Пелоневст
| Пелоневст Временной диапазон: средняя юра ,
| |
|---|---|

| |
| Скелет, Музей палеонтологии, Тюбинген. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Суперзаказ: | † Зауроптеригия |
| Заказ: | † Плезиозаврия |
| Семья: | † Плиозавриды |
| Род: | † Пелоневст Лидеккер , 1889 г. |
| Разновидность: | † П. филаркус
|
| Биномиальное имя | |
| † Пелонеуст филарх Сили , 1869 г.
| |
| Синонимы | |
| |
Peloneustes (что означает « пловец в грязи » ) — род плиозаврид - плезиозавров из средней юры Англии. Его останки известны из Питерборо, пачки Оксфордской глиняной формации которой относится к келловею , возраст . Первоначально он был описан как вид плезиозавра в 1869 году, а палеонтологом Гарри Говьером Сили в 1889 году выделил ему собственный род. затем натуралист Ричард Лидеккер Хотя многие виды были отнесены к Peloneustes , P. philarchus в настоящее время является единственным, который все еще считается действительным. , а остальные перешли в другие роды, считающиеся nomina dubia или синонимами P. philarchus . Часть материала, ранее отнесенного к P. evansi, с тех пор была передана « Pliosaurus » andrewsi . Peloneustes известен по многим экземплярам, включая очень полный материал.
При общей длине 3,5–4 метра (11–13 футов) Peloneustes не является крупным плиозавридом. У него был большой треугольный череп, занимавший примерно пятую часть длины его тела. Передняя часть черепа вытянута в узкий рострум (рыло). Нижнечелюстной симфиз , где передние концы каждой стороны нижней челюсти удлинен сливаются, у Peloneustes и помогает укрепить челюсть. Между зубными рядами на нижнечелюстном симфизе расположен приподнятый гребень. Зубы Peloneustes имеют коническую форму, круглое поперечное сечение и несут вертикальные гребни со всех сторон. Передние зубы крупнее задних. Имея всего от 19 до 21 шейного позвонка , у Пелоневста была короткая шея для плезиозавра. Конечности Пелоневста превратились в ласты, причем задняя пара была больше передней.
Peloneustes интерпретировался либо как близкий родственник Pliosaurus , либо как более базальный (раннерасходящийся) плиозаврид внутри Thalassophonea , причем последняя интерпретация нашла большую поддержку. Как и другие плезиозавры, Пелоневст был хорошо приспособлен к водной жизни, используя свои ласты для метода плавания, известного как подводный полет . Черепа плиозавридов были усилены, чтобы лучше противостоять стрессам, связанным с кормлением. Длинная и узкая морда Пелоневста могла быстро махать водой, чтобы поймать рыбу, которую он протыкал своими многочисленными острыми зубами. Пелоневст обитал в эпиконтинентальном (внутреннем) море глубиной около 30–50 метров (100–160 футов). Он разделял свою среду обитания с множеством других животных, включая беспозвоночных , рыб, талаттозухов , ихтиозавров и других плезиозавров. По крайней мере, пять других плиозавридов известны из Питерборо, но их анатомия весьма различалась, что указывает на то, что они питались разными источниками пищи, тем самым избегая конкуренции.
История исследований
[ редактировать ]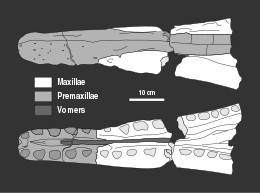
Пласты Оксфордской Питерборо, члена глиняной формации, уже давно разрабатываются для изготовления кирпича . С конца XIX века, когда начались эти работы, окаменелости многих морских животных. из скал были извлечены [ 2 ] Среди них был экземпляр, который впоследствии стал голотипом Peloneustes philarchus , обнаруженный геологом Генри Портером в глиняной яме недалеко от Питерборо , Англия. Образец включает нижнюю челюсть , переднюю часть верхней челюсти, различные позвонки по всему телу, элементы плечевого пояса и таза , плечевые кости (кости плеча), бедра (кости верхней части голени) и различные другие кости конечностей. [ 3 ] В 1866 году геолог Адам Седжвик приобрел образец для Кембриджского университета ( Вудвордского музея ныне Седжвикский музей наук о Земле, Кембридж). [ 2 ] образец занесен в каталог как CAMSM J.46913 и хранится в лекционной аудитории университета в кабинете D. [ 2 ] [ 3 ] Палеонтолог Гарри Говьер Сили описал этот экземпляр как новый вид ранее существовавшего рода Plesiosaurus , Plesiosaurus philarchus , в 1869 году. [ 3 ] Видовое имя означает « властолюбивый » , возможно, из-за его большого и мощного черепа. [ 4 ] Сили не описывал этот экземпляр подробно, а лишь перечислил известный материал. [ 3 ] Хотя в более поздних публикациях эти останки будут описаны более подробно, CAMSM J.46913 остается плохо описанным. [ 2 ]
Альфред Лидс и его брат Чарльз Лидс собирали окаменелости из Оксфордской глины примерно с 1867 года при поддержке геолога Джона Филлипса из Оксфордского университета , собрав то, что стало известно как Коллекция Лидса. Хотя Чарльз в конце концов ушел, Альфред, собравший большую часть образцов, продолжал собирать окаменелости до 1917 года. В конце концов, после посещения Генри Вудвордом Британского музея естественной истории (ныне Музей естественной истории в Лондоне) коллекции Лидса. В Айбери в 1885 году музей купил около 5 тонн (5,5 коротких тонн) окаменелостей в 1890 году. Это принесло коллекции Лидса более широкую известность, и позже он продал образцы музеи по всей Европе и даже некоторые в США. [ 5 ] Тщательно подготовленный материал обычно находился в хорошем состоянии, хотя нередко он был раздроблен и разрушен геологическими процессами. Черепа были особенно уязвимы для этого. [ 6 ] : v – vi

