Генетическая связь
Генетическая связь - это тенденция последовательностей ДНК , которые находятся близко друг к другу на хромосоме , которая должна быть унаследована вместе во время мейоза фазы сексуального размножения . Два генетических маркера , которые физически близки друг к другу, вряд ли будут разделены на разные хроматиды во время хромосомного кроссовера , и, следовательно, считаются более связанными, чем маркеры, которые находятся далеко друг от друга. Другими словами, чем более ближе два гена находятся на хромосоме, тем ниже вероятность рекомбинации между ними, и тем больше вероятность, что они будут унаследовать вместе. Маркеры на разных хромосомах совершенно не связаны , хотя на проникновение потенциально вредных аллелей может влиять присутствие других аллелей, и на эти другие аллели могут располагаться на других хромосомах, чем на конкретном потенциально долговечном аллеле. [ 1 ]
Генетическая связь является наиболее выдающимся исключением из Грегора Менделя закона о независимом ассортименте . Первый эксперимент, демонстрирующий связь, был проведен в 1905 году. В то время причина, по которой определенные признаки, как правило, унаследованы вместе, была неизвестна. Более поздняя работа показала, что гены - это физические структуры, связанные с физическим расстоянием.
Типичной единицей генетической связи является сантиморган (CM). Расстояние 1 см между двумя маркерами означает, что маркеры разделены на различные хромосомы в среднем один раз на 100 мейотических продуктов, таким образом, один раз на 50 мейозов.
Открытие
[ редактировать ]Грегора Менделя гласит Закон независимого ассортимента , что каждая черта унаследована независимо от любой другой черты. Но вскоре после того, как работа Менделя была вновь открыта , были найдены исключения из этого правила. В 1905 году британские генетики Уильям Бейтсон , Эдит Ребекка Сондерс и Реджинальд Паннетт, перекрестные растения, в экспериментах, аналогичных Менделу. [ 2 ] [ 3 ] Они были заинтересованы в наследстве черты в сладком гороше и изучали два гена - ген для цвета цветов ( P , фиолетовый и P , красный) и ген, влияющий на форму пыльцевых зерен ( L , Long и L , круглый) Полем Они пересекли чистые линии PPLL и PPLL , а затем самостоятельно перекрещивали полученные линии PPLL .
Согласно Mendelian Genetics , ожидаемые фенотипы будут происходить в соотношении 9: 3: 3: 1 PL: PL: PL: PL. К их удивлению, они наблюдали повышенную частоту PL и PL и уменьшенную частоту PL и PL:
| Фенотип и генотип | Наблюдается | Ожидается от 9: 3: 3: 1 |
|---|---|---|
| Фиолетовый, длинный ( p_l_ ) | 284 | 216 |
| Фиолетовый, круглый ( p_ll ) | 21 | 72 |
| Красный, длинная ( ppl_ ) | 21 | 72 |
| Красный, круглый ( ppll ) | 55 | 24 |
Их эксперимент выявил связь между аллелями P и L и P и L. аллелями Частота P встречается вместе с L и P, встречающимися вместе с L, больше, чем у рекомбинантного PL и PL . Частоту рекомбинации труднее вычислять на кресте F2, чем обратная скрещивание, [ 4 ] Но отсутствие соответствия между наблюдаемым и ожидаемым количеством потомства в вышеуказанной таблице указывает на то, что оно составляет менее 50%. Это указывало на то, что два фактора каким -то образом взаимодействовали, чтобы создать эту разницу, маскируя внешний вид двух других фенотипов. Это привело к выводу, что некоторые признаки связаны друг с другом из -за их почти близости друг к другу на хромосоме.
Понимание связи было расширено работой Томаса Ханта Моргана . Наблюдение за Морганом о том, что количество пересечения между связанными генами отличается, привело к идее, что частота кроссовера может указывать на расстояние, разделяющие гены на хромосоме . Семьморган , который выражает частоту пересечения, названа в его честь.
