Термостабильная ДНК-полимераза
Термостабильные ДНК-полимеразы — это ДНК-полимеразы , которые происходят от термофилов , обычно бактерий или архей, и поэтому являются термостабильными . Они используются для полимеразной цепной реакции и связанных с ней методов амплификации и модификации ДНК .
Характеристики
[ редактировать ]Было описано несколько ДНК-полимераз с различными свойствами, которые определяют их специфическое использование в ПЦР, ПЦР в реальном времени или при изотермической амплификации . Будучи ДНК-полимеразами, все термостабильные ДНК-полимеразы обладают 5'→3'-полимеразной активностью, а также экзонуклеазной активностью либо 5'→3', либо 3'→5'.
| Полимераза | Так | Тфл | Тт | Фрагмент Бст Кленова (BF), смещение пряди | Тли (Вент) | P GB-D (глубокая вентиляция) | Pfx (КОД) | Пфу |
|---|---|---|---|---|---|---|---|---|
| Организм | Термус водный | Термус желтый | Термус термофилус | Геобацилла стеаротермофилус | Термококк литоралис | Пирококк сп. штамм ГБ-Д | Пирококк кодакаренсис | Пирококк безумный |
| Источник | бактериальный | бактериальный | бактериальный | бактериальный | архейный | архейный | архейный | архейный |
| Молекулярный вес | 80 кДа [1] | 94 кДа [1] | 94 кДа [1] | 67 кДа [2] | 90 кДа [1] | 90 кДа [3] | 90 кДа [3] | 92кДа [1] |
| Температура расширения | 74 °С [1] | 74 °С [1] | 74 °С [1] | 65 °С [2] | 74 °С [1] | 75 °С [3] | 75 °С [3] | 75 °С [1] |
| 5'→3' экзонуклеазная активность | Да [1] | Да [1] | Да [1] | Нет [2] | Нет [1] | Нет [1] | ||
| 3'→5'-экзонуклеазная активность | Нет [1] | Нет [1] | Нет [1] | Нет [2] | Да [1] | Да [3] | Да [3] | Да [1] |
| Активность обратной транскриптазы | Слабый [1] | Да [1] | Да [1] | Слабый [2] | Нет [1] | Н/Д [1] | ||
| ПЦР заканчивается | 3'-А [1] | 3'-А [1] | 3'-А [1] | 3'-А [2] | 70% Блант; 30% Однобазовый [1] | Тупой [3] | Тупой [3] | Тупой [1] |
| Верность (ошибки на базу и удвоение) | 8 × 10 −6 [4] 1.5 × 10 −4 [5] 3-5.6 × 10 −5 [6] | 1.5 × 10 −5 [7] | 2.8 × 10 −6 [4] | 2.7 × 10 −6 [4] 4.0 × 10 −6 [5] | 3.5 × 10 −6 [8] 1.2 × 10 −5 [5] 7.6 × 10 −6 [6] | 1.3 × 10 −6 [9] [4] 5.1 × 10 −6 [5] 2.8 × 10 −6 [6] | ||
| Скорость синтеза (баз/сек.) | 21–47 [2] 61 [10] | 191 [2] | 23 [10] | 120 [11] 106–138 [10] | 9.3–25 [10] | |||
| Процессивность (базы) | 10–42 [10] | <20 [10] | >300 [10] | 6.4–20 [10] |
Структура
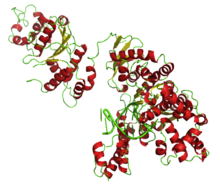
[ редактировать ]ДНК-полимеразы имеют форму руки с большим пальцем, ладонью и остальными пальцами. [12] [13] Большой палец участвует в связывании и перемещении двухцепочечной ДНК . [12] Ладонь несет активный центр полимеразы , тогда как пальцы связывают субстраты (матрицу ДНК и нуклеозидтрифосфаты ). [12] [14] Экзонуклеазная активность находится в отдельном белковом домене . [12] мг 2+ является кофактором .
Активный центр полимеразы на ладони катализирует удлинение ДНК, начиная с праймера, связанного с одной цепью матричной ДНК:
- дезоксинуклеозидтрифосфат + ДНК n ⇌ пирофосфат + ДНК n+1 .