Натуралисту Ричарду Лидеккеру о скелете плезиозавра в Британском музее естественной истории сообщил геолог Джордж Чарльз Крик работавший там . Образец, внесенный в каталог под номером NHMUK PV R1253, [ 2 ] был обнаружен в формации Оксфорд-Клей в Грин-Энде , Кемпстон, недалеко от Бедфорда . Хотя Лидеккер предположил, что когда-то скелет был целым, он был поврежден во время раскопок. Когда экземпляр прибыл в музей, пояса конечностей были сильно фрагментированы, но работнику по имени Лингард из отдела геологии удалось восстановить большую часть из них. Помимо поясов конечностей, экземпляр также состоит из части нижней челюсти, зубов, нескольких позвонков (но не шейных) и большей части конечностей. Лидеккер идентифицировал этот экземпляр как особь Plesiosaurus philarchus и опубликовал его описание в 1889 году. Изучив этот и другие экземпляры из коллекции Лидса, он пришел к выводу, что плезиозавры с укороченной шеей и большими головами не могут быть классифицированы как виды Plesiosaurus , т.е. что "П." philarchus принадлежал к другому роду. Первоначально он приписал его Тауматозавру в 1888 году. [ 7 ] но позже решил, что он достаточно отличен, чтобы выделить собственный род, который он назвал Peloneustes в своей публикации 1889 года. [ 8 ] Название Пелонеустес происходит от греческих слов pelos , что означает « ил » или « глина » , в отношении формации Оксфорд-Клей, и neustes , что означает « пловец » . [ 4 ] Сили, однако, в 1892 году объединил Пелоневста с Плиозавром , заявив, что эти два вида недостаточно различаются, чтобы их можно было выделить в отдельные роды. [ 9 ] Сили и Лидеккер не смогли прийти к единому мнению, к какому роду отнести P. philarchus , что является частью вражды между двумя учеными. Однако с тех пор имя Пелоневст стало общепринятым. [ 7 ]
Коллекция Лидса содержала несколько экземпляров Peloneustes . [ 10 ] : 63–70 В 1895 году палеонтолог Чарльз Уильям Эндрюс описал анатомию черепа Пелоневста на основе четырех частичных черепов из коллекции Лидса. [ 11 ] В 1907 году геолог Фредерик Жаккар опубликовал описание двух экземпляров Peloneustes из Оксфордской глины недалеко от Питерборо, хранившихся в Палеонтологическом музее Лозанны, Швейцария . Более полный из двух экземпляров включает полный череп с сохранением обеих челюстей; множественные изолированные зубы; 13 шейных (шейных), 5 грудных (плечевых) и 7 хвостовых (хвостовых) позвонков; ребра; обе лопатки, клювовидный отросток ; частичный межключичный; полный таз, за исключением седалищной кости ; и все четыре конечности, которые были почти целы. У другого экземпляра сохранилось 33 позвонка и несколько ребер. Поскольку экземпляр, описанный Лидеккером, нуждался в некоторой реставрации, а недостающая информация была восполнена данными из других образцов в его публикации, Жаккар счел уместным опубликовать описание, содержащее фотографии более полного экземпляра из Лозанны, чтобы лучше проиллюстрировать анатомию Пелоневст . [ 12 ]
В 1913 году натуралист Герман Линдер описал несколько экземпляров Peloneustes philarchus, хранившихся в Институте геологических исследований Тюбингенского университета и Государственном музее естественной истории Штутгарта , Германия. Эти экземпляры также были из коллекции Лидса. [ 2 ] Среди экземпляров, которые он описал из бывшего учреждения, был почти полный скелет, в котором отсутствовали два шейных позвонка, несколько хвостовых позвонков на конце хвоста и несколько дистальных фаланг. Только задняя часть черепа была в хорошем состоянии, а нижняя челюсть практически не повреждена. Еще одним экземпляром, описанным Линдером, был череп хорошей сохранности (GPIT RE/3409). [ 2 ] также из Тюбингенского университета, сохранив склеротическое кольцо (набор мелких костей, поддерживающих глаз), только в четвертый раз об этих костях сообщалось у плезиозавра. [ 13 ]


Позже Эндрюс описал образцы морских рептилий из коллекции Лидса, которые находились в Британском музее естественной истории, опубликовав два тома, один в 1910 году, а другой в 1913 году. Анатомия образцов пелонеустов была описана во втором томе, основанном главным образом на хорошо сохранившиеся черепа NHMUK R2679 и NHMUK R3808 и NHMUK R3318, почти полный скелет. NHMUK R3318 настолько хорошо сохранился, что его можно было пересобрать и установить, хотя недостающие части таза и конечностей пришлось заполнить. Установленный скелет был выставлен в Галерее ископаемых рептилий музея. [ 6 ] : IX [ 10 ] : 35, 63 Эндрюс описал это животное в 1910 году, отметив, что это был первый скелет плиозаврид , предоставив тем самым важную информацию об общей анатомии группы. [ 14 ]
В 1960 году палеонтолог Ламберт Беверли Тарло опубликовал обзор видов плиозавров, о которых сообщалось из верхней юры . Многие виды плиозавридов были названы на основе отдельных фрагментов, что создавало путаницу. Тарло также обнаружил, что неточные описания материала и игнорирование работ палеонтологами друг друга только усугубляли эту путаницу. Из 36 видов, которые он рассмотрел, он нашел только девять действительными, включая Peloneustes philarchus . [ 7 ] В 2011 году палеонтологи Хилари Кетчум и Роджер Бенсон описали анатомию черепа Пелоневста . Со времени предыдущих анатомических исследований Эндрюса и Линдера было обнаружено больше образцов, в том числе NHMUK R4058, череп, сохранившийся в трех измерениях, что дает лучшее представление о форме черепа. [ 2 ]
Другие назначенные виды