Карта сцепления
[ редактировать ]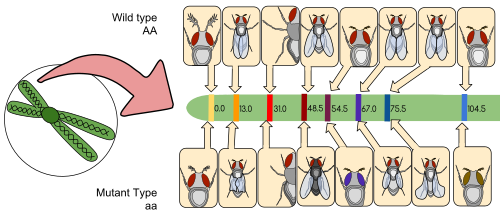
Карта сцепления (также известная как генетическая карта ) представляет собой таблицу для вида или экспериментальной популяции, которая показывает положение его известных генов или генетических маркеров относительно друг друга с точки зрения частоты рекомбинации, а не определенного физического расстояния вдоль каждой хромосомы Полем Карты связи были впервые разработаны Альфредом Стертевантом , студентом Томаса Ханта Моргана .
Карта сцепления - это карта, основанная на частотах рекомбинации между маркерами во время кроссовера гомологичных хромосом . Чем больше частота рекомбинации (сегрегации) между двумя генетическими маркерами, тем дальше они предполагаются. И наоборот, чем ниже частота рекомбинации между маркерами, тем меньше физическое расстояние между ними. Исторически, первоначально использованные маркеры были обнаруживаемыми фенотипами (производство ферментов, цвет глаз), полученные из кодирующих последовательностей ДНК; подтвержденные или предполагаемые некодирующие последовательности ДНК, такие как микросателлиты или те, которые генерируют полиморфизмы длины ограничения фрагмента ( RFLP В конечном итоге были использованы ).
Карты сцепления помогают исследователям найти другие маркеры, такие как другие гены, тестируя генетическую связь уже известных маркеров. На ранних стадиях разработки карты сцепления данные используются для сборки групп сцепления , набор генов, которые, как известно, связаны. По мере продвижения знаний, в группу можно добавить больше маркеров, пока группа не покрывает целую хромосому. [ 6 ] Для хорошо изученных организмов группы сцепления соответствуют один-один с хромосомами.
Карта сцепления не является физической картой (например, радиационной гибридной картой) или картой генов .
Анализ сцепления
[ редактировать ]Анализ сцепления - это генетический метод, который ищет хромосомные сегменты, которые косегрегируют с фенотипом болезни через семейства. [ 7 ] Его можно использовать для отображения генов как для бинарных, так и для количественных признаков. [ 7 ] Анализ сцепления может быть либо параметрическим (если мы знаем взаимосвязь между фенотипическим и генетическим сходством), либо непараметрическим. Анализ параметрического сцепления - это традиционный подход, посредством которого вероятность того, что ген, важный для заболевания, связан с генетическим маркером, изучается через оценку LOD, которая оценивает вероятность того, что данная родословная, где заболевание и маркер являются косегрегацией, является из -за существования связи (с данным значением связи) или случайной. Непараметрический анализ связей, в свою очередь, изучает вероятность того, что аллель будет идентичен по себе спусканием .

Анализ параметрического сцепления
[ редактировать ]Оценка LOD (логарифм (база 10) шансов), разработанный Ньютоном Мортоном , [ 8 ] является статистическим тестом, часто используемым для анализа связей в популяциях человека, животных и растений. Оценка LOD сравнивает вероятность получения тестовых данных, если два локуса действительно связаны, с вероятностью наблюдения одних и тех же данных исключительно случайно. Положительные оценки LOD предпочитают наличие связи, тогда как отрицательные оценки LOD указывают на то, что связь менее вероятно. Компьютерный анализ оценки LOD - это простой способ проанализировать сложные родословные семейства, чтобы определить связь между мендельными чертами (или между чертой и маркером, или двумя маркерами).
Метод более подробно описывается Страчаном и чтением. [1] Кратко, это работает следующим образом:
- Установить родословную
- Сделайте ряд оценок частоты рекомбинации
- Рассчитайте оценку LOD для каждой оценки
- Оценка с самым высоким показателем LOD будет считаться лучшей оценкой
Оценка LOD рассчитывается следующим образом:

NR обозначает количество нерекомбинантных потомков, а R обозначает количество рекомбинантных потомков. Причина 0,5 используется в знаменателе, заключается в том, что любые аллели, которые полностью не связаны (например, аллели на отдельных хромосомах), имеют 50% вероятность рекомбинации из -за независимого ассортимента. θ - это рекомбинантная фракция, то есть доля рождений, в которой произошла рекомбинация между изученным генетическим маркером и предполагаемым геном, связанным с заболеванием. Таким образом, это равно r / ( nr + r ) .
