Факультативный анаэробный организм
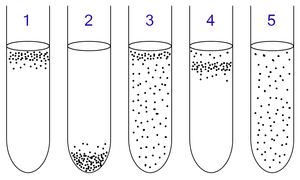
1: Обязательные аэробузы нуждаются в кислороде, потому что они не могут ферментировать или дышать анаэробно . Они собираются в верхней части трубки, где концентрация кислорода является самой высокой.
2: Обязательные анаээробиты отравлены кислородом, поэтому они собираются на дне трубки, где концентрация кислорода является самой низкой.
3: факультативные анаээробиты могут расти с кислородом или без него, потому что они могут метаболизировать энергию аэробно или анаэробно. Они собираются в основном на вершине, потому что аэробное дыхание генерирует больше АТФ, чем ферментация.
4: Микроаэрофилы нуждаются в кислороде, потому что они не могут ферментировать или дышать анаэробно. Тем не менее, они отравлены высокими концентрациями кислорода. Они собираются в верхней части пробирки, но не самая вершина.
5: Аэротолерантные анаэробные анаэробные. Не требуют кислорода, поскольку они используют ферментацию для изготовления АТФ. В отличие от облигатных анаэробных, они не отравлены кислородом. Их можно найти равномерно распространяться по всей пробирке.
Факультативный анаэробный организм - это организм , который делает АТФ путем аэробного дыхания, если присутствует кислород , но способен переключаться на ферментацию , если кислород отсутствует. [ 1 ] [ 2 ]
Некоторые примеры факультативно анаэробных бактерий являются Staphylococcus spp. В [ 3 ] Escherichia coli , Salmonella , Listeria spp., [ 4 ] Shewanella onidensis и Yersinia pestis . Определенные эукариоты также являются факультативными анаэробными анаэробными группами, включая грибы , такие как Saccharomyces cerevisiae [ 5 ] и многие водные беспозвоночные, такие как нереидные полихеты . [ 6 ]
Было отмечено, что у мутантов сальмонеллы тифимурий , который подвергался мутациям, как облигатные аэробузы, либо анаэробные, существовали различные уровни белков-хроматина. Позже было обнаружено, что облигационные аэроботы имеют дефектную субъединицу ДНК -гиразы A ( Gyra ), в то время как облигатные анаэробные были дефектными в топоизомеразе I ( Topi ). Это указывает на то, что топоизомераза I и связанная с ней расслабление хромосомной ДНК необходимы для транскрипции генов, необходимых для аэробного роста, в то время как обратное верно для ДНК -гиразы. [ 7 ] Кроме того, в Escherichia coli K-12 было отмечено, что фосфофруктокиназа (PFK) существует в виде димера в аэробных условиях и в качестве тетрамера в анаэробных условиях. Учитывая роль PFK в гликолизе, это имеет значение для влияния кислорода на метаболизм глюкозы E. coli K-12 в отношении механизма эффекта Пастера . [ 8 ] [ 9 ]
Существует основная сеть транскрипционных факторов (TFS), которая включает в себя основную кислород-чувствительную ARCA и FNR, контролирующие адаптацию Escherichia coli к изменениям в доступности кислорода. Активность этих двух регуляторов свидетельствует о пространственных эффектах, которые могут влиять на экспрессию генов в микроаэробном диапазоне. Также наблюдалось, что эти чувствительные к кислороду белки защищены в цитоплазме потребителями кислорода в клеточной мембране, известной как терминальные оксидазы . [ 10 ]
Функции
[ редактировать ]Факультативные анаээробиты способны расти как в присутствии, так и в отсутствие кислорода из -за экспрессии как аэробных, так и анаэробных дыхательных цепей с использованием либо кислорода, либо альтернативного акцептора электронов. [ 11 ] Например, в отсутствие кислорода E. coli может использовать фумарат , нитрат , нитрит , диметилсульфоксид или оксид триметиламина в качестве акцептора электронов. [ 11 ] Эта гибкость позволяет факультативным анаээробам выжить в ряде среды и в средах с часто меняющимися условиями. [ 1 ]
Несколько видов протистов используют факультативный анаэробный метаболизм для усиления продукции АТФ, а некоторые могут производить дигидроген в рамках этого процесса. [ 12 ]
Как патогены
[ редактировать ]Поскольку факультативные анаээробиты способны расти как в присутствии, так и в отсутствии кислорода, они могут выжить во многих различных средах, легко адаптироваться к изменяющимся условиям и, таким образом, иметь избирательное преимущество перед другими бактериями. В результате большинство опасных для жизни патогенных микроорганизмов являются факультативными анаээробами. [ 1 ]
Способность факультативных анаэробных патогенов выжить без кислорода важна, поскольку их инфекция снижает уровень кислорода в кишечной ткани их хозяина. [ 13 ] Более того, способность факультативных анаэробных ограничений на уровне кислорода в местах инфекции полезна для них и других бактерий, поскольку диоксидген может образовывать реактивные формы кислорода (АФК). Эти виды токсичны для бактерий и могут повредить их ДНК, среди других компонентов. [ 1 ]



Смотрите также
[ редактировать ]- Аэробное дыхание
- Анаэробное дыхание
- Ферментация
- Обязанный аэробный
- Обязанный анаэробный
- Микроаэрофил
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Андре, Антонин С.; Дебанд, Лорина; Мартин, Бенуа С. (август 2021 г.). «Селективное преимущество факультативных анаэробных изделий зависит от их уникальной способности справляться с изменением уровня кислорода во время инфекции» . Клеточная микробиология . 23 (8): E13338. doi : 10.1111/cmi.13338 . ISSN 1462-5814 . PMID 33813807 . S2CID 233027658 .