[ редактировать ]Многие другие виды были отнесены к Peloneustes на протяжении всей его таксономической истории, но с тех пор все они были отнесены к другим родам или признаны недействительными. [ 2 ] В той же публикации, в которой он назвал P. philarchus , Сили также назвал другой вид плезиозавра на , P. sterrodeirus, основе семи экземпляров в Вудвордском музее, состоящих из черепного и позвоночного материала. [ 3 ] Когда Лидеккер выделил род Peloneustes для P. philarchus , он также реклассифицировал «Plesiosaurus» sterrodeirus и « Pleiosaurus » aequalis (вид, названный Джоном Филлипсом в 1871 году). [ 15 ] : 365 [ 7 ] как представители этого рода. [ 8 ] плиозавридов 1960 года В своем обзоре таксономии Тарло считал P. aequalis недействительным, поскольку он основан на проподиалах (костях верхних конечностей), которые нельзя использовать для дифференциации разных видов плиозавридов. Он считал, что Peloneustes sterrodeirus вместо этого принадлежит Pliosaurus , возможно, в пределах P. brachydeirus . [ 7 ]

Еще одним видом, описанным Сили в 1869 году, был Pliosaurus evansi , основанный на экземплярах из Вудвордского музея. [ 3 ] Они состояли из шейных и спинных (спинных) позвонков , ребер и клювовидного отростка. Из-за того, что это был меньший вид плиозавра и его сходство с Peloneustes philarchus , Лидеккер в 1890 году отнес его к Peloneustes , отметив, что он был крупнее Peloneustes philarchus . [ 16 ] Он также считал, что большая нижняя челюсть и лопатка, приписываемые Pleiosaurus ?grandis. Филлипсом в 1871 году [ 15 ] : 318 вместо этого принадлежал к этому виду. [ 7 ] В 1913 году Эндрюс приписал частичный скелет другого крупного плиозавра, найденного Лидсом, к Peloneustes evansi , отметив, что, хотя нижняя челюсть и позвонки были похожи на другие экземпляры Peloneustes evansi , они сильно отличались от таковых у Peloneustes philarchus . Следовательно, Эндрюс считал возможным, что P. evansi действительно принадлежал к отдельному роду, морфологически занимавшему промежуточное положение между Peloneustes и Pliosaurus . [ 10 ] : 72 В своем обзоре плиозавридов 1960 года Тарло синонимизировал Peloneustes evansi с Peloneustes philarchus , поскольку их шейные позвонки идентичны (за исключением разницы в размере). Он считал более крупные экземпляры Peloneustes evansi отдельными и относил их к новому виду Pliosaurus , P. andrewsi (хотя этот вид больше не считается принадлежащим к Pliosaurus ). [ 7 ] [ 17 ] Хилари Ф. Кетчум и Роджер Б. Дж. Бенсон не согласились с этой синонимией и в 2011 году посчитали, что, поскольку голотип Peloneustes evansi не имеет диагностических свойств (отсутствует отличительные особенности), P. evansi является nomen dubium и, следовательно, неопределенным плиозавридом. [ 2 ]
Палеонтолог Э. Кокен описал в 1905 году другой вид Peloneustes , P. kanzleri , из мела Германии. [ 2 ] В 1960 году Тарло повторно идентифицировал этот вид как эласмозаврид . [ 7 ] В 1913 году Линдер создал подвид Peloneustes , P. philarchus var. spathyrhynchus , дифференцируя его по лопатчатому нижнечелюстному симфизу (где две стороны нижней челюсти встречаются и сливаются). [ 13 ] Тарло считал его синонимом Peloneustes philarchus в 1960 году. [ 7 ] а нижнечелюстной симфиз Peloneustes пропорционально шире у более крупных экземпляров, что делает этот признак более вероятным следствием внутривидовой изменчивости (изменений внутри вида). Дробление затрудняет точное измерение этих пропорций. [ 2 ] В 1948 году палеонтолог Нестор Новожилов назвал новый вид Peloneustes , P. irgisensis , на основе PIN 426, частичного скелета, состоящего из большого неполного черепа, позвонков и частичной задней конечности с сохранившимся содержимым желудка. Экземпляр был обнаружен в бассейне Нижней Волги в России. [ 18 ] [ 19 ] В своем обзоре 1960 года Тарло посчитал этот вид слишком отличным от Peloneustes philarchus, чтобы принадлежать к Peloneustes , предварительно отнеся его к Pliosaurus . Он предположил, что Новожилов ошибочно считал Пелоневста единственным длиннорылым плиозавридом, отсюда и первоначальное назначение. [ 7 ] В 1964 году Новожилов выделил для этого вида новый род — Strongylokrotaphus , но дальнейшие исследования согласились с мнением Тарло и отнесли этот вид к Pliosaurus , возможно, синониму Pliosaurus rossicus . К тому времени PIN 426 сильно пострадал от пирита . [ 19 ] [ 17 ]

В 1998 году палеонтолог Фрэнк Робин О'Киф предположил, что образец плиозаврид из нижнеюрских сланцев Посидония в Германии может представлять собой новый вид Peloneustes . В 2001 году он посчитал его принадлежащим к отдельному роду, назвав его Hauffiosaurus zanoni . [ 2 ] [ 20 ] Палеонтологи Зульма Гаспарини и Мануэль А. Итурральде-Винент отнесли плиозаврид из верхнеюрской формации Джагуа на Кубе к Peloneustes sp. в 2006 году. [ 21 ] В 2009 году Гаспарини переописал его как Gallardosaurus iturraldei . [ 22 ] В 2011 году Кетчум и Бенсон считали, что Пелонеустес содержит только один вид — P. philarchus . Они обнаружили двадцать один определенный экземпляр Peloneustes philarchus , все из пачки Питерборо формации Оксфордской глины. Они считали, что некоторые экземпляры из члена Питерборо и Маркизы , Франция, ранее отнесенные к Пелоневсту , принадлежали к разным, в настоящее время безымянным плиозавридам. [ 2 ]
Описание
[ редактировать ]
Пелоневст – небольшой [ 10 ] : 34 до среднего представителя Pliosauridae. [ 23 ] : 12 NHMUK R3318, установленный скелет в Музее естественной истории в Лондоне, имеет длину 3,5 метра (11,5 футов). [ 14 ] а длина установленного скелета в Институте геонаук Тюбингенского университета составляет 4,05 метра (13,3 фута). [ 13 ] Плезиозавров обычно можно охарактеризовать как представителей морфотипа «плезиозавроморф» с маленькой головой и длинной шеей или морфотипа «плиозавроморф» с большой головой и короткой шеей. [ 24 ] Peloneustes принадлежит к последнему морфотипу, [ 24 ] череп составляет немногим менее пятой части общей длины животного. [ 23 ] : 13 Пелонеуст , как и все плезиозавры, имел короткий хвост, массивное туловище, а все его конечности превратились в большие ласты. [ 23 ] : 3