По соглашению, оценка LOD, превышающий 3,0, считается доказательством связи, так как это указывает на от 1000 до 1 шансов на то, что наблюдаемая связь не возникала случайно. С другой стороны, оценка LOD меньше -2,0 считается доказательством исключения связи. Хотя очень маловероятно, что оценка LOD 3 будет получена из одной родословной, математические свойства теста позволяют комбинировать данные из ряда родословных, суммируя их оценки LOD. Оценка LOD 3 переводится на p -значение приблизительно 0,05, [ 9 ] И не многократная коррекция тестирования (например, коррекция Бонферрони ). требуется [ 10 ]
Ограничения
[ редактировать ]Анализ сцепления имеет ряд методологических и теоретических ограничений, которые могут значительно увеличить частоту ошибок типа 1 и уменьшить мощность для картирования локусов количественных признаков человека (QTL). [ 11 ] В то время как анализ связей был успешно использован для выявления генетических вариантов, которые способствуют редким расстройствам, таким как болезнь Хантингтона , он не очень хорошо работал при применении к более распространенным расстройствам, таким как болезни сердца или различные формы рака . [ 12 ] Объяснение этого заключается в том, что генетические механизмы, влияющие на общие расстройства, отличаются от тех, что вызывают некоторые редкие расстройства. [ 13 ]
Частота рекомбинации
[ редактировать ]Частота рекомбинации является мерой генетической связи и используется при создании карты генетической связи. Частота рекомбинации ( θ ) - это частота, с которой один хромосомный кроссовер будет проходить между двумя генами во время мейоза . Сантиморган ( CM ) - это единица, которая описывает частоту рекомбинации 1%. Таким образом, мы можем измерить генетическое расстояние между двумя локусами, основываясь на их частоте рекомбинации. Это хорошая оценка реального расстояния. Двойные кроссоверы не превратятся в рекомбинацию. В этом случае мы не можем сказать, произошли ли кроссоверы. Если локусы, которые мы анализируем, очень близки (менее 7 см), двойной кроссовер очень маловероятно. Когда расстояния становятся выше, вероятность двойного кроссовера увеличивается. По мере увеличения вероятности двойного кроссовера можно систематически недооценивать генетическое расстояние между двумя локусами, если только кто -то не использовал подходящую математическую модель.
Двойная связь - это более историческая забота о растениях. У животных двойной кроссовер случается редко. Например, у людей одна хромосома имеет два кроссовера в среднем во время мейоза. Кроме того, у современных генетиков достаточно генов, чтобы только близлежащие гены должны быть проведены аналитическими, в отличие от первых дней, когда было известно только несколько генов. [ 14 ]
Во время мейоза хромосомы случайным образом ассорти в гаметы , так что сегрегация аллелей одного гена не зависит от аллелей другого гена. Это указано во втором законе Менделя и известно как закон независимого ассортимента . Закон независимого ассортимента всегда верен для генов, которые расположены на разных хромосомах, но для генов, которые находятся на одной и той же хромосоме, он не всегда верно.
В качестве примера независимого ассортимента рассмотрим пересечение чисточеного гомозиготного штамма родительского языка с генотипом AABB с другим пятном штаммом с генотипом AABB . A и A и B и B представляют аллели генов A и B., пересекающие эти гомозиготные родительские штаммы приведут к потомству Generation F1, которые являются двойными гетерозиготами с генотипом AABB. Потомство F1 AABB производит гаметы, которые являются AB , AB , AB и AB с равными частотами (25%), потому что аллели гена A ассорти независимо от аллелей для гена B во время мейоза. Обратите внимание, что 2 из 4 гамет (50%) - AB и AB - не присутствовали в родительском поколении. Эти гамет представляют рекомбинантные гаметы . Рекомбинантные гаметы - это те гаметы, которые отличаются от обоих гаплоидных гамет, которые составляли оригинальную диплоидную ячейку. В этом примере частота рекомбинации составляет 50%, поскольку 2 из 4 гамет были рекомбинантными гаметами.