- ^ Müller, Volker (2001-04-19). "Ферментационная бактерия " эл два : 10,1038/npg . ISBN 9780470016176 .
- ^ Райан К.Дж; Рэй CG, ред. (2004). Шеррис Медицинская микробиология (4 -е изд.). МакГроу Хилл. С. 261–271, 273–296. ISBN 0-8385-8529-9 .
- ^ Singleton P (1999). Бактерии в биологии, биотехнологии и медицине (5 -е изд.). Уайли. С. 444–454. ISBN 0-471-98880-4 .
- ^ Carlile MJ, Watkinson SC, Gooday GW (2001). Грибы (2 -е изд.). Академическая пресса. С. 85–105. ISBN 0-12-738446-4 .
- ^ Schöttler, U. (30 ноября 1979 г.). «Об анаэробном метаболизме трех видов Nereis (Annelida)» (PDF) . Серия прогресса в морской экологии . 1 : 249–54. Bibcode : 1979meps .... 1..249s . doi : 10.3354/meps001249 . ISSN 1616-1599 . Получено 14 февраля 2010 года .
- ^ Yamamoto, N. & Droffner, ML (1985). Механизмы, определяющие аэробный или анаэробный рост в факультативном анаэробном сальмонелле. Материалы Национальной академии наук , 82 (7), 2077-2081. https://doi.org/10.1073/pnas.82.7.2077
- ^ Doelle, HW (1974). Димерная и тетрамерная фосфофтрукокиназа и эффект пастера в Eschericia coli K-1 Письмо FEBS , 49 (2), 220-2 PII: 0014-5793 (74) 80516-8 ( core.ac.uk )
- ^ Pasteur L (1857). «Мемуары о ферментации лактического приступа» [диссертация на ферментацию, называемую лактической]. Отчеты Академии наук (по -французски). 45 (913–916): 1032–1036.
- ^ Rolfe, MD, Ocone, A., Stapleton, MR, Hall, S., Trotter, EW, Poole, RK, ... & Green, J. (2012). Системный анализ активности фактора транскрипции в средах со стабильными и динамическими концентрациями кислорода. Открытая биология , 2 (7), 120091. Https://doi.org/10.1098/rsob.120091
- ^ Jump up to: а беременный Unden, Готфрид; Trageser, Martin (1991). «Экспрессия генов, регулируемая кислородом в Escherichia coli: контроль анаэробного дыхания белком FNR» . Антони Ван Леувенхук . 59 (2): 65–76. doi : 10.1007/bf00445650 . ISSN 0003-6072 . PMID 1854188 . S2CID 33083164 .
- ^ Джинджер, Майкл Л.; Фриц-Лейлин, Лилиан К.; Фултон, Чендлер; Cande, W. Zacheus; Доусон, Скотт С. (2010-12-01). «Международный метаболизм в протистах: основанный на последовательности взгляд на факультативный анаэробный метаболизм у эволюционно разнообразных эукариот» . Профист . 161 (5): 642–671. doi : 10.1016/j.protis.2010.09.001 . ISSN 1434-4610 . PMC 3021972 . PMID 21036663 .
- ^ Дженневейн, Джонас; Матушак, Жасмин; Уолтер, Стеффи; Фелми, Боас; Гендера, Катрин; Шац, Валентин; Новоттни, Моника; Либш, Грегор; Хенсель, Майкл; Хардт, Вольф-Диетрих; Герлах, Роман Г.; Джейнч, Джонатан (декабрь 2015 г.). «Низко -кислородная напряженность, обнаруженная в S Almonella -инфицированной репликации кишечной ткани S, в макрофагах путем нарушения антимикробной активности и увеличивая вирулентность S -almonella: гипоксия и репликация сальмонеллы» . Клеточная микробиология . 17 (12): 1833–1847. doi : 10.1111/cmi.12476 . PMID 26104016 .
Внешние ссылки
[ редактировать ]- Факультативные анаэробные бактерии
- Обязательные анаэробные бактерии
- Анаэробные бактерии и анаэробные бактерии в разложении (стабилизация) органического вещества. Архивированный 2009-02-05 на машине Wayback
| Медицинский Микробиология | |||||||
|---|---|---|---|---|---|---|---|
| Биохимия и экология |
| ||||||
| Форма | |||||||
| Структура |
| ||||||
| Таксономия и эволюция |
| ||||||