сумба , Индонезия | Вымершие (средний плейстоцен) | |
| Тиморский карликовый стегодонт [ 26 ] | Стегодон тиморенсис | Тимор | Вымерший | |
| Карликовый стеголофодонт [ 33 ] | Стеголофодон псевдолатиденс | Япония | Вымершие ( миоцен ) |  Стеголофодон |
Приматы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
| Карликовый лемур Нуси Хара [ 34 ] | Хейрогалеус сп. | Остров Нуси Хара у Мадагаскара | Неизвестный |  Карликовые лемуры |
 Цветы Мужчина [ 35 ] |
Человек флоресский | Флорес | Вымершие (поздний плейстоцен) |  Мужчина встал |
 Кальяо Мэн |
Человек лузонский [ 36 ] [ 37 ] | Лусон , Филиппины | Вымершие (поздний плейстоцен) | |
| Современные пигмеи Флореса [ 38 ] | Мудрый человек | Флорес | Сохранившийся | другие представители Homo Sapiens |
| Ранние современные люди Палау (оспариваются) [ 39 ] | Мудрый человек | дворец | Вымерший (?) | |
| Андаманский [ 40 ] | Мудрый человек | Андаманские острова | Сохранившийся | |
 Сардинская макака [ 41 ] |
Большие макаки | Сардиния | Вымершие (плейстоцен) |  Берберийская макака |
 Занзибарский красный колобус |
Пилиоколобус кирки | Унгуджа | Находящийся под угрозой исчезновения |  Красный колобус Удзунгва |
Хищники
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
 Сицилийский волк |
Хрустальная волчья собака | Сицилия | Вымершие (1970 г. н.э.) |  Серый волк |
|
 Японский волк |
Canis lupus hodophilax | Япония (кроме Хоккайдо ) | Вымершие (1905 г. н.э.) | ||
 Сардинский волк (вперед) |
Цинотерий сардозный | Корсика и Сардиния | Вымершие (ок. 8300 г. до н. э.) | 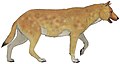 Ксеноцион |
|
| Тринилская собака | Мецецион триниленсис | Ява | Вымершие (плейстоцен) | ||
| Пальто с острова Косумель [ 14 ] | Насуа Нарика Нельсони | Косумель | Находится под угрозой исчезновения |  Юкатанская белоносая коати |
|
 Занзибарский леопард |
Пантера пардус пардус | Унгуджа | Находящиеся под угрозой исчезновения или вымершие |  Африканский леопард |
|
 Балийский тигр |
Пантера тигр сондайка | Бали | Вымершие (ок. 1940 г. н. э.) |  Суматранский тигр | |
 Яванский тигр |
Ява | Вымершие (ок. 1975 г. н. э.) | |||
 Косумельский енот |
Процион Пигмей | Косумель | Находится под угрозой исчезновения |  Обыкновенный енот |
|
 Островная лиса |
Уроцион прибрежный | Шесть Нормандских островов Калифорнии | Рядом с угрозой |  Серая лиса |
ЛР ≈ 0,84 [ и ] ЛР ≈ 0,75 [ ж ] |
| Косумельская лиса | Уроцион сп. | Косумель | Находящиеся под угрозой исчезновения или вымершие |
Нежующие копытные
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
 Эумайохерус |
Эумайохоэр этрусский | Бачинелло , Монтебамболи | Вымершие ( миоцен ) |  Микростоникс |
 Малагасийские карликовые бегемоты |
Бегемот лалумена Х. Лемерлей H. madagascariensis |
Мадагаскар | Вымершие (ок. 1000 г. н. э.) |  Обыкновенный бегемот |
| Карликовый бегемот Бумиаю [ 26 ] | Гексапротодон симплекс | Остров Бумиаю (ныне Ява ) | Вымершие ( ранний плейстоцен ) |  Азиатские бегемоты |
 Критский карликовый бегемот |
Бегемот Крейцбурги | Крит | Вымершие ( средний плейстоцен ) |  Европейский бегемот |
 Мальтийский карликовый бегемот |
Бегемот Мелитенсис | Мальта | Вымершие (плейстоцен) | |