Бактериальные полимеразы
[ редактировать ]Термостабильные ДНК-полимеразы природного происхождения обнаружены у термофильных бактерий , архей и их возбудителей. Среди бактериальных термостабильных ДНК-полимераз полимераза Taq , полимераза Tfl, полимераза Tma, полимераза Tne, полимераза Tth и Bst. используются [4] [15] [16] [2]
В дополнение к 5'→3'-полимеразной активности бактериальные термостабильные ДНК-полимеразы (принадлежащие к ДНК-полимеразам А-типа) обладают 5'→3'-экзонуклеазной активностью и генерируют аденозиновый выступ ( липкие концы ) на 3'-конце. вновь образованная прядь. Фрагмент Кленова Bst (BF) обладает активностью замещения цепи, что позволяет использовать его в изотермической амплификации без необходимости денатурации ДНК в термоциклере , а его экзонуклеазную активность 5'→3' удаляют для более высокого выхода. [2]
Архейные полимеразы
[ редактировать ]
Часто используемые ДНК-полимеразы B-типа — это полимераза Pfu , [4] Pwo-полимераза, [17] полимераза КОД, [3] полимераза Tli (также называемая Vent), происходящая из различных архей, [18] тег-полимераза, [19] Tce-полимераза, [20] полимераза Тго, [8] полимераза TNA1, [21] полимераза Tpe, [22] полимераза Tthi, [23] Neq-полимераза [24] и Pab-полимераза. [25]
Архейные варианты (относящиеся к В-типу) дают тупые концы (полимераза Tli дает выступ примерно в 30% продуктов) и вместо 5'→3' экзонуклеазной активности обладают активностью по исправлению ошибок синтеза (доказательство- чтение), активность экзонуклеазы 3'→5'. [26] [27] В архейных полимеразах частота ошибок снижается при создании аналога фрагмента Кленова, поскольку при этом корректирующая экзонуклеазная активность удаляется. [4] Некоторые ДНК-полимеразы архей характеризуются не столько пригодностью для стандартной ПЦР, сколько сниженным ингибированием амплификации А-ДНК. [28] или ДНК с модифицированными основаниями. [29] [30]
Модифицированные полимеразы
[ редактировать ]Различные слитые белки с низкой частотой ошибок архей и высокой скоростью синтеза бактериальных термостабильных ДНК-полимераз ( полимеразы Q5 ) были получены из различных термостабильных полимераз и ДНК-зажима термостабильного ДНК-связывающего белка SSo7d путем белкового дизайна . [31] Слитый белок гомолога PCNA из Archaeoglobus fulgidus также был получен с помощью термостабильных ДНК-полимераз архей. [32] слитые белки термостабильных ДНК-полимераз с термостабильным ДНК-связывающим белковым доменом топоизомеразы ( тип V, с мотивом спираль-шпилька-спираль , HhH) из Methanopyrus kandleri Аналогично были созданы ( TopoTaq и PfuC2 ). [33] [34] Модифицированная полимераза Pfu также была создана с помощью дизайна белка ( Pfu Ultra ). [35] Аналогичные эффекты достигаются и смесями термостабильных ДНК-полимераз обоих типов при соотношении в смеси ферментативных активностей полимераз типа А и В 30 к 1. [22] [36] например Геркулаза [8] и ТакПлюс [10] как коммерческая смесь полимераз Taq и Pfu, Развернуть как коммерческую смесь Taq и Pwo, [37] Расширьте High Fidelity как коммерческую смесь Taq и Tgo. [10] Platinum Taq High Fidelity как коммерческая смесь Taq и Tli (Vent), [10] и Advantage HF 2 в виде коммерческой смеси Titanium Taq и неназванной корректорской полимеразы. [10] Эти смеси можно использовать для ПЦР на большие расстояния для синтеза продуктов длиной до 35 т.п.н. [36] [38] Другие добавки используются, чтобы помочь против сложных последовательностей, богатых G C , избежать или нейтрализовать негативные эффекты ингибиторов ПЦР (например, компонентов крови или детергентов). [39] или дУТП [40] ), или изменить кинетику реакции . [41]
Скорость и процессивность
[ редактировать ]Сравнены базовые скорости синтеза (скорость, продуктивность) различных полимераз. [8] Скорость синтеза Taq-полимеразы составляет около 60 пар оснований в секунду. Среди немодифицированных термостабильных ДНК-полимераз только скорость синтеза КОД-полимеразы превышает 100 пар оснований в секунду (около 120 п.о./с). [11] Среди модифицированных термостабильных ДНК-полимераз описаны различные мутации, увеличивающие скорость синтеза. [42] [43] [44] КОД-полимераза и некоторые модифицированные термостабильные ДНК-полимеразы ( iProof / Phusion , Pfu Ultra , Velocity или Z-Taq ) используются как вариант ПЦР с более короткими циклами амплификации (быстрая ПЦР, высокоскоростная ПЦР) из-за их высокой скорости синтеза. Процессивность описывает среднее количество пар оснований до того, как полимераза оторвется от матрицы ДНК. Процессивность полимеразы ограничивает максимальное расстояние между праймером и зондом в некоторых формах количественной ПЦР в реальном времени (кПЦР).
Верность
[ редактировать ]Описаны коэффициенты ошибок различных полимераз (верность). Частота ошибок Taq-полимеразы составляет 8 × 10. −6 ошибок на базу, у Advantage HF 6,1 × 10 −6 ошибок на базу, у Platinum Taq High Fidelity 5,8 × 10 −6 ошибки на базу и удвоение, как у TaqPlus 4 × 10 −6 ошибки на основание и удвоение, у КОД-полимеразы 3,5 × 10 −6 ошибки на основание и удвоение, у Tli-полимеразы и Геркулазы 2,8 × 10 −6 ошибки на базу и удвоение, у Deep Vent 2,8 × 10 −6 ошибки на основание и удвоение, таковые для Pfu, ДНК-полимеразы Phusion (идентичной ДНК-полимеразе iProof ) и Herculase II Fusion 1,3 × 10 −6 ошибки на основание и удвоение, а также у Pfu Ultra и Pfu Ultra II 4,3 × 10 −7 ошибки на базу и удвоение. [4] [8] [10] Более новый анализ выявил несколько иную частоту ошибок: Deep Vent (экзо-) полимераза (5,0 × 10 −4 ошибки на основание и удвоение), Taq-полимераза (1,5 × 10 −4 ошибок на базу и удвоение), Kapa HiFi HotStart ReadyMix (1,6 × 10 −5 ошибки на базу и удвоение), КОД (1,2 × 10 −5 ошибок на базу и удвоение), PrimeSTAR GXL (8,4 × 10 −6 ошибок на основание и удвоение), Pfu (5,1 × 10 −6 ошибки на основание и удвоение), ДНК-полимераза Deep Vent (4,0 × 10 −6 ) ошибки на основание и удвоение, Phusion (3,9 × 10 −6 ошибки на основание и удвоение) и ДНК-полимераза Q5 (5,3 × 10 −7 ошибки на базу и удвоение). [5] Еще один обнаружил частоту ошибок 3–5,6 × 10. −6 для Така, 7,6 × 10 −6 для КОДА, 2,8 × 10 −6 для Пфу, 2,6×10 −6 для Phusion и 2,4×10 −6 для Пво. [6] Чтобы уменьшить количество мутаций в продукте ПЦР (например, при молекулярном клонировании ), в ПЦР можно использовать больше матричной ДНК и меньше циклов. [10]