Череп
[ редактировать ]
Хотя у голотипа Peloneustes отсутствует задняя часть черепа, к этому роду отнесено множество дополнительных хорошо сохранившихся экземпляров, в том числе тот, который не был раздавлен сверху вниз. Эти черепа различаются по размеру: длина их составляет 60–78,5 см (1,97–2,58 фута). Череп Пелоневста удлинен и наклонен вверх к заднему концу. [ 2 ] При осмотре сверху череп имеет форму равнобедренного треугольника . [ 10 ] : 35 задняя часть черепа широкая, а передняя часть вытянута в узкий рострум . Самая задняя часть черепа имеет примерно параллельные стороны, в отличие от сужающихся передних областей. Наружные ноздри (отверстия для ноздрей) маленькие и расположены примерно на середине длины черепа. Почковидные глазницы обращены вперед и наружу и расположены в задней половине черепа. Склеротические кольца состоят как минимум из 16 отдельных элементов, что является необычно большим количеством для рептилий. ( Височные окна отверстия в задней части черепа) увеличены, эллиптические и расположены в самой задней четверти черепа. [ 2 ]

Характерно, что предчелюстные кости (передние верхние зубные кости) Peloneustes несут по шесть зубов каждая, а диастемы (промежутки между зубами) верхней челюсти узкие. Хотя утверждалось, что у Пелоневста были носовые кости (кости, ограничивающие внешние ноздри), хорошо сохранившиеся экземпляры указывают на то, что это не так. кости Лобные (кости, граничащие с глазницами) Пелоневстеса соприкасаются как с глазницами, так и с внешними ноздрями, что является отличительной чертой Пелоневстеса . Были некоторые разногласия относительно того, ли у Пелоневста были слезные кости (кости, граничащие с нижними передними краями глазниц) из-за плохой сохранности. Однако хорошо сохранившиеся экземпляры указывают на то, что слезные кости представляют собой отдельные кости, как и у других плиозавридов, в отличие от расширений скуловых костей ( костей, граничащих с нижними задними краями глазниц). Нёбо . Peloneustes плоское и имеет множество отверстий, в том числе внутренние ноздри (отверстие носового хода в рот) Эти отверстия соприкасаются с небными костями, известными как palatines — конфигурация, используемая для идентификации этого рода. Парасфеноид (кость , образующая нижнюю переднюю часть черепной коробки ) несет длинный культообразный отросток (выступ черепной коробки вперед), который виден, если смотреть на небо снизу, что является еще одной отличительной особенностью Peloneustes . Затылок . (задняя часть черепа) Пелоневста открытый, с большими отверстиями [ 2 ]
Peloneustes известен по множеству нижних челюстей, некоторые из которых хорошо сохранились. Самая длинная из них составляет 87,7 сантиметра (2,88 фута). Нижнечелюстной симфиз удлинен, составляет около трети общей длины нижней челюсти. Позади симфиза две стороны нижней челюсти расходятся, а затем плавно изгибаются внутрь возле заднего конца. Каждая зубная кость (несущая зубы кость нижней челюсти) имеет от 36 до 44 зубов, от 13 до 15 из которых расположены на симфизе. со второго по седьмой Зубные лунки (зубные лунки) крупнее тех, что расположены дальше назад, а наиболее широк симфиз около пятого и шестого. В дополнение к характеристикам зубов нижней челюсти, Peloneustes также можно узнать по венечным отросткам (верхним внутренним нижнечелюстным костям), которые вносят вклад в нижнечелюстной симфиз. Между зубными рядами нижнечелюстной симфиз имеет приподнятый гребень в месте соприкосновения зубных костей. Это уникальная особенность Пелоневста , не встречающаяся ни у каких других плезиозавров. Нижнечелюстной сустав (гнездо челюстной сустав ) широкий, почковидной формы, загнут вверх и внутрь. [ 2 ]

Зубы Peloneustes имеют круглое поперечное сечение, как и у других плиозавридов того же возраста. [ 7 ] Зубы имеют форму загнутых конусов . Эмаль коронок со всех сторон имеет равномерно расположенные вертикальные гребни различной длины. Эти гребни более сконцентрированы на вогнутом крае зубов. Большинство гребней простираются на половину-две трети общей высоты коронки, и лишь немногие из них фактически достигают вершины зуба. [ 2 ] Зубной ряд пелонеуста гетеродонтный , то есть имеет зубы разной формы. Более крупные зубы имеют клыковидную форму и расположены в передней части челюстей, тогда как меньшие зубы более резко загнуты назад. [ 2 ] более толстый и расположен дальше назад. [ 25 ]
Посткраниальный скелет
[ редактировать ]
В 1913 году Эндрюс сообщил, что у Пелонеуста было от 21 до 22 шейных, от 2 до 3 грудных и около 20 спинных позвонков, при этом точное количество крестцовых (бедренных) и хвостовых позвонков неизвестно, судя по образцам из коллекции Лидса. [ 10 ] : 47, 52 Однако в том же году Линдер сообщил о 19 шейных, 5 грудных, 20 спинных, 2 крестцовых и, по крайней мере, 17 хвостовых позвонках у Peloneustes , основываясь на образце из Института геовиссеншафтена Тюбингенского университета. [ 13 ] [ 2 ] Первые два шейных позвонка — атлант и аксис — у взрослых срослись, но у молодых они присутствуют в виде нескольких несросшихся элементов. [ 10 ] : 47 Межцентральная часть (часть тела позвонка) оси имеет примерно прямоугольную форму и проходит под центром (телом позвонка) атланта. [ 2 ] Шейные позвонки несут высокие нервные отростки , сжатые из стороны в сторону. [ 2 ] [ 10 ] : 50 Длина шейных центров примерно вдвое короче ширины. Они имеют сильно вогнутые суставные поверхности с выступающим краем вокруг нижнего края позвонков, расположенных ближе к передней части ряда. Каждый шейный центр имеет сильный киль вдоль средней линии нижней стороны. [ 7 ] Большинство шейных ребер имеют две головки, разделенные выемкой. [ 10 ] : 53