 Кипрский карликовый бегемот |
Бегемот минор | Кипр | Вымершие (ок. 8000 г. до н.э.) | |
 Сицилийский карликовый бегемот |
Бегемот пентланди | Сицилия | Вымершие (плейстоцен) | |
| Косумельский пекари с воротником [ 14 ] | Пекари таджаку карлик | Косумель | Неизвестный |  Пекари с воротником |
| Филиппинский носорог [ 44 ] | Незоринус филиппинский | Лусон | Вымершие (средний плейстоцен) |  Яванский носорог |
Бовиды
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
| Сицилийский бизон [ 27 ] | Древний сицилийский бизон | Сицилия | Вымершие (поздний плейстоцен) |  Степной зубр |
| Сицилийские зубры [ 45 ] | Оригинальный сицилийский скот [ 27 ] | Сицилия | Вымершие (поздний плейстоцен) |  Евразийские зубры |
| Себу Тамарав | Бубалус cebuensis | Себу , Филиппины | Вымерший |  Дикий водяной буйвол |
 Значение низменности |
Буффало депрессикорнис | Сулавеси и Бутон , Индонезия | Находящийся под угрозой исчезновения | |
| Бубалус гровеси | Бубалус гровеси | Сулавеси , Индонезия | Вымерший | |
 Тамарав |
Бубалус миндорский | Миндоро , Филиппины | Находится под угрозой исчезновения | |
 Снова гора |
Буйвол из Куарлези | Сулавеси и Бутон , Индонезия | Находящийся под угрозой исчезновения | |
 Пещерный козел Балеарских островов |
Миотрагус балеарикус | Майорка и Менорка | Вымершие (после 3000 г. до н.э.) | Галлогорал |
| Несогорал [ 46 ] | Несогорал виды. | Сардиния | Вымерший | |
| Газель Дахлак Кебир [ 47 ] | Нангер сёммерринги ssp. | Остров Дахлак Кебир , Эритрея | Уязвимый |  Газель Земмерринга |
Тиренотрагус |
Самый тонкий тирренотрагус | Бачинелло | Вымерший | Antilopinae sp. |
Олени и родственники
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
 Критский олень [ г ] |
Candiacervus виды. | Крит | Вымершие (плейстоцен) | Неизвестный |
 Сардинский олень [ 10 ] |
Praemegaceros cazioti | Сардиния | Вымершие (ок. 5500 г. до н. э.) | Премегасерос |
 Карликовый олень Рюкю [ 50 ] |
Цервус астилодон | Острова Рюкю | Вымерший |  Пятнистый олень (?) Дояпонский олень (?) |
| Популяция благородных оленей в Джерси [ 51 ] | Джерси олень | Джерси | Вымершие (плейстоцен) |  Красный олень |
 Корсиканский благородный олень |
Корсиканский элафский олень | Корсика и Сардиния | Рядом с угрозой | |
| Сицилийский благородный олень [ 27 ] | Олень Сицилии | Сицилия | Вымершие (поздний плейстоцен) | |
 Гоплитомерикс [ ч ] |
Hoplitomeryx виды. | Остров Гаргано | Вымершие ( ранний плиоцен ) |  Скотоводство |
| Сицилийская лань | Дама карбурангеленсис | Сицилия | Вымершие (поздний плейстоцен) | Лань |
 Флоридский ключевой олень |
Odocoileus Virginianus ключей | Флорида-Кис | Находящийся под угрозой исчезновения |  Вирджиния олень |
 Шпицбергенский олень |
Рангифер тарандус платиринхус | Шпицберген | Уязвимый |  Северный олень |
 Филиппинский олень |
Русская Мариана | Филиппины | Уязвимый |  Самбарский олень |
| Возможный пример | Биномиальное имя | Родной диапазон | Статус | Континентальный родственник |
|---|---|---|---|---|
 Островные слоновьи кактусы [ 52 ] [ 53 ] |
Пахицереус принглей | Отдаленные острова в море Кортеса (например, Санта-Крус , Сан-Педро-Мартир ) |
Не оценено |  Материковые слоновьи кактусы |