Урожай
[ редактировать ]Бактериальные термостабильные ДНК-полимеразы обычно производят более высокие концентрации продуктов, чем архейные, но с большим количеством ошибок копирования. В бактериальных термостабильных ДНК-полимеразах фрагмент Кленова ( Klen-Taq ) или фрагмент Стоффеля может быть получен путем удаления экзонуклеазного домена в ходе дизайна белка, аналогично ДНК-полимеразе из E. coli , что приводит к более высокому продукту. концентрация. [45] [15] Две аминокислоты, необходимые для экзонуклеазной функции Taq-полимеразы, были идентифицированы с помощью мутагенеза как аргинины в положениях 25 и 74 (R25 и R74). [46] Мутация гистидина в положении 147 (сокращенно : на глутаминовую кислоту H147E) в полимеразе KOD снижает относительно высокую экзонуклеазную активность KOD. [27]
Нуклеотидная специфичность
[ редактировать ]Предпочтение отдельных нуклеотидов термостабильной ДНК-полимеразой называется нуклеотидной специфичностью (смещенностью). на основе ПЦР При секвенировании ДНК с субстратами обрыва цепи (дидезокси-метод) часто желательно их равномерное включение и, следовательно, беспристрастное получение всех продуктов обрыва цепи, чтобы обеспечить более высокую чувствительность и упростить анализ. Для этой цели KlenTaq была создана полимераза путем делеции, а фенилаланин в положении 667 был заменен на тирозин посредством сайт-направленного мутагенеза (сокращенно: F667Y) и назван термосеквеназой . [47] [48] Эту полимеразу также можно использовать для включения дидезоксинуклеотидов, меченных флуоресценцией. [49]
Термостабильные ДНК-полимеразы с горячим стартом
[ редактировать ]Матричная специфичность полимераз повышается за счет использования полимераз с горячим стартом, чтобы избежать связывания праймеров с нежелательными матрицами ДНК или друг с другом при низких температурах перед началом ПЦР. [50] Примерами являются ингибируемая антителами полимераза Pfu Pfu Turbo , Platinum Pfx в виде коммерческой полимеразы KOD с ингибирующим антителом и Platinum Taq в качестве полимеразы Taq, ингибируемой антителами. [8] Полимеразы горячего старта ингибируются инактивацией формальдегидом . [51] [52] (или малеиновый ангидрид , экзо-цис-3,6-эндоксо-Δ4-тетрагидрофталевый ангидрид, цитраконовый ангидрид, 3,4,5,6-тетрагидрофталевый ангидрид, цис-аконитовый ангидрид или 2,3-диметилмалеиновый ангидрид), [53] путем комплексообразования магния с фосфатами [54] или путем связывания антитела с их активным сайтом. [55] [56] При нагревании до 95°С формальдегид диссоциирует от белков. [57] [58] [59] или ионы магния высвобождаются, [54] или антитело денатурируется и высвобождается в процессе. [60] [61] Кроме того, полимеразы можно ингибировать аптамерами , которые денатурируют при нагревании. [62] [63] Пятый вариант — это полимераза, адсорбированная на латексных шариках за счет гидрофобного эффекта и растворяющаяся при повышении температуры. В шестом, самом старом варианте, реакционную смесь без полимеразы покрывают воском и добавляют полимеразу поверх охлажденного воска. При нагревании восковой слой плавится и полимераза смешивается с реакционной смесью. [64]
Другие ДНК-полимеразы
[ редактировать ]Некоторые ДНК-полимеразы, используемые при изотермической амплификации ДНК, например, при петлевой изотермической амплификации , множественной амплификации смещения , амплификации рекомбиназной полимеразы или изотермической сборке , для амплификации целых геномов (например, ДНК-полимераза φ29 из бактериофага phi29 , B35DNAP из фага Bam35 ) не термостабильны, в то время как другие, такие как фрагмент Бст Кленова, термостабильны. [65] ДНК-полимеразы Т4, Т6 и Т7 также не являются термостабильными.
РНК-зависимые ДНК-полимеразы
[ редактировать ]Стандартные обратные транскриптазы (РНК-зависимые ДНК-полимеразы) ретровирусного происхождения, используемые для ОТ-ПЦР , такие как AMV и MoMuLV обратная транскриптаза , не являются термостабильными при 95 °C. При более низких температурах обратной транскрипции может произойти неспецифическая гибридизация праймеров вторичных с неправильными последовательностями, а также образование нежелательных структур в матрице ДНК, что может привести к образованию нежелательных продуктов ПЦР и менее желательных продуктов ПЦР. Обратная транскриптаза AMV может использоваться при температуре до 70 °C. [66] Кроме того, некоторые термостабильные ДНК-зависимые ДНК-полимеразы можно использовать в качестве РНК-зависимых ДНК-полимераз путем замены Mg 2+ в качестве кофакторов с Mn 2+ , чтобы их можно было использовать для RT-PCR. [67] Но поскольку скорость синтеза Taq с Mn 2+ относительно невелик, для этого подхода все чаще использовался Tth. [68] Использование Мн 2+ также увеличивается частота ошибок и необходимое количество шаблонов, поэтому этот метод используется редко. Этих проблем можно избежать с помощью термостабильной 3173 -полимеразы термофильного бактериофага , которая выдерживает высокие температуры ПЦР и предпочитает РНК в качестве матрицы. [69]
Приложения
[ редактировать ]Помимо выбора термостабильной ДНК-полимеразы, в ходе оптимизации ПЦР специально изменяются и другие параметры ПЦР.
Помимо ПЦР, термостабильные ДНК-полимеразы также используются для вариантов ОТ-ПЦР , кПЦР в различных вариантах, сайт-специфического мутагенеза и секвенирования ДНК. Их также используют для производства гибридизационных зондов для Саузерн-блоттинга и Нозерн-блоттинга путем случайного прайминга. Активность экзонуклеазы 5'→3' используется для трансляции ников и TaqMan , среди прочего, без репликации (амплификации) ДНК.