Грудные позвонки несут сочленения соответствующих ребер частично как на центральных, так и на нервных дугах. За этими позвонками следуют спинные позвонки, которые более удлинены, чем шейные позвонки, и имеют более короткие нервные отростки. И крестцовые, и хвостовые позвонки имеют менее удлиненные центральные части, ширина которых превышает высоту. Многие ребра от бедра и основания хвоста имеют увеличенные внешние концы, которые, кажется, сочленяются друг с другом. В 1913 году Эндрюс предположил, что такая конфигурация могла бы усилить хвост, возможно, для поддержки больших задних конечностей. Конечные (последние) хвостовые позвонки резко уменьшаются в размерах и могли бы поддерживать пропорционально более крупные шевроны, чем хвостовые позвонки, расположенные дальше вперед. В 1913 году Эндрюс предположил, что эта морфология могла поддерживать небольшую структуру, похожую на хвостовой плавник . [ 10 ] : 52–53 Предполагается, что и у других плезиозавров были хвостовые плавники, отпечатки такой структуры, возможно, известны у одного вида. [ 26 ]
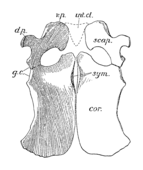

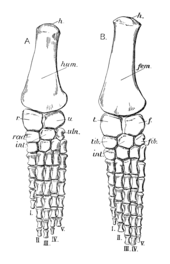
Плечевой пояс Пелоневста был большим, хотя и не таким массивным, как у некоторых других плезиозавров. Коракоиды — самые крупные кости плечевого пояса, имеют пластинчатую форму. Плечевой сустав образован как лопаткой (лысиной), так и клювовидным отростком, при этом две кости образуют угол 70° друг с другом. Лопатки типичны по форме для плиозаврид и трехлучевых, с тремя выступающими выступами или ветвями. Дорсальная (верхняя) ветвь направлена наружу, вверх и назад. [ 10 ] : 55 [ 7 ] На нижней стороне каждой лопатки имеется гребень, направленный к переднему краю ее вентральной (нижней) ветви. [ 7 ] Вентральные ветви двух лопаток были отделены друг от друга треугольной костью, известной как межключица . Как и у других плиозавров, таз Peloneustes имеет большие и плоские седалищные и лобковые кости . Третья тазовая кость, подвздошная , меньше по размеру и удлинена, сочленяется с седалищной костью. Верхний конец подвздошной кости демонстрирует большое количество вариаций внутри P. philarchus , известны две формы: одна с закругленным верхним краем, другая с плоским верхним краем и более угловатой формой. [ 10 ] : 55–56, 58–60
Задние конечности Пелоневста длиннее передних, при этом бедренная кость длиннее плечевой кости, хотя плечевая кость более крепкая из двух элементов. [ 10 ] : 57, 60 ( Лучевая кость одна из нижних костей передних конечностей) примерно равна ширине и длине, в отличие от локтевой кости (другой нижней кости передней конечности), ширина которой превышает длину. [ 7 ] Радиус — больший из этих двух элементов. [ 10 ] : 58 Большеберцовая кость больше малоберцовой кости (нижние кости задних конечностей) и длиннее ширины, а у некоторых экземпляров ширина малоберцовой кости превышает длину. [ 7 ] , Пястные кости плюсневые кости и проксимальные фаланги пальцев (некоторые кости, составляющие внешнюю часть лопатки) уплощены. Большинство фаланг обеих конечностей имеют округлое поперечное сечение, и все они имеют заметные перетяжки посередине. Количество фаланг на каждом пальце неизвестно как на передних, так и на задних конечностях. [ 10 ] : 58, 62
Классификация
[ редактировать ]
Первоначально Сили описал Peloneustes как вид Plesiosaurus , что было довольно распространенной практикой (в то время объем родов был аналогичен тому, который сейчас используется для семейств ). [ 23 ] : 7 В 1874 году Сили назвал новое семейство плезиозавров Pliosauridae, содержащее формы, похожие на Pliosaurus . [ 27 ] В 1890 году Лидеккер отнес Пелоневста . к этой семье [ 16 ] которому с тех пор он последовательно присваивался. [ 9 ] [ 10 ] : 1 [ 7 ] [ 2 ] Точно неизвестно, как плиозавриды связаны с другими плезиозаврами. В 1940 году палеонтолог Теодор Э. Уайт считал плиозавридов близкими родственниками Elasmosauridae на основании анатомии плеча. [ 28 ] Палеонтолог Сэмюэл П. Уэллс , однако, считал, что плиозавриды были более похожи на Polycotylidae , поскольку у них обоих были большие черепа и короткие шеи, среди других характеристик. Он сгруппировал эти два семейства в надсемейство Pliosauroidea , а другие плезиозавры образовали надсемейство Plesiosauroidea . [ 29 ] Другое семейство плезиозавров, Rhomaleosauridae , с тех пор было отнесено к Pliosauroidea. [ 30 ] [ 20 ] а Polycotylidae были присвоены Plesiosauroidea. [ 31 ] [ 32 ] Однако в 2012 году Бенсон и его коллеги обнаружили другую топологию: Pliosauridae были более тесно связаны с Plesiosauroidea, чем с Rhomaleosauridae. Эта клада плиозавридов-плезиозавроидов получила название Neoplesiosauria . [ 32 ]

Внутри Pliosauridae точное филогенетическое положение Peloneustes неясно. [ 2 ] В 1889 году Лидеккер считал Пелонеуста переходной формой между плиозавром и более ранними плезиозаврами, хотя он нашел маловероятным, что Пелонеуст был предком плиозавра . [ 8 ] В 1960 году Тарло считал Peloneustes близким родственником Pliosaurus , поскольку оба таксона имели удлиненные нижнечелюстные симфизы. [ 7 ] В 2001 году О'Киф обнаружил его как базального (раннерасходящегося) члена этого семейства, вне группы, включающей лиоплевродона , плиозавра и брахаучениуса . [ 20 ] [ 2 ] Однако в 2008 году палеонтологи Адам С. Смит и Гарет Дж. Дайк обнаружили, Peloneustes является сестринским таксоном Pliosaurus что . [ 30 ] [ 2 ] В 2013 году Бенсон и палеонтолог Патрик С. Дракенмиллер назвали новую кладу внутри Pliosauridae — Thalassophonea . В эту кладу вошли «классические» плиозавриды с короткой шеей, но исключались более ранние, более изящные формы с длинной шеей. Пелоневст оказался самым базальным талассофоном. [ 33 ] Последующие исследования выявили аналогичное положение и у Пелоневста . [ 34 ] [ 35 ] [ 36 ]