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Примером неостровной филетической карликовости является эволюция карликовых игрунок и тамаринов среди обезьян Нового Света, кульминацией которой стало появление самого маленького экземпляра, Cebuella pygmaea . [ 2 ]
- ^ Судя по высотам на рис. 1 Heupink et al. , 2011 г. [ 16 ]
- ^ На основе максимальной длины 2,5 м для карликовой формы. [ 19 ] и 5,74 м для материковой формы. [ 20 ]
- ^ На основе максимальной общей длины питона Танахджампеа (TL) 2,10 м для самцов и 3,35 м для самок. [ 21 ] и максимальная длина от морды южного суматранского питона до жерла (SVL) 4,5 м для самцов и 6,1 м для самок. [ 22 ] с SVL, скорректированными до TL путем умножения на коэффициент 1,127, полученный на основе средней относительной длины хвоста (0,113) африканских и индийских каменных питонов. [ 23 ]
- ^ Для близлежащих материковых серых лисиц. [ 42 ]
- ^ Для материковых серых лисиц в целом. [ 43 ]
- ^ Как и Hoplitomeryx , Candiacervus представляет собой необычный случай, поскольку представители этого рода эволюционировали в островные виды широкого диапазона размеров, причем не только карликовые формы, но и некоторые, которые можно считать гигантами. [ 48 ] [ 49 ]
- ^ Hoplitomeryx , очевидно, представляет собой довольно необычный случай, поскольку представители этого рода, по-видимому, эволюционировали как в карликовые, так и в гигантские островные формы на одном и том же острове(ах). [ 48 ]
Ссылки
[ редактировать ]- ^ Протеро, Дональд Росс ; Серено, Пол Каллистус (зима 1982 г.). «Аллометрия и палеоэкология карликовых носорогов среднего миоцена с прибрежной равнины Техасского залива» . Палеобиология . 8 (1): 16–30. Бибкод : 1982Pbio....8...16P . дои : 10.1017/S0094837300004322 . JSTOR 2400564 . S2CID 88464305 .
- ^ Перельман П.; и др. (2011). «Молекулярная филогения живых приматов» . ПЛОС Генетика . 7 (3): 1–17. дои : 10.1371/journal.pgen.1001342 . ПМК 3060065 . ПМИД 21436896 .
- ^ Jump up to: а б с д и Ван Ден Берг, Геррит Дирк; Слава, Рохус Дуэ; Морвуд, Майкл Джон ; Сутикна, Томас; Джатмико; Вахью Саптомо, Э. (май 2008 г.). «Самый молодой стегодон остается в Юго-Восточной Азии из археологических раскопок позднего плейстоцена Лян Буа, Флорес, Индонезия». Четвертичный интернационал . 182 (1): 16–48. Бибкод : 2008QuInt.182...16V . дои : 10.1016/j.quaint.2007.02.001 .
- ^ Jump up to: а б с д Райя, Паскуале; Мейри, Шай (август 2006 г.). «Островное правило у крупных млекопитающих: палеонтология встречается с экологией». Эволюция . 60 (8): 1731–1742. дои : 10.1111/j.0014-3820.2006.tb00516.x . ПМИД 17017072 . S2CID 26853128 .
- ^ Jump up to: а б Кио, Дж.С.; Скотт, IAW; Хейс, К. (январь 2005 г.). «Быстрое и неоднократное возникновение островного гигантизма и карликовости у австралийских тигровых змей» . Эволюция . 59 (1): 226–233. дои : 10.1111/j.0014-3820.2005.tb00909.x . ПМИД 15792242 . S2CID 58524 .
- ^ Jump up to: а б Эванс, Арканзас; и др. (30 января 2012 г.). «Максимальная скорость эволюции млекопитающих» . ПНАС . 109 (11): 4187–4190. Бибкод : 2012PNAS..109.4187E . дои : 10.1073/pnas.1120774109 . ПМК 3306709 . ПМИД 22308461 .