История
[ редактировать ]Элис Чиен и ее коллеги первыми охарактеризовали термостабильную полимеразу Taq в 1976 году. [70] Впервые термостабильную ДНК-полимеразу использовали Рэндалл К. Сайки и его коллеги в 1988 году, представив Taq-полимеразу для ПЦР. [71] [72] Термостабильность Taq-полимеразы устраняет необходимость добавления нетермостабильной ДНК-полимеразы в реакцию после каждой фазы плавления ПЦР, поскольку полимераза Taq не денатурируется при нагревании до 95 ° C во время фазы плавления каждого цикла. полимеразы Taq В 1989 году ген был клонирован, и полимераза Taq была получена в Escherichia coli в виде рекомбинантного белка . [73] [72] ДНК длиной до 35 000 пар оснований была синтезирована Уэйном М. Барнсом с использованием различных смесей полимераз типа A и B. [36] [72] тем самым создавая ПЦР дальнего действия. Высокая скорость синтеза полимеразы KOD была опубликована в 1997 году Масахиро Такаги и его коллегами. [3] [72] [14] тем самым создавая основы высокоскоростной ПЦР. В последующие годы были разработаны и другие оптимизации ПЦР, например, обход ингибиторов ПЦР и амплификация сложных последовательностей ДНК, богатых GC, [41] а также модификация термостабильных ДНК-полимераз путем создания белков. разработали изотермическую амплификацию, опосредованную петлей В 1998 году Цугунори Нотоми и его коллеги из Eiken Chemical Company , с использованием полимеразы Bst при 65 ° C. [74] [75]
Дальнейшее чтение
[ редактировать ]- Дж. Сэмбрук , Т. Маниатис, Д. У. Рассел: Молекулярное клонирование: лабораторное руководство . Колд-Спринг-Харбор Лабораторный пресс ; 3-е издание (2001 г.), ISBN 0-87969-577-3.
- Ф. Акрам, Ф. И. Шах, Р. Ибрар, Т. Фатима, И. Ю. Хак, В. Насим, М. А. Гул, Л. Техрим, Г. Хайдер: Бактериальные термофильные ДНК-полимеразы: акцент на выдающиеся биотехнологические применения. В кн.: Аналитическая биохимия. Том 671, июнь 2023 г., с. 115150, два : 10.1016/j.ab.2023.115150 , ПМИД 37054862 .
- К. Терпе: Обзор термостабильных ДНК-полимераз для классических приложений ПЦР: от молекулярных и биохимических основ до коммерческих систем. В кн.: Прикладная микробиология и биотехнология . Том 97, выпуск 24, декабрь 2013 г., с. 10243–10254, два : 10.1007/s00253-013-5290-2 , ПМИД 24177730 .
Внешние ссылки
[ редактировать ]- НЭБ Полбаза . По состоянию на 27 сентября 2012 г.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление Промега: Свойства термостабильных ДНК-полимераз (PDF; 208 КБ). По состоянию на 27 сентября 2012 г.
- ^ Перейти обратно: а б с д и ж г час я дж И. Оскорбин, М. Филипенко: Bst-полимераза – скромная родственница Taq-полимеразы. В: Журнал вычислительной и структурной биотехнологии. Том 21, 2023 г., с. 4519–4535, два : 10.1016/j.csbj.2023.09.008 , ПМИД 37767105 , ПМК 1052051 .
- ^ Перейти обратно: а б с д и ж г час я дж М. Такаги, М. Нисиока, Х. Какихара, М. Китабаяши, Х. Иноуэ, Б. Каваками, М. Ока, Т. Иманака: Характеристика ДНК-полимеразы Pyrococcus sp. штамм KOD1 и его применение в ПЦР. В: Прикл. Окружающая среда. Микробиол. , Том 63, Выпуск 11, 1997, с. 4504–4510. ПМИД 9361436 ; ПМК 168769 .
- ^ Перейти обратно: а б с д и ж г час Дж. Клайн, Дж. К. Браман, Х. Х. Хогрефе: Точность ПЦР ДНК-полимеразы БОЕ и других термостабильных ДНК-полимераз. В: Нуклеиновые кислоты Res . , Том 24, Выпуск 18, 1996 г., с. 3546–3551. ПМИД 8836181 ; ПМК 146123 .
- ^ Перейти обратно: а б с д и Потапов В, Онг Дж.Л. (2017). «Исследование источников ошибок в ПЦР путем секвенирования одиночных молекул» . ПЛОС ОДИН . 12 (1): e0169774. Бибкод : 2017PLoSO..1269774P . дои : 10.1371/journal.pone.0169774 . ПМК 5218489 . ПМИД 28060945 .
- ^ Перейти обратно: а б с д П. МакИнерни, П. Адамс, М.З. Хади: Сравнение частоты ошибок во время полимеразной цепной реакции с помощью ДНК-полимеразы. В: Международная молекулярная биология. 2014, с. 287430, дои : 10.1155/2014/287430 , ПМИД 25197572 , ПМК 4150459 .
- ^ Хун Го Фань, Чжай Фэн, Хуан Вэй-Хуа: EP0810288 (A2) - 3 декабря 1997 г.: Новая ДНК-полимераза с корректурной 3'-5' экзонуклеазной активностью.
- ^ Перейти обратно: а б с д и ж Бахрам Арези, Вэймэй Син, Джозеф А. Зорге, Холли Х. Хогрефе (15 октября 2003 г.), «Эффективность амплификации термостабильных ДНК-полимераз» (PDF) , Аналитическая биохимия , том. 321, нет. 2, стр. 226–235, номер документа : 10.1016/S0003-2697(03)00465-2 , PMID 14511688.
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Х.Х. Хогрефе, М. Борнс, Высокоточные ферменты ПЦР, в: К.В. Диффенбах, Г.С. Двекслер (ред.), PCR Primer: Лабораторное руководство, Лабораторное издательство Колд-Спринг-Харбор, Колд-Спринг-Харбор, Нью-Йорк, 2003.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Agilent: Ферменты для высокоточной ПЦР: свойства и определение частоты ошибок.