Следующая кладограмма соответствует Кетчуму и Бенсону, 2022 г. [ 37 ]
Палеобиология
[ редактировать ]
Плезиозавры были хорошо приспособлены к морской жизни. [ 38 ] [ 39 ] [ 40 ] Они росли со скоростью, сравнимой со скоростью птиц, и имели высокий метаболизм , что указывает на гомеотермию. [ 41 ] или даже эндотермия . [ 40 ] Костный лабиринт , полость внутри черепа, в которой находился орган чувств, отвечающий за равновесие и ориентацию, у Пелоневста и других плезиозавров, по форме похож на лабиринт морских черепах . В 2017 году палеонтолог Джеймс Нинан и его коллеги выдвинули гипотезу, что эта форма, вероятно, развилась вместе с взмахивающими движениями, используемыми плезиозаврами для плавания. У пелонеуста и других плезиозавров с короткой шеей лабиринты были меньше, чем у плезиозавров с более длинной шеей, такая же закономерность наблюдается и у китообразных . [ 38 ] Кроме того, у Пелоневста , вероятно, были солевые железы в голове, чтобы справляться с избыточным количеством соли в теле. Тем не менее, пелонеуст , по-видимому, был хищником позвоночных животных , которые содержат меньше соли, чем беспозвоночные , поэтому ведущий палеонтолог Лесли Ноэ предположил в диссертации 2001 года, что эти железы не должны были быть особенно большими. [ 42 ] : 257 У Peloneustes , как и у многих других плиозавров, был снижен уровень окостенения костей. Палеонтолог Артур Круикшанк и его коллеги в 1966 году предположили, что это могло помочь Пелоневсте сохранить плавучесть или улучшить его маневренность. [ 43 ] Исследование, проведенное палеонтологом Коринной Флейшле и ее коллегами в 2019 году, показало, что у плезиозавров были увеличенные эритроциты , исходя из морфологии их сосудистых каналов , что помогло бы им во время ныряния. [ 39 ]
Плезиозавры, такие как Пелоневст, использовали метод плавания, известный как подводный полет , используя свои ласты в качестве подводных крыльев . Плезиозавры необычны среди морских рептилий тем, что для движения они использовали все четыре конечности, а не движения позвоночного столба. Короткий хвост, хотя вряд ли использовался для передвижения животного, мог помочь стабилизировать или направить плезиозавра. [ 44 ] [ 26 ] Передние ласты Пелоневста имеют соотношение сторон 6,36, а задние ласты — 8,32. Эти соотношения аналогичны соотношениям крыльев современных соколов . В 2001 году О'Киф предположил, что, как и соколы, плиозавроморфные плезиозавры, такие как Пелоневст, вероятно, были способны быстро и ловко передвигаться, хотя и неэффективно, для захвата добычи. [ 24 ] Компьютерное моделирование, проведенное палеонтологом Сусаной Гутаррой и ее коллегами в 2022 году, показало, что из-за своих больших ласт плезиозавр создавал бы большее сопротивление , чем китообразное или ихтиозавр сопоставимого размера . Однако плезиозавры противодействовали этому своим большим хоботом и размером тела. [ 45 ] Из-за уменьшения сопротивления из-за их более коротких и глубоких тел палеонтолог Джуди Массаре в 1988 году предположила, что плезиозавры могут активно искать и преследовать свою пищу, вместо того, чтобы ждать ее в ожидании. [ 44 ]
Механика кормления
[ редактировать ]
В диссертации 2001 года Ноэ отметил множество адаптаций черепов плиозавридов к хищничеству. Чтобы избежать повреждений во время кормления, черепа плиозавридов, таких как Peloneustes, имеют высокую акинетичность: кости черепа и нижней челюсти в основном зафиксированы на месте, чтобы предотвратить движение. Морда содержит удлиненные кости, которые помогают предотвратить изгиб, и имеет усиленное соединение с лицевой областью, чтобы лучше противостоять нагрузкам во время кормления. При взгляде сбоку видно небольшое сужение нижней челюсти, укрепляющее ее. Нижнечелюстной симфиз помог бы обеспечить равномерный прикус и предотвратить независимое движение нижних челюстей. Увеличенное венечное возвышение обеспечивает большую и прочную область для крепления челюстных мышц, хотя эта структура не такая большая у Peloneustes , как у других современных плиозавридов. Области, где закреплялись мышцы челюсти, расположены дальше от черепа, чтобы не мешать кормлению. Гленоид нижней челюсти в форме почки сделал бы челюстной сустав более устойчивым и предотвратил бы вывих нижней челюсти. Зубы плиозаврид прочно укоренены и сцеплены между собой, что укрепляет края челюстей. Эта конфигурация также хорошо сочетается с простыми вращательными движениями, которыми были ограничены челюсти плиозаврид, и укрепляет зубы, защищая их от борьбы с добычей. Более крупные передние зубы использовались, чтобы пронзить добычу, в то время как меньшие задние зубы раздавливали и направляли добычу назад к горлу. Благодаря своим широким зевам плиозавриды не особо перерабатывали пищу перед проглатыванием. [ 42 ] : 193, 236–240