- ^ Jump up to: а б Бернесс, врач общей практики; Даймонд, Дж .; Фланнери, Т. (4 декабря 2001 г.). «Динозавры, драконы и карлики: эволюция максимального размера тела» . Труды Национальной академии наук . 98 (25): 14518–14523. Бибкод : 2001PNAS...9814518B . дои : 10.1073/pnas.251548698 . ISSN 0027-8424 . JSTOR 3057309 . ПМК 64714 . ПМИД 11724953 .
- ^ ван дер Гир, Александра А.Е.; ван ден Берг, Геррит Д.; Лирас, Джордж А.; Прасетио, Унггул В.; Должное, Рокус Трепет; Сетиябуди, Эрик; Дриния, Хара (август 2016 г.). «Влияние площади и изоляции на островные карликовые хоботки» . Журнал биогеографии . 43 (8): 1656–1666. дои : 10.1111/jbi.12743 . ISSN 0305-0270 .
- ^ «Остров карликовых динозавров действительно существовал, утверждают ученые» . Телеграф Медиа Группа . 22 февраля 2010 г. Архивировано из оригинала 25 февраля 2010 г. Проверено 26 февраля 2010 г.
- ^ Jump up to: а б с д Бентон, MJ; Чики, З.; Григореску, Д.; Редельсторфф, Р.; Сандер, премьер-министр; Штейн, К.; Вейшампель, Д.Б. (28 января 2010 г.). «Динозавры и власть острова: карликовые динозавры с острова Хацег» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 293 (3–4): 438–454. Бибкод : 2010PPP...293..438B . дои : 10.1016/j.palaeo.2010.01.026 . Архивировано из оригинала (PDF) 10 июля 2011 г. Проверено 30 июля 2017 г.
- ^ Дайк, Г. (20 сентября 2011 г.). «Динозавр-барон Трансильвании». Научный американец . 305 (4): 80–83. Бибкод : 2011SciAm.305c..80D . doi : 10.1038/scientificamerican1011-80 . ПМИД 22106812 .
- ^ Компания, Дж. (2010). «Гистология костей титанозавра Lirainosaurus astibiae (Dinosauria: Sauropoda) из позднего мела Испании». Naturwissenschaften . 98 (1): 67–78. дои : 10.1007/s00114-010-0742-3 . hdl : 10251/148874 . ПМИД 21120450 . S2CID 31752413 .
{{cite journal}}:|last1=имеет общее имя ( справка ) - ^ Карпентер, К. (2001) Бронированные динозавры. Издательство Университета Индианы, 526 страниц.
- ^ Jump up to: а б с д Куарон, AD; Мартинес-Моралес, Массачусетс; Макфадден, Квебек; Валенсуэла, Д.; Гомппер, Мэн (2004). «Состояние карликовых хищников на острове Косумель, Мексика». Биоразнообразие и сохранение . 13 (2): 317–331. Бибкод : 2004BiCon..13..317C . CiteSeerX 10.1.1.511.2040 . doi : 10.1023/b:bioc.0000006501.80472.cc . S2CID 25730672 .
- ^ Паркер С. (1984) Вымерший эму с острова Кенгуру, до сих пор непризнанный вид. Бюллетень Клуба британских орнитологов 104: 19–22.
- ^ Jump up to: а б Хойпинк, TH; Хайнен, Л.; Ламберт, DM (2011). «Древняя ДНК предполагает, что карликовые и «гигантские» эму относятся к одному виду» . ПЛОС ОДИН . 6 (4): e18728. Бибкод : 2011PLoSO...618728H . дои : 10.1371/journal.pone.0018728 . ПМК 3073985 . ПМИД 21494561 .
- ^ Коул, Тереза Л. и др. «Митогеномы открывают вымершие таксоны пингвинов и показывают, что образование островов является ключевым фактором видообразования». Молекулярная биология и эволюция 36.4 (2019): 784-797.
- ^ Гло, Ф.; Келер, Дж.; Таунсенд, ТМ; Венсес, М. (14 февраля 2012 г.). «Соперничество с самыми маленькими в мире рептилиями: открытие миниатюрных и микроэндемичных новых видов листовых хамелеонов ( брукезия ) из Северного Мадагаскара» . ПЛОС ОДИН . 7 (2): e31314. Бибкод : 2012PLoSO...731314G . дои : 10.1371/journal.pone.0031314 . ПМЦ 3279364 . ПМИД 22348069 .