Частота рекомбинации будет составлять 50%, когда два гена расположены на разных хромосомах или когда они широко разделены на одной и той же хромосоме. Это следствие независимого ассортимента.
Когда два гена находятся близко друг к другу на одной и той же хромосоме, они не ассорти независимо и говорят, что они связаны. Принимая во внимание, что гены, расположенные на различных хромосомах, независимо от ассорти, и имеют частоту рекомбинации 50%, связанные гены имеют частоту рекомбинации, которая составляет менее 50%.
В качестве примера связи, рассмотрите классический эксперимент Уильяма Бейтсона и Реджинальда Паннетта . [ 15 ] Они были заинтересованы в наследстве черты в сладком гороше и изучали два гена - ген для цвета цветов ( P , фиолетовый и P , красный) и ген, влияющий на форму пыльцевых зерен ( L , Long и L , круглый) Полем Они пересекли чистые линии PPLL и PPLL , а затем самостоятельно перекрещивали полученные линии PPLL . Согласно Mendelian Genetics , ожидаемые фенотипы будут происходить в соотношении 9: 3: 3: 1 PL: PL: PL: PL. К их удивлению, они наблюдали повышенную частоту PL и PL и уменьшенную частоту PL и PL (см. Таблицу ниже).
| Фенотип и генотип | Наблюдается | Ожидается от 9: 3: 3: 1 |
|---|---|---|
| Фиолетовый, длинный ( p_l_ ) | 284 | 216 |
| Фиолетовый, круглый ( p_ll ) | 21 | 72 |
| Красный, длинная ( ppl_ ) | 21 | 72 |
| Красный, круглый ( ppll ) | 55 | 24 |

Их эксперимент выявил связь между аллелями P и L и P и L. аллелями Частота P , встречающаяся вместе с L и с P, встречающейся вместе с L, больше, чем у рекомбинантного PL и PL . Частоту рекомбинации труднее вычислять на кресте F2, чем обратная скрещивание, [ 4 ] Но отсутствие соответствия между наблюдаемым и ожидаемым количеством потомства в вышеуказанной таблице указывает на то, что оно составляет менее 50%.
Потомство в этом случае получило два доминирующих аллеля, связанных с одной хромосомой (называемой связью или расположением CIS ). Однако после кроссовера некоторое потомство могло бы получить одну родительскую хромосому с доминирующим аллелем для одной черты (например, фиолетового цвета), связанного с рецессивным аллелем для второй черты (например, раунд), а наоборот, истинная для другой родительской хромосомы (например, красный и долго). Это называется отталкиванием или транс -композицией . Фенотип здесь все еще будет фиолетовый и длинный , но испытательный крест этого человека с рецессивным родителем будет производить потомство с гораздо большей долю двух кроссоверных фенотипов. Хотя такая проблема не может показаться вероятной из этого примера, неблагоприятные отталкивающие связи появляются при размножении для устойчивости к болезням в некоторых культурах.
Два возможных договоренности, цис и транс, аллелей в двойной гетерозиготе, называются гаметическими фазами , а фазирование - это процесс определения того, кто из двух присутствует у данного человека.
Когда два гена расположены на одной и той же хромосоме, вероятность кроссовера , продуцирующей рекомбинацию между генами, связана с расстоянием между двумя генами. Таким образом, использование частот рекомбинации использовалось для разработки карт сцепления или генетических карт .