Многочисленные зубы Пелоневста редко ломаются, но на их кончиках часто имеются следы износа. Их острые кончики, слегка изогнутая, изящная форма и заметное расстояние между ними указывают на то, что они были созданы для пирсинга. Тонкая, вытянутая морда по форме похожа на дельфинью. Морфология морды и зубов позволила Ноэ предположить, что Пелоневст был рыбоядным животным (рыбоедом). Чтобы поймать добычу, Пелоневст быстро повернул голову в сторону. Примерно круглое поперечное сечение изящной морды сводило сопротивление к минимуму, а длинные челюсти подходили для быстрого захвата подвижной добычи. Плоское, увеличенное небо и усиленный мозговой череп Пелоневста уменьшили бы скручивание, изгиб и сдвиг, вызванные длинной мордой. Усиленная мозговая оболочка уменьшила бы поглощение ударов в области мозга. Поскольку кончик морды находился дальше от челюстного сустава, он оказывал меньшую силу укуса, чем участки, расположенные дальше назад. Передние отделы челюстей Пелоневсты имеют удлиненную форму, что указывает на то, что их использовали для быстрого нападения и задержания добычи. Эти адаптации указывают на предпочтение более мелкой добычи, которую, несмотря на ее подвижность, было бы легче вывести из строя и она была менее мощной. Однако Пелоневст все равно был способен атаковать рыбу среднего размера. В то время как скелет, занесенный в каталог как NHMUK R3317, с остатками белемнита приписал Пелоневсту , в желудке, Эндрюс в 1910 году [ 6 ] : xvi – xvii он очень неполный и на самом деле может принадлежать современным плиозавридам Simolestes , как предполагает Ноэ. [ 42 ] : 233–234, 241–242 Пелоневст Брюс Ротшильд и его коллеги также предположили, что оставил следы укусов на экземпляре Cryptoclidus в 2018 году. [ 46 ]
Палеоэкология
[ редактировать ]Палеосреда
[ редактировать ]
Пелонеустес известен из пачки Питерборо (ранее известной как Нижняя Оксфордская глина) формации Оксфордской глины. [ 2 ] Хотя Пелоневст считается выходцем из оксфордского периода (примерно от 164 до 157 миллионов лет назад). [ 1 ] ) верхней юры, [ 7 ] член Питерборо на самом деле относится к келловейскому этапу (примерно от 166 до 164 миллионов лет назад). [ 1 ] ) средней юры . [ 2 ] Пачка Питерборо простирается от позднего нижнего келловея до раннего верхнего келловея, занимая весь средний келловей. [ 47 ] Он перекрывает формацию Келлауэйс. [ 47 ] и перекрыт пачкой Стюартби формации Оксфорд-Клей. [ 48 ] Член Питерборо в основном состоит из серого битума (содержащего асфальт). [ 47 ] сланец и глина [ 49 ] богат органическими веществами . [ 50 ] Эти породы иногда бывают делящимися (раскалываются на тонкие плоские плиты). [ 48 ] Толщина этого члена составляет около 16–25 метров (52–82 футов) и простирается от Дорсета до Хамбера . [ 47 ]
Член Питерборо представляет собой эпиконтинентальное море во время повышения уровня моря . [ 50 ] На момент отложения он должен был располагаться на 35° северной широты . [ 48 ] Это море, известное как море Оксфордской глины, было в значительной степени окружено островами и континентами, которые обеспечивали морской путь отложениями . [ 48 ] Его близость к суше подтверждается сохранением наземных окаменелостей, таких как коряги в Оксфордской глине, а также обломочной дайки на нижних уровнях члена Питерборо, причем образованию дайки способствовала дождевая вода . [ 49 ] Южный регион Оксфордского глиняного моря был соединен с океаном Тетис , а на его северной стороне — с более бореальными регионами. Это позволило осуществить обмен фауной между Тетическим и бореальным регионами. Это море имело глубину примерно 30–50 метров (100–160 футов) в пределах 150 километров (93 миль) от береговой линии. [ 48 ] [ 2 ]

Окружающая земля имела средиземноморский климат с сухим летом и влажной зимой, хотя она становилась все более засушливой . На основании информации δ 18 Изотопы O в двустворчатых моллюсках , температура воды на морском дне члена Питерборо варьировалась от 14–17 ° C (57–63 ° F) из-за сезонных колебаний, со средней температурой 15 ° C (59 ° F). Окаменелости белемнита дают аналогичные результаты, давая диапазон температуры воды от минимум 11 ° C (52 ° F) до максимума от 14 ° C (57 ° F) или 16 ° C (61 ° F), со средней температурой 13 ° C. °С (55 °Ф). [ 48 ] В то время как следы зеленых серных бактерий указывают на эвксиновую воду с низким содержанием кислорода и высоким уровнем сероводорода , обильные следы бентосных (донных) организмов позволяют предположить, что придонные воды не были бескислородными . [ 51 ] [ 50 ] Уровни кислорода, по-видимому, различались: некоторые отложения отлагались в более аэрированных условиях, чем другие. [ 48 ]
Современная биота
[ редактировать ]В членстве Питерборо сохранилось много видов беспозвоночных. К ним относятся головоногие моллюски , к которым относятся аммониты , белемниты и наутилоиды . Двустворчатые моллюски — еще одна многочисленная группа, тогда как брюхоногие моллюски и кольчатые черви менее распространены, но все же довольно распространены. Членистоногие также присутствуют. Брахиоподы и иглокожие встречаются редко. не известны по окаменелостям, Несмотря на то, что полихеты они, вероятно, присутствовали в этой экосистеме из-за их обилия в аналогичных современных средах и норах, подобных тем, которые производят эти черви. Микрофоссилии, относящиеся к фораминиферам , кокколитофороидам и динофлагеллятам , широко распространены в пачке Питерборо. [ 52 ]
В членстве Питерборо известно большое разнообразие рыб. К ним относятся хондрихтианы Asteracanthus , Brachymylus , Heterodontus (или Paracestracion ), [ 52 ] Гибодус , Исхиодус , Палеобрахаэлур , Пахимил , Протоспинакс , Лептакант , Нотидан , Оректолобоидес , Спатобатис и Сфенодус . Присутствовали также актиноптеригии , представленные Aspidorhynchus , Asthenocormus , Caturus , Coccolepis , Heterostrophus , Hypsocormus , Leedsichthys , Lepidotes , Leptolepis , Mesturus , Osteorachis , Pachycormus , Pholidophorus и Sauropsis . [ 53 ] Среди этих рыб есть поверхностно обитающие, средневодные и донные виды различных размеров, некоторые из которых могут достигать довольно крупных размеров. Они заполнили множество ниш, в том числе пожирателей беспозвоночных, рыбоядных животных и, в случае Leedsichthys , гигантских фильтраторов . [ 52 ]