- ^ Перейти обратно: а б Европейский патент 1752534A1: Высокоскоростная ПЦР с использованием высокоскоростной ДНК-полимеразы , 12 мая 2005 г. / 14 февраля 2007 г., Тойо Босеки (заявитель) и Масая Сегава и др. (изобретатели).
- ^ Перейти обратно: а б с д Т. А. Стейтц: ДНК-полимеразы: структурное разнообразие и общие механизмы. В: Журнал биологической химии . Том 274, выпуск 25, июнь 1999 г., с. 17395–17398, дои : 10.1074/jbc.274.25.17395 , ПМИД 10364165 .
- ^ П. Дж. Ротвелл, Г. Ваксман: Структура и механизм ДНК-полимераз. В: Достижения в химии белков. Том 71, 2005 г., с. 401–440, два : 10.1016/S0065-3233(04)71011-6 , ПМИД 16230118 .
- ^ Перейти обратно: а б Х. Хасимото, М. Нисиока, П. Фудзивара, М. Такаги, Т. Иманака, Т. Иноуэ, Ю. Кай: Кристаллическая структура ДНК-полимеразы гипертермофильных архей Pyrococcus kodakaraensis KOD1. В: Журнал молекулярной биологии. Том 306, выпуск 3, февраль 2001 г., с. 469–477, два : 10.1006/jmbi.2000.4403 , ПМИД 11178906 .
- ^ Перейти обратно: а б Б. Виллбрандт, Х. Собек, Б. Фрей, Д. Шомбург: Обмен доменов: химеры ДНК-полимеразы Thermus aquaticus, ДНК-полимеразы I Escherichia coli и ДНК-полимеразы Thermotoga neapolitana. В: Белок англ. , Том. 13, выпуск 9, 2000 г., с. 645–654. ПМИД 11054459 .
- ^ В. Абу Аль-Суд, П. Радстрем: Способность девяти термостабильных ДНК-полимераз опосредовать амплификацию ДНК в присутствии образцов, ингибирующих ПЦР. В: Прикл. Окружающая среда. Микробиол. , Том. 64, выпуск 10, 1998 г., с. 3748–3753. ПМИД 9758794 ; ПМЦ 106538 .
- ^ А. Гасеми, А. Х. Салманян, Н. Садегифард, А. А. Салариан, М. К. Голи: Клонирование, экспрессия и очистка полимеразы Pwo из Pyrococcus woesei. В: Иранский журнал микробиологии. Том 3, выпуск 3, сентябрь 2011 г., с. 118–122, ПМИД 22347593 , ПМК 3279813 .
- ^ Х. Конг, Р.Б. Кучера, В.Е. Джек: Характеристика ДНК-полимеразы гипертермофильной археи Thermococcuslitoralis. Вентиляционная ДНК-полимераза, кинетика устойчивого состояния, термическая стабильность, процессивность, смещение цепи и экзонуклеазная активность. В: J Biol Chem . , Том 268, Выпуск 3, 1993, с. 1965–1975. ПМИД 8420970 .
- ^ К. Бёлке, Ф. М. Пизани, К. Э. Воргиас, Б. Фрей, Х. Собек, М. Росси, Г. Антраникян: эффективность ПЦР ДНК-полимеразы B-типа термофильной эвриархеи Thermococcus aggregans улучшена за счет мутаций в Y-GG. / Мотив. В: Нуклеиновые кислоты Рез. , Том 28, Выпуск 20, 2000, с. 3910–3917. ПМИД 11024170 ; ПМК 110800 .
- ^ КП Ким, Х. Бэ, И. Х. Ким, с. Т. Квон: Клонирование, экспрессия и применение ПЦР ДНК-полимеразы гипертермофильного архея Thermococcus celer. В: Biotechnol Lett. (2011), том 33(2), с. 339–46. ПМИД 20953664 .
- ^ Ю. Чо, Х.п. Ли, Ю. Дж. Ким, с. Г. Канг, с. Дж. Ким, Дж. Х. Ли: Характеристика дУТФазы гипертермофильной археи Thermococcus onnurineus NA1 и ее применение в амплификации полимеразной цепной реакции . В: Mar Biotechnol (Нью-Йорк) , том 9, выпуск 4, 2007 г., с. 450–458. ПМИД 17549447 .
- ^ Перейти обратно: а б Дж.И. Ли, Ю.Дж. Ким, Х. Бэ, стр. Чо, Дж.Х. Ли, стр. Т. Квон: Биохимические свойства и эффективность ПЦР ДНК-полимеразы семейства B гипертермофильной эвриархеи Thermococcus peptonophilus. В: Appl Biochem Biotechnol. , Том 160, Выпуск 6, 2010, с. 1585–1899. ПМИД 19440663 .
- ^ Д. Марсик, Дж. М. Фламан, Дж. Д. Нг: Новая ДНК-полимераза гипертермофильного морского архея Thermococcus thioreducens. В: Экстремофилы , Том 12, Выпуск 6, 2008, с. 775–788. ПМИД 18670731 .
- ^ JG Song, EJ Kil, стр. Чо, IH Ким, стр. Т. Квон: Аминокислотный остаток в середине субдомена пальцев участвует в процессивности ДНК-полимеразы Neq: повышенная процессивность сконструированной ДНК-полимеразы Neq и ее применение в ПЦР. В: Белок англ. Дез. Сел. , Том 23, Выпуск 11, 2010 г., с. 835–842. ПМИД 20851826 .
- ^ Дж. Дитрих, П. Шмитт, М. Цигер, Б. Преве, Ж. Л. Роллан, Х. Чаабихи, Ю. Геген: Характеристики ПЦР высокотермостабильной корректурной ДНК-полимеразы B-типа из Pyrococcus abyssi. В: FEMS Microbiol Lett. , Том 217, Выпуск 1, 2002, с. 89–94. ПМИД 12445650 .
- ^ Э.М. Кеннеди, К. Херготт, с. Дьюхерст, Б. Ким: Механистическая архитектура термостабильного мотива ДНК-полимеразы A Pyrococcus Furiosus семейства B и его взаимодействие с субстратом dNTP. В: Биохимия (2009), том 48(47), с. 11161–8. ПМИД 19817489 ; ПМК 3097049 .