Однако важно отметить, что частота рекомбинации имеет тенденцию недооценивать расстояние между двумя связанными генами. Это связано с тем, что по мере того, как два гена расположены дальше друг от друга, вероятность двойного или даже количества кроссоверов между ними также увеличивается. Двойное или даже количество кроссоверов между двумя генами приводит к тому, что они становятся косегрегированными по одному гамете, что дает родительское потомство вместо ожидаемого рекомбинантного потомства. Как упомянуто выше, преобразования Косамби и Холдейна пытаются исправить для нескольких кроссоверов. [ 16 ]
Связь генетических сайтов внутри гена
[ редактировать ]В начале 1950 -х годов преобладающим мнением было то, что гены в хромосоме являются дискретными сущностями, неделимыми генетической рекомбинацией и расположены как бусины на струне. В течение 1955 года по 1959 год Бензер провел эксперименты по генетической рекомбинации , используя RII мутанты бактериофага T4 . Он обнаружил, что на основе тестов на рекомбинацию сайты мутации могут быть нанесены на карту в линейном порядке. [ 17 ] [ 18 ] Этот результат предоставил доказательства ключевой идеи о том, что ген имеет линейную структуру, эквивалентную длине ДНК со многими сайтами, которые могут независимо мутировать.
Edgar et al. [ 19 ] Выполненные картирование экспериментов с r мутантами бактериофага T4, показывающие, что частоты рекомбинации между мутантами RII не являются строго добавленными. Частота рекомбинации из скрещивания двух мутантов RII (AXD) обычно меньше, чем сумма частот рекомбинации для соседних внутренних подметок (AXB) + (BXC) + (CXD). Хотя это и не аддитивно, наблюдалась систематическая связь [ 20 ] Это, вероятно, отражает основной молекулярный механизм генетической рекомбинации .
Изменение частоты рекомбинации
[ редактировать ]В то время как рекомбинация хромосом является важным процессом во время мейоза, существует большой диапазон частоты перекрестных оверов по организмам и внутри видов. Половой диморфической рекомбинации называется гетерохиамией и наблюдается чаще, чем общая скорость между мужчинами и женщинами. У млекопитающих женщины часто имеют более высокую скорость рекомбинации по сравнению с мужчинами. Теоретизируется, что существуют уникальные выборы, действующие или мейотические драйверы, которые влияют на разницу в показателях. Разница в скоростях может также отражать совершенно разные среды и условия мейоза в оогенезе и сперматогенезе. [ 21 ]
Гены, влияющие на частоту рекомбинации
[ редактировать ]Мутации в генах , которые кодируют белки, участвующие в обработке ДНК, часто влияют на частоту рекомбинации . У бактериофага T4 мутации, которые снижают экспрессию репликативной ДНК -полимеразы [генного продукта 43 (GP43)], увеличивают рекомбинацию (уменьшение связи) в несколько раз. [ 22 ] [ 23 ] Увеличение рекомбинации может быть связано с ошибками репликации дефектной ДНК -полимеразой, которые сами являются событиями рекомбинации, такие как переключатели шаблонов, то есть события рекомбинации выбора копии. [ 24 ] Рекомбинация также увеличивается мутациями, которые снижают экспрессию ДНК -лигазы (GP30) [ 25 ] [ 23 ] и DCMP гидроксиметилаза (GP42), [ 22 ] [ 23 ] два фермента, используемые в синтезе ДНК .
Рекомбинация снижается (сцепление увеличивается) мутациями в генах, которые кодируют белки с нуклеазы (GP46 и GP47) функциями [ 25 ] [ 23 ] и ДНК-связывающий белок (GP32) [ 23 ] Мутация в гене UVSX бактериофага также существенно снижает рекомбинацию. [ 26 ] Ген UVSX аналогичен хорошо изученному Reca гену Escherichia coli , который играет центральную роль в рекомбинации. [ 27 ]
Мейоз показатели
[ редактировать ]С очень большими родословными или с очень плотными данными генетических маркеров, например, из секвенирования всего генома, можно точно определить рекомбинации. С этим типом генетического анализа индикатор мейоза назначается каждому положению генома для каждого мейоза в родословной. Индикатор указывает, какая копия родительской хромосомы способствует переданной гамете в этой позиции. Например, если передается аллель из «первой» копии родительской хромосомы, это может быть назначен «0». Если аллель из «второй» копии родительской хромосомы передается, на этот мейоз будет назначен «1». Два аллеля в родителе пришли, по одному, от двух бабушек и дедушек. Затем эти показатели используются для определения идентичных состояний (IBD) или состояний наследования, которые, в свою очередь, используются для идентификации генов, ответственных за заболевания.