Плезиозавры распространены в Питерборо-Члене и, кроме плиозавридов, представлены криптоклидидами , включая Cryptoclidus , Muraenosaurus , Tricleidus и Picrocleidus . [ 6 ] : viii [ 2 ] Это были плезиозавры меньшего размера с тонкими зубами и длинной шеей. [ 52 ] и, в отличие от плиозавридов, таких как Peloneustes , питались в основном мелкими животными. [ 52 ] Ихтиозавр Офтальмозавр также обитал в формации Оксфорд-Клэй. Офтальмозавр был хорошо приспособлен к глубокому нырянию благодаря своему обтекаемому, как у морской свиньи , телу и гигантским глазам и, вероятно, питался головоногими моллюсками. [ 6 ] : xiv [ 52 ] Многие роды крокодилов также известны из Питерборо. ним относятся гавиалоподобные К телеозавроиды Charitomenosuchus , Lemmysuchus , Mycterosuchus и Neosteneosaurus. [ 54 ] и мозазавра похожий на [ 52 ] metriorhynchids Gracilineustes , Суходус , Талаттозух , [ 10 ] : 180 [ 55 ] и Тираннонейст . [ 56 ] Маленький рыбоядный птерозавр Rhamphorhynchus , хоть и редкость, но также был частью этой морской экосистемы. [ 52 ]

Из члена Питерборо известно больше видов плиозавридов, чем из любого другого комплекса. [ 2 ] Помимо Peloneustes , к этим плиозавридам относятся Liopleurodonferox , Simolestes vorax , «Pliosaurus» andrewsi , Marmornectes candrewi , [ 57 ] Eardasaurus powelli и, возможно, Pachycostasaurus Dawni . [ 58 ] [ 37 ] Однако существуют значительные различия в анатомии этих видов, что указывает на то, что они питались разной добычей, тем самым избегая конкуренции ( разделение ниш ). [ 42 ] : 249–251 [ 43 ] Крупный и мощный плиозаврид Liopleurodonferox, по-видимому, был приспособлен для охоты на крупную добычу, включая других морских рептилий и крупную рыбу. [ 42 ] : 242–243, 249–251 Длиннорылый Eardasaurus powelli , как и Liopleurodon, также имеет зубы с режущими краями и, возможно, также ловил крупную добычу. [ 37 ] Simolestes vorax с широким и глубоким черепом и мощным укусом, по-видимому, был хищником крупных головоногих моллюсков. [ 42 ] : 243–244, 249–251 «Pliosaurus» andrewsi , как и Peloneustes , обладает удлиненной мордой, приспособленной для питания маленькими, проворными животными. [ 52 ] Однако его зубы приспособлены для резки, что указывает на предпочтение более крупной добычи, тогда как зубы Пелоневста лучше приспособлены для прокалывания. [ 59 ] «Плиозавр» andrewsi также крупнее Пелоневста . [ 52 ] Marmornectes candrewi также похож на Peloneustes , имеет длинную морду и, возможно, также питается рыбой. [ 33 ] [ 57 ] Pachycostasaurus Dawni — небольшой, массивный плиозавр, который, вероятно, питался донной добычей. У него более слабый череп, чем у других плиозавридов, и он был более стабилен, поэтому, вероятно, он использовал разные методы питания, чтобы избежать конкуренции. [ 43 ] В отличие от других плиозавридов Оксфордской глиняной формации, пахикостазавр был довольно редким, возможно, в основном обитавшим за пределами зоны отложения Оксфордской глиняной формации, возможно, населяя вместо этого прибрежные районы, глубокие воды или даже реки. [ 43 ] Хотя в средней юре присутствовало несколько различных типов плиозавридов, длиннорылые рыбоядные формы, такие как Peloneustes, вымерли на границе средней и верхней юры. Похоже, это была первая фаза постепенного сокращения разнообразия плезиозавров. Хотя причина этого неясна, возможно, на это повлияло изменение химического состава океана, а на более поздних этапах - падение уровня моря. [ 33 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Коэн, К.М.; Финни, С.; Гиббард, Польша (2015). «Международная хроностратиграфическая карта» (PDF) . Международная комиссия по стратиграфии.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из Кетчум, HF; Бенсон, RBJ (2011). «Черепная анатомия и таксономия Peloneustes philarchus (Sauropterygia, Pliosauridae) из члена Питерборо (келловей, средняя юра) Соединенного Королевства». Палеонтология . 54 (3): 639–665. Бибкод : 2011Palgy..54..639K . дои : 10.1111/j.1475-4983.2011.01050.x . S2CID 85851352 .
- ^ Перейти обратно: а б с д и ж Сили, Х.Г. (1869). Указатель ископаемых останков Aves, Ornithosauria и Reptilia из вторичной системы слоев, хранящихся в Вудвардском музее Кембриджского университета . Кембридж, Дейтон, Белл и компания. стр. 139–140.
- ^ Перейти обратно: а б Крейслер, Б. (2012). «Руководство Бена Крейслера по произношению плезиозавров» . Океаны Канзаса . Проверено 26 июня 2021 г.
- ^ Араужо, Р.; Смит, А.С.; Листон, Дж. (2008). «Коллекция Альфреда Лидса Национального музея Ирландии - естественная история» (PDF) . Ирландский журнал наук о Земле . 26 :17–32. дои : 10.3318/IJES.2008.26.17 . S2CID 131513080 .
- ^ Перейти обратно: а б с д и Эндрюс, CW (1910). Описательный каталог морских рептилий Оксфордской глины. На основе коллекции Лидса в Британском музее (естественной истории) в Лондоне . Том. 1. Лондон: Британский музей.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Тарло, Л.Б. (1960). «Обзор верхнеюрских плиозавров» . Бюллетень Британского музея (Естественная история) . 4 (5): 145–189.
- ^ Перейти обратно: а б с Лидеккер, Р. (1889). «О остатках и родстве пяти родов мезозойских рептилий» . Ежеквартальный журнал Лондонского геологического общества . 45 (1–4): 41–59. дои : 10.1144/GSL.JGS.1889.045.01-04.04 . S2CID 128586645 .
- ^ Перейти обратно: а б Сили, Х.Г. (