- ^ Перейти обратно: а б Т. Куроита, Х. Мацумура, Н. Ёкота, М. Китабаяши, Х. Хашимото, Т. Иноуэ, Т. Иманака, Ю. Кай: Структурный механизм координации корректурной и полимеразной активности ДНК-полимераз архей. В: J Мол Биол. (2005), том 351(2), с. 291–8. ПМИД 16019029 . дои:10.1016/j.jmb.2005.06.015 .
- ^ Дж. П. Макдональд, А. Холл, Д. Гаспарутто, Дж. Кадет, Дж. Баллантайн, Р. Вудгейт: Новые термостабильные полимеразы Y-семейства: применение для ПЦР-амплификации поврежденных или древних ДНК. В: Нуклеиновые кислоты Рез. (2006), том 34(4), с. 1102–11. ПМИД 16488882 ; ПМЦ 1373694 .
- ^ Е. Еремеева, П. Хердевейн: ПЦР-амплификация ДНК с модифицированным основанием. В: Современные протоколы химической биологии. Том 10, выпуск 1, март 2018 г., с. 18–48, два : 10.1002/cpch.33 , ПМИД 30040232 .
- ^ X. Ван, Дж. Чжан, Ю. Ли, Г. Чен, X. Ван: Ферментативный синтез модифицированных олигонуклеотидов с помощью PEAR с использованием ДНК-полимераз Phusion и KOD. В: Терапия нуклеиновыми кислотами. Том 25, выпуск 1, февраль 2015 г., с. 27–34, два : 10.1089/нат.2014.0513 , ПМИД 25517220 , ПМК 4296748 .
- ^ Ю. Ван, Д. Е. Просен, Л. Мей, Дж. К. Салливан, М. Финни, П. Б. Вандер Хорн: Новая стратегия разработки ДНК-полимераз для повышения процессивности и повышения производительности in vitro. В: Нуклеиновые кислоты Рез. (2004), том 32(3), с. 1197–207. ПМИД 14973201 ; ПМК 373405 .
- ^ М. Моц, И. Кобер, К. Жирардо, Э. Лозер, У. Бауэр, М. Альберс, Г. Мёкель, Э. Минч, Х. Восс, К. Килгер, М. Кегль: Выяснение репликации архей. белковая сеть для генерации улучшенных ферментов ПЦР . В: J Biol Chem. (2002), том 277(18), с. 16179–88. ПМИД 11805086 . PDF .
- ^ П. Фортер (2006), «ДНК-топоизомераза V: новая складка загадочного происхождения», Trends Biotechnol , vol. 24, нет. 6, стр. 245–247, doi : 10.1016/j.tibtech.2006.04.006 , PMID 16650908
- ^ А. Р. Павлов, Н. В. Павлова, С. А. Козявкин, А. И. Слесарев: Последние разработки в области оптимизации термостабильных ДНК-полимераз для эффективного применения. В: Тенденции в биотехнологии. Том 22, выпуск 5, май 2004 г., с. 253–260, дои : 10.1016/j.tibtech.2004.02.011 , ПМИД 15109812 .
- ^ Холли Х. Хогрефе, М. Борнс: Высокоточные ферменты ПЦР . В: К.В. Диффенбах, Г.П. Двекслер (ред.): Учебник для ПЦР: Лабораторное руководство , Лабораторное издательство Колд-Спринг-Харбор , Колд-Спринг-Харбор, Нью-Йорк, 2003.
- ^ Перейти обратно: а б с У. М. Барнс: ПЦР-амплификация ДНК размером до 35 т.п.н. с высокой точностью и высоким выходом на матрицах лямбда-бактериофага. В: Proc Natl Acad Sci USA (1994), том 91 (6), с. 2216–20. ПМИД 8134376 ; ПМЦ 43341 .
- ^ Бруно Фрей, Бернхард Суппманн: Демонстрация большей точности и более высоких выходов системы ПЦР Expand TM с помощью анализа точности ПЦР на основе lacI. (2000).
- ^ Р. Телье, Дж. Бух, С.У. Эмерсон, Р.Х. Перселл: Длинная ПЦР-амплификация больших фрагментов вирусных геномов: технический обзор. В кн.: Методы молекулярной биологии. Том 226, 2003 г., с. 167–172, дои : 10.1385/1-59259-384-4:167 , ПМИД 12958497 .
- ^ М. Миура, К. Танигава, Ю. Фуджи, С. Канеко: Сравнение шести коммерчески доступных ДНК-полимераз для прямой ПЦР. В: Revista do Instituto de Tropical Medicina de Sao Paulo. Том 55, выпуск 6, 2013 г., с. 401–406, два : 10.1590/S0036-46652013000600005 , ПМИД 24213192 , ПМК 4105087 .
- ^ Х.Х. Хогрефе, К.Дж. Хансен, Б.Р. Скотт, К.Б. Нильсон: Архейная дУТФаза усиливает ПЦР-амплификацию с помощью ДНК-полимераз архей, предотвращая включение dUTP. В: Труды Национальной академии наук . Том 99, выпуск 2, январь 2002 г., с. 596–601, два : 10.1073/pnas.012372799 , ПМИД 11782527 , ПМК 117351 .
- ^ Перейти обратно: а б Х. Карунанати, П.С. Ки, С.Ф. Нг, М.А. Кеннеди, Э.В. Чуа: Усилители ПЦР: типы, механизмы и приложения в ПЦР на большие расстояния. В: Биохимия. Том 197, июнь 2022 г., с. 130–143, два : 10.1016/j.biochi.2022.02.009 , ПМИД 35231536 .
- ^ Патент США 2013034879A1: ДНК-полимеразы , 02 августа 2012 г. / 14 февраля 2007 г., Fermentas UAB et al (заявитель), Remigijus Skirgaila et al. (изобретатель).