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Купер, Дн; Krawczak, M; Polychronakos, c; Тайлер-Смит, C; Керер-Савацки, ч (октябрь 2013 г.). «Где генотип не является предиктором фенотипа: к пониманию молекулярной основы снижения пенетрантности при наследственном заболевании человека» . Человеческая генетика . 132 (10): 1077–130. doi : 10.1007/s00439-013-1331-2 . PMC 3778950 . PMID 23820649 .
- ^ Лобо, Ингрид; Шоу, Кенна. «Открытие и типы генетической связи» . Scaute . Природное образование . Получено 21 января 2017 года .
- ^ Бейтсон, w ; Сондерс, эр ; Паннетт, RC (18 мая 1904 г.). Отчеты Комитету эволюции Королевского общества . Лондон: Харрисон и сыновья, принтеры . Получено 21 января 2017 года .
- ^ Jump up to: а беременный Фишер, Ра ; Balmukand, B (июль 1928 г.). «Оценка связи от потомства самооткрытых гетерозигот». Журнал генетики . 20 (1): 79–92. doi : 10.1007/bf02983317 . S2CID 27688031 .
- ^ Мадер, Сильвия (2007). Биология девятое издание . Нью-Йорк: МакГроу-Хилл. п. 209. ISBN 978-0-07-325839-3 .
- ^ Griffiths, AJF (2000). Введение в генетический анализ (7 -е изд.). WH Freeman.
- ^ Jump up to: а беременный Кантор, Рита М. (2013), Римин, Дэвид; Пириц, Рид; Брюс. , Корф 1–9, doi : 10.1016/b978-12-38344–6.0010-0 , ISBN 9780123838346
- ^ Мортон Н.Е. (1955). «Последовательные тесты для обнаружения связи» . Американский журнал человеческой генетики . 7 (3): 277–318. PMC 1716611 . PMID 13258560 .
- ^ Nyholt, Dale R (август 2000 г.). «Все LOD не созданы равными» . Американский журнал человеческой генетики . 67 (2): 282–288. doi : 10.1086/303029 . PMC 1287176 . PMID 10884360 .
- ^ Риш, Нил (июнь 1991 г.). «Примечание о нескольких процедурах тестирования в анализе сцепления» . Американский журнал человеческой генетики . 48 (6): 1058–1064. PMC 1683115 . PMID 2035526 .
- ^ Ferreira, Manuel AR (2004-10-01). «Анализ связей: принципы и методы анализа количественных признаков человека». Двойные исследования и генетика человека . 7 (5): 513–530. doi : 10.1375/twin.7.5.513 . ISSN 2053-6003 . PMID 15527667 . S2CID 199001341 .
- ^ Гуселла, Джеймс Ф.; Фронтали, Марина; Wasmuth, John J.; Коллинз, Фрэнсис С.; Лерач, Ганс; Майерс, Ричард; Альтерр, Майкл; Аллитто, Бернис; Тейлор, Шерри (1992-05-01). «В регионе кандидата в болезнь Хантингтона есть много разных гаплотипов». Природа генетика . 1 (2): 99–103. doi : 10.1038/ng0592-99 . ISSN 1546-1718 . PMID 1302016 . S2CID 25472459 .
- ^ Марк Дж. Дейли; Хиршхорн, Джоэл Н. (2005-02-01). «Обще геномские ассоциации исследований для общих заболеваний и сложных признаков». Nature Reviews Genetics . 6 (2): 95–108. doi : 10.1038/nrg1521 . ISSN 1471-0064 . PMID 15716906 . S2CID 2813666 .
- ^ Менели, Филипп Марк; Доус Хоанг, Рэйчел; Океке, Ирука Н.; Хестон, Кэтрин (2017). Генетика: гены, геномы и эволюция . Оксфорд: издательство Оксфордского университета. п. 361. ISBN 978-0-19-879536-0 Полем OCLC 951645141 .