- ^ де Ланг Р., Фогель Г. (2005). Змеи Сулавеси: Полевой справочник по наземным змеям Сулавеси с идентификационными ключами . Франкфуртский вклад в группу 25 по естественной истории, издание Chimaira, 2005 г. ISBN 3-930612-85-2 . стр. 23–27, 198–201.
- ^ Баркер, Д.Г.; Бартен, СЛ; Эрсам, JP; Даддоно, Л. (2012). «Исправленная длина двух известных гигантских питонов и установление нового рекорда максимальной длины для бирманских питонов, Python bivittatus » (PDF) . Бюллетень Чикагского герпетологического общества . 47 (1): 1–6 . Проверено 02 марта 2020 г.
- ^ Jump up to: а б Аулия, М.; Маусфельд, П.; Шмитц, А.; Бёме, В. (9 апреля 2002 г.). «Обзор сетчатого питона ( Python reticulatus Schneider, 1801) с описанием нового подвида из Индонезии». Naturwissenschaften . 89 (5): 201–213. Бибкод : 2002NW.....89..201A . дои : 10.1007/s00114-002-0320-4 . ПМИД 12135085 . S2CID 4368895 .
- ^ Шайн, Р.; Харлоу, PS; Кио, Дж.С.; Боади, Н.И. (1998). «Влияние пола и размера тела на пищевые привычки гигантской тропической змеи Python reticulatus » . Функциональная экология . 12 (2): 248–258. Бибкод : 1998FuEco..12..248S . дои : 10.1046/j.1365-2435.1998.00179.x .
- ^ Шихи, CM; Альберт, Дж.С.; Лиллиуайт, HB; Ван Дамм, Р. (2016). «Эволюция длины хвоста у змей, связанная с различными гравитационными средами» . Функциональная экология . 30 (2): 244–254. Бибкод : 2016FuEco..30..244S . дои : 10.1111/1365-2435.12472 . ; см. Таблицу S1
- ^ Ширбер, Майкл. Пережить вымирание: где выжили шерстистые мамонты . Живая наука. Корпорация Имагинова. Проверено 20 июля 2007 г.
- ^ Мамонты острова Врангеля , к северу от Сибири , больше не считаются карликами. См.: Тихонов Алексей; Ларри Агенброуд; Сергей Вартанян (2003). Сравнительный анализ популяций мамонтов на острове Врангеля и Нормандских островах . ДЕИНСЕА 9: 415–420. ISSN 0923-9308
- ^ Jump up to: а б с д и ж Азиз, Ф.; ван ден Берг, Грузия (25 сентября 1995 г.). «Карликовый стегодон из Самбунгмакана (Центральная Ява, Индонезия)» . Учеб. Кон. Нед. акад. В. Ветенш . 98 (3): 229–241 . Проверено 31 июля 2017 г.
- ^ Jump up to: а б с д Сондаар, штат Пенсильвания; ААЭ ван дер Гир (2005). «Эволюция и вымирание островных копытных плио-плейстоцена» . Международный журнал Французской четвертичной ассоциации . 2 : 241–256 . Проверено 31 июля 2017 г.
- ^ Айба, Хироаки; Баба, Кацуёси; Мацукава, Масаки (10 марта 2010 г.). «Новый вид стегодона (Mammalia, Proboscidea) из группы Казуса (нижний плейстоцен), город Хатиодзи, Токио, Япония и его эволюционная морфодинамика: STEGODON PROTOAURORAE SP. NOV. И МОРФОДИНАМИКА» . Палеонтология . 53 (3): 471–490. дои : 10.1111/j.1475-4983.2010.00953.x . S2CID 128161878 .
- ^ ван ден Берг, Герт Д.; де Вос, Джон; Сондаар, Пол Ю. (25 сентября 2000 г.). «Позднечетвертичная палеогеография эволюции млекопитающих на Индонезийском архипелаге» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 171 (3–4): 385–408. дои : 10.1016/S0031-0182(01)00255-3 .