- ^ Патент США 2009280539A1: ДНК-полимеразы и родственные методы , 16 апреля 2009 г. / 12 ноября 2009 г., Roche Molecular Systems Inc (заявитель), Кейт А. Бауэр (изобретатель).
- ^ Дж. Ли, Ю. Ли, Ю. Ли, Ю. Ма, В. Сюй, Дж. Ван: Повышенная активность и термостабильность химерной ДНК-полимеразы Bst для приложений изотермической амплификации. В кн.: Прикладная микробиология и биотехнология . Том 107, выпуск 21, ноябрь 2023 г., с. 6527–6540, два : 10.1007/s00253-023-12751-6 , ПМИД 37672070 .
- ^ WM Barnes: Точность Taq-полимеразы, катализирующей ПЦР, повышается за счет делеции N-конца . В: Джин (1992), том 112 (1), с. 29–35. ПМИД 1551596 .
- ^ Л. п. Меркенс, с. К. Брайан, Р.Э. Мозес: Инактивация 5'-3' экзонуклеазы ДНК-полимеразы Thermus aquaticus. В: Biochim Biophys Acta (1995), том 1264(2), с. 243–8. ПМИД 7495870 .
- ^ с. Табор, К.С. Ричардсон: Один остаток в ДНК-полимеразах семейства ДНК-полимераз I Escherichia coli имеет решающее значение для различения дезокси- и дидезоксирибонуклеотидов. В: Proc Natl Acad Sci US A. , том 92, выпуск 14, 1995, с. 6339–6343. ПМИД 7603992 ; ПМЦ 41513 .
- ^ П. Б. Вандер Хорн, М. К. Дэвис, Дж. Дж. Каннифф, К. Руан, Б. Ф. Макардл, стр. 10. Б. Самолс, Й. Сас, Г. Ху, К.М. Худжер, с. Т. Домке, с. Р. Браммет, Р.Б. Моффетт, К.В. Фуллер: ДНК-полимераза термосеквеназы и пирофосфатаза T. acidophilum: новые термостабильные ферменты для секвенирования ДНК. В: Биотехника , Том 22, Выпуск 4, 1997, с. 758–762, 764–765. ПМИД 9105629 .
- ^ Дж. М. Пробер, Г. Л. Трейнор, Р. Дж. Дам, Ф. В. Хоббс, К. В. Робертсон, Р. Дж. Загурски, А. Дж. Кокуцца, М. А. Дженсен, К. Баумайстер: Система для быстрого секвенирования ДНК с флуоресцентными дидезоксинуклеотидами, обрывающими цепь. В: Science , том 238, выпуск 4825, 1987, с. 336–341. ПМИД 2443975 .
- ^ RT D'Aquila, LJ Bechtel, JA Videler, JJ Eron, P. Gorczyca, JC Kaplan: Максимизация чувствительности и специфичности ПЦР за счет нагревания перед амплификацией. В: Исследования нуклеиновых кислот. Том 19, выпуск 13, июль 1991 г., с. 3749, два : 10.1093/nar/19.13.3749 , ПМИД 1852616 , ПМК 328414 .
- ^ С. Буратовский: Очистка Taq с горячим стартом . 2015.
- ^ TG Graham, C. Dugast-Darzacq, GM Dailey, XH Nguyenla, E. Van Dis, MN Esbin, A. Abidi, SA Stanley, X. Darzacq, R. Tjian: Методы экстракции РНК с открытым исходным кодом и методы RT-qPCR для Обнаружение SARS-CoV-2. В: ПЛОС ОДИН . Том 16, выпуск 2, 2021 г., с. е0246647, дои : 10.1371/journal.pone.0246647 , ПМИД 33534838 , ПМК 7857565 .
- ^ Дэвид Эдвард Берч, Уолтер Джозеф Лэрд, Майкл Энтони Зокколи: амплификация нуклеиновой кислоты с использованием обратимо инактивированного термостабильного фермента . Патент США 5773258.
- ^ Перейти обратно: а б В.М. Барнс, К.Р. Роулик: Метод горячего старта с осадком магния для ПЦР. В: Молекулярные и клеточные зонды. Том 16, выпуск 3, июнь 2002 г., с. 167–171, два : 10.1006/mcpr.2002.0407 , ПМИД 12219733 .
- ^ Н. Пол, Дж. Шум, Т. Ле: ПЦР с горячим стартом . В кн.: Методы Мол Биол. (2010), том 630, с. 301–18. ПМИД 20301005 .
- ^ М.Ф. Крамер, Д.М. Коэн: Ферментативная амплификация ДНК с помощью ПЦР: стандартные процедуры и оптимизация . В: Curr Protoc Immunol. (2001), Глава 10, Раздел 10.20. ПМИД 18432685 .
- ^ Х. ФРЕНКЕЛЬ-КОНРАТ, Б.А. БРЭНДОН, Х.С. ОЛКОТТ: Реакция формальдегида с белками; участие индольных групп; грамицидин. В: Журнал биологической химии. Том 168, выпуск 1, апрель 1947 г., ISSN 0021-9258 , с. 99–118, ПМИД 20291066 .
- ^ Х. ФРЕНКЕЛЬ-КОНРАТ, Х.С. ОЛКОТТ: Реакция формальдегида с белками; сшивание между амино и первичными амидными или гуанидиловыми группами. В: Журнал Американского химического общества. Том 70, выпуск 8, август 1948 г., ISSN 0002-7863 , стр. 2673–2684, ПМИД 18876976 .
- ^ Х. ФРЕНКЕЛЬ-КОНРАТ, Х.С. ОЛКОТТ: Реакция формальдегида с белками; сшивание аминогрупп фенольными, имидазольными или индольными группами. В: Журнал биологической химии. Том 174, выпуск 3, июль 1948 г., ISSN 0021-9258 , с. 827–843, ПМИД 18871242 .