- ^ Паннетт, RC; Бейтсон, В. (1908-05-15). «Наследственность секса» . Наука . 27 (698): 785–787. Bibcode : 1908sci .... 27..785p . doi : 10.1126/science.27.698.785 . ISSN 0036-8075 . PMID 17791047 .
- ^ Гриффитс, AJF; Миллер, JH; Suzuki, DT (2000). «Точный расчет больших расстояний карты, рисунок 6-4» . Введение в генетический анализ (7 -е изд.). Нью -Йорк: WH Freeman . ISBN 978-0-7167-3520-5 Полем График функции отображения по сравнению с идеализированной эквивалентностью частоты рекомбинации (RF%) для MAP единиц.
- ^ Бензер С. Прекрасная структура генетической области у бактериофагов. Proc Natl Acad Sci US A. 1955; 41 (6): 344-354. doi: 10.1073/pnas.41.6.344
- ^ Бензер С. О топологии генетической тонкой структуры. Proc Natl Acad Sci US A. 1959; 45 (11): 1607-1620. doi: 10.1073/pnas.45.11.1607
- ^ Эдгар Р.С., Фейнман Р.П., Кляйн С., Лиелаузис I, Стейнберг С.М. Картирование экспериментов с R -мутантами бактериофага T4D. Генетика. 1962; 47: 179–186. PMC 1210321 . PMID 13889186
- ^ Фишер К.М., Бернштейн Х. Додажность интервалов в RIA Cistron of Phage T4D. генетика. 1965; 52 (6): 1127–1136. 1210971PMC PMID 5882191
- ^ Макки, Брюс Д. (2004-03-15). «Гомологичная пара и динамика хромосом при мейозе и митозе» . Biochimica et Biophysica Acta (BBA) - структура и экспрессия гена . 1677 (1–3): 165–180. doi : 10.1016/j.bbaexp.2003.11.017 . ISSN 0006-3002 . PMID 15020057 .
- ^ Jump up to: а беременный Бернштейн Х. Влияние на рекомбинацию мутационных дефектов в ДНК-полимеразе и дезоксицитидилат-гидроксиметилазе фага T4D. Генетика. 1967; 56 (4): 755-769
- ^ Jump up to: а беременный в дюймовый и Бергер Х, Уоррен А.Дж., Фрай К. Вариации в генетической рекомбинации из -за мутаций янтаря в бактериофагах T4D. J Virol. 1969; 3 (2): 171-175. doi: 10.1128/jvi.3.2.171-175.1969
- ^ Бернштейн Х. О механизме внутригенной рекомбинации. I. Область RII бактериофага T4. (1962) Журнал теоретической биологии. 1962; 3, 335-353. https://doi.org/10.1016/S0022-5193(62)80030-7
- ^ Jump up to: а беременный Бернштейн Х. Ремонт и рекомбинация в фаге T4. I. Гены, влияющие на рекомбинацию. Холодный весенний симп Quant Biol. 1968; 33: 325-331. doi: 10.1101/sqb.1968.033.01.037
- ^ Hamlett NV, Berger H. Мутации, изменяющие генетическую рекомбинацию и восстановление ДНК в бактериофаге T4. Вирусология. 1975; 63 (2): 539-567. doi: 10.1016/0042-6822 (75) 90326-8
- ^ Fujisawa H, Yonesaki T, Minagawa T. Последовательность гена рекомбинации T4, UVSX и его сравнение с последованием гена Reca Escherichia coli. Нуклеиновые кислоты Res. 1985; 13 (20): 7473-7481. doi: 10.1093/nar/13.20.7473
- Гриффитс AJF; Миллер JH; Suzuki dt; Lewontin RC; и др. (1993). «Глава 5» . Введение в генетический анализ (5 -е изд.). Нью -Йорк: WH Freeman and Company. ISBN 978-0-7167-2285-4 .
- Poehlman JM; Sleeper DA (1995). «Глава 3». Полевые культуры размножения (4 -е изд.). Айова: штат Айова Пресс. ISBN 978-0-8138-2427-7 .
| Базы данных управления авторитетом : национальный |
|---|