- ^ Сисванто, С., и Ноэрвиди, С. (2014). ИСКОПАЕМЫЕ ХОБОСКИДЕИ С ПЛОЩАДКИ СЕМЕДО: ее корреляция с биостратиграфией и прибытием человека на Яву. Беркала Археология, 34 (2).
- ^ Заим, Ю. (20 августа 2010 г.). «Геологические свидетельства самого раннего появления гомининов в Индонезии» . Во Флигле, Дж. Г.; Ши, Джей-Джей; Грин, FE; Баден, Алабама; Лики, Р.Э. (ред.). Из Африки I: Первая колонизация Евразии гомининами . Springer Science & Business Media. п. 106. ИСБН 978-90-481-9036-2 . OCLC 668096676 .
- ^ Сетиябуди, Эрик; Курниаван, Иван; Ван Ден Берг, Геррит. «Окаменелости Stegodon и Varanus komodoensis Sumba и Flores: сухопутный мост плейстоцена» . Факультет науки, медицины и здравоохранения.
- ^ Саэгуса, Х. (2008). «Карликовый стеголофодон из миоцена Японии: пассажиры на тонущих лодках». Четвертичный интернационал . 182 (1): 49–62. Бибкод : 2008QuInt.182...49S . дои : 10.1016/j.quaint.2007.08.001 .
- ^ «Новая группа карликовых лемуров может оказаться редчайшим приматом в мире »
- ^ Ученый, изучающий морфинг хоббитов , abc.net.au
- ^ Уэйд, Л. (10 апреля 2019 г.). «Новый вид древнего человека обнаружен на Филиппинах». Наука . 364 . doi : 10.1126/science.aax6501 . S2CID 189045520 .
- ^ Детройт, Ф.; Михарес, А.С.; Корни, Дж.; Дэйвер, Г.; Занолли, К.; Дизон, Э.; Роблес, Э.; Грюн, Р.; Пайпер, Пи Джей (2019). «Новый вид Homo из позднего плейстоцена Филиппин» (PDF) . Природа . 568 (7751): 181–186. Бибкод : 2019Natur.568..181D . дои : 10.1038/s41586-019-1067-9 . ПМИД 30971845 . S2CID 106411053 .
- ^ Туччи, С.; и др. (03.08.2018). «Эволюционная история и адаптация человеческой популяции пигмеев на острове Флорес, Индонезия» . Наука . 361 (6401): 511–516. Бибкод : 2018Sci...361..511T . дои : 10.1126/science.aar8486 . ПМК 6709593 . ПМИД 30072539 .
- ^ « Древние маленькие люди на Палау, а не карлики, говорится в исследовании » . Национальные географические новости . 27 августа 2008 г.
- ^ Мондаль, М.; Казальс, Ф.; Сюй, Т.; Далл'Олио, генеральный менеджер; Пибус, М.; Нетеа, Миннесота; Комас, Д.; Лааюни, Х.; Ли, К.; Маджумдер, ПП; Бертранпети, Дж. (2016). «Геномный анализ андаманцев дает представление о миграции древних людей в Азию и их адаптации» (PDF) . Природная генетика . 48 (9): 1066–1070. дои : 10.1038/ng.3621 . hdl : 10230/34401 . ПМИД 27455350 . S2CID 205352099 .
- ^ Рук, Л. (31 декабря 2008 г.). «Первый семинар по летописям европейских ископаемых приматов (Сиена и Гроссето, 11–13 сентября 2008 г.) с обновленной информацией об итальянских исследованиях в области палеоприматологии» (PDF) . Атти Мусс. Стор. Нат. Маремма (22): 129–143.
- ^ Парфит, М.; Гру, М. (22 апреля 2020 г.). «Воодушевляющая история об этих крошечных островных лисицах, почти уничтоженных катастрофой» . NationalGeographic.com . Нэшнл Географик . Архивировано из оригинала 17 апреля 2020 года . Проверено 23 апреля 2020 г.