- ^ Д.Э. Келлог, И. Рыбалкин, С. Чен, Н. Мухамедова, Т. Власик, П.Д. Зиберт, А. Ченчик: Антитела TaqStart: ПЦР «горячего старта», облегченная нейтрализующим моноклональным антителом, направленным против ДНК-полимеразы Taq. В: Биотехника. Том 16, выпуск 6, июнь 1994 г., с. 1134–1137, ПМИД 8074881 .
- ^ DJ Sharkey, ER Scalice, KG Christy, SM Atwood, JL Daiss: Антитела как термолабильные переключатели: запуск полимеразной цепной реакции при высокой температуре. В: Био/технологии. Том 12, выпуск 5, май 1994 г., с. 506–509, два : 10.1038/nbt0594-506 , PMID 7764710 .
- ^ O. Y. Yakimovich, Y. I. Alekseev, A. V. Maksimenko, O. L. Voronina, V. G. Lunin: Influence of DNA aptamer structure on the specificity of binding to Taq DNA polymerase. In: Biochemistry. Biokhimiia. Volume 68, issue 2, February 2003, p. 228–235, два : 10.1023/a:1022609714768 , ПМИД 12693970 .
- ^ Т. Нома, К. Соде, К. Икебукуро: Характеристика и применение аптамеров для ДНК-полимеразы Taq, выбранных с использованием алгоритма, имитирующего эволюцию. В: Биотехнологические письма. Том 28, выпуск 23, декабрь 2006 г., с. 1939–1944, два : 10.1007/s10529-006-9178-4 , ПМИД 16988782 .
- ^ К. Чоу, М. Рассел, Д. Е. Берч, Дж. Рэймонд, В. Блох: Предотвращение неправильного прайминга перед ПЦР и димеризации праймера улучшает амплификацию с низким числом копий. В: Нуклеиновые кислоты Рез. (1992), том 20, вып. 7, с. 1717–23. ПМИД 1579465 ; ПМК 312262 .
- ^ Карлос Д. Ордоньес, Модесто Редрехо-Родригес: ДНК-полимеразы для амплификации всего генома: соображения и будущие направления. В: Международный журнал молекулярных наук . 2023, том 24, выпуск 11, с. 9331 два : 10.3390/ijms24119331 . ПМИД 37298280 . ПМЦ 10253169 .
- ^ Б. Фукс, К. Чжан, М. Г. Рок, М. Е. Боландер, Г. Саркар: Высокотемпературный синтез кДНК с помощью обратной транскриптазы AMV повышает специфичность ПЦР. В кн.: Молекулярная биотехнология. Том 12, выпуск 3, октябрь 1999 г., с. 237–240, дои : 10.1385/МБ:12:3:237 , ПМИД 10631680 .
- ^ А. М. Карозерс, Г. Урлауб, Дж. Муха, Д. Грюнбергер, Л. А. Часин: Анализ точечных мутаций в гене млекопитающих: быстрое получение тотальной РНК, ПЦР-амплификация кДНК и секвенирование Taq новым методом. В: Биотехника. Том 7, выпуск 5, май 1989 г., с. 494–6, 498, ПМИД 2483818 .
- ^ Т.В. Майерс, Д.Х. Гельфанд: Обратная транскрипция и амплификация ДНК с помощью ДНК-полимеразы Thermus thermophilus. В: Биохимия (1991), Band 30(31), S. 7661–6. ПМИД 1714296 .
- ^ М.Дж. Мозер, Р.А. ДиФранческо, К. Гауда, А.Дж. Клингеле, Д.Р. Шугар, С. Стоки, Д.А. Мид, Т.В. Шенфельд: Термостабильная ДНК-полимераза из вирусного метагенома является мощным ферментом ОТ-ПЦР. В: PLoS One (2012), том 7(6), с. е38371. дои : 10.1371/journal.pone.0038371 . ПМИД 22675552 ; ПМК 3366922 .
- ^ Чиен А., Эдгар Д.Б., Трела Дж.М. (сентябрь 1976 г.). «Полимераза дезоксирибонуклеиновой кислоты крайнего термофила Thermus aquaticus» . Журнал бактериологии . 127 (3): 1550–1557. дои : 10.1128/JB.127.3.1550-1557.1976 . ПМК 232952 . ПМИД 8432 .
- ^ Р.К. Сайки, Д.Х. Гельфанд, П. Стоффель, П.Дж. Шарф, Р. Хигучи, Г.Т. Хорн, К.Б. Муллис, Х.А. Эрлих: Ферментативная амплификация ДНК с помощью праймера с помощью термостабильной ДНК-полимеразы. В: Наука . Том 239, выпуск 4839, январь 1988 г., с. 487–491, дои : 10.1126/science.2448875 , ПМИД 2448875 .
- ^ Перейти обратно: а б с д П. Ишино, Ю. Ишино: ДНК-полимеразы как полезные реагенты для биотехнологии - история исследований в этой области. В: Границы микробиологии. Том 5, 2014, с. 465, два : 10.3389/fmicb.2014.00465 , ПМИД 25221550 , ПМК 4148896 .
- ^ Юрист ФК, П. Стоффель, Р.К. Сайки, К. Мьямбо, Р. Драммонд, Д. Х. Гельфанд: Выделение, характеристика и экспрессия в Escherichia coli гена ДНК-полимеразы из Thermus aquaticus. В: Журнал биологической химии . Том 264, выпуск 11, апрель 1989 г., с. 6427–6437, ПМИД 2649500 .
- ^ М. Сорока, Б. Васович, А. Рымашевска: Петлевая изотермическая амплификация (LAMP): лучший брат ПЦР? В: Клетки. Том 10, выпуск 8, июль 2021 г., с. , дои : 10.3390/cells10081931 , ПМИД 34440699 , ПМК 8393631 .
- ^ Нотоми Т., Окаяма Х., Масубучи Х., Ёнекава Т., Ватанабе К., Амино Н., Хасэ Т. (2000). «Петлевая изотермическая амплификация ДНК» . Нуклеиновые кислоты Рез . 28 (12): 63д–63. дои : 10.1093/нар/28.12.e63 . ПМЦ 102748 . ПМИД 10871386 .