- ^ Мур, CM; Коллинз, PW (1995). «Виды млекопитающих - Urocyonlittoralis » (PDF) . 489 : 1–7. Архивировано из оригинала (PDF) 22 января 2012 года . Проверено 16 сентября 2011 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Ренема, Виллем (2007). Биогеография, время и место: распространение, барьеры и острова . Springer Science & Business Media. п. 334. ИСБН 978-1-4020-6374-9 . OCLC 228153573 .
- ^ ван Вюре, СНГ (2005). По следам зубра: история, морфология и экология вымершего дикого быка . Коронет Букс Инкорпорейтед. ISBN 978-954-642-235-4 . OCLC 472741798 .
- ^ ван дер Гир, А.; Лирас, Г; де Вос, Дж.; Дермицакис, М. (14 февраля 2011 г.). «Сардиния и Корсика» . Эволюция островных млекопитающих: адаптация и вымирание плацентарных млекопитающих на островах . Джон Уайли и сыновья. ISBN 978-1-4443-9128-2 . OCLC 894698082 .
- ^ Чиоцци, Г.; Барделли, Г.; Риччи, М.; Де Марки, Г.; Кардини, А. (2014). «Просто еще один островной карлик? Фенотипические особенности малоизвестной газели Земмерринга, Nanger soemmerringii (Cetartiodactyla: Bovidae), с острова Дахлак-Кебир» . Биологический журнал Линнеевского общества . 111 (3): 603–620. дои : 10.1111/bij.12239 .
- ^ Jump up to: а б Мацца, PPA; Росси, Массачусетс; Агостини, С. (2015). «Hoplitomerycidae (поздний миоцен, Италия), пример гигантизма у островных жвачных животных». Журнал эволюции млекопитающих . 22 (2): 271–277. дои : 10.1007/s10914-014-9277-2 . S2CID 16437411 .
- ^ ван дер Гир, AAE (2018). «Однородность в разнообразии: морфология и эволюция рогов в среде, свободной от хищников» . Электронная палеонтология (21.1.9A): 1–31. дои : 10.26879/834 .
- ^ Кайфу, Ю.; Фудзита, М.; Йонеда, М.; Ямасаки, С. (15 февраля 2015 г.). «Плейстоценовое мореплавание и колонизация островов Рюкю, юго-запад Японии» . В Кайфу, Ю.; Идзухо, М.; Гебель, Т.; Сато, Х.; Оно, А. (ред.). Возникновение и разнообразие поведения современного человека в палеолитической Азии . Издательство Техасского университета A&M. ISBN 978-1-62349-277-9 . OCLC 985023261 .
- ^ Листер, AM (30 ноября 1989 г.). «Быстрое уменьшение численности благородных оленей на Джерси в последнее межледниковье». Природа . 342 (6249): 539–542. Бибкод : 1989Natur.342..539L . дои : 10.1038/342539a0 . ПМИД 2685610 . S2CID 4343091 .
- ^ Уайлдер, Британская Колумбия; Фельгер, Р.С. (30 сентября 2010 г.). «Карликовые гиганты, гуано и изоляция: растительность и флористическое разнообразие острова Сан-Педро-Мартир, Калифорнийский залив, Мексика» (PDF) . Труды Общества естественной истории Сан-Диего . 42 : 1–24, см. стр. 9–13 . Проверено 05 января 2020 г.
(стр. 12) Карликовость растений Сан-Педро-Мартира, по-видимому, связана с отбором более коротких особей, чтобы пережить жестокие тропические штормы, возможной конкуренцией корней в таком густом лесу и неопределенным эффектом высоких уровней азота и фосфора из обилие гуано, которое может замедлить рост. Генетические исследования не проводились...
- ^ Бернс, КК (май 2019 г.). Эволюция в изоляции: поиск островного синдрома у растений . Издательство Кембриджского университета . стр. 174–177. дои : 10.1017/9781108379953 . ISBN 978-1108379953 . OCLC 1108160200 . S2CID 186536407 .
(стр. 174-175) ... степень, в которой его карликовый рост генетически детерминирован, и объяснение того, почему островная карликовость может быть выборочно выгодной, ждут дополнительных исследований.
Внешние ссылки
[ редактировать ]| Правила |
|  |
|---|---|---|
| Связанный |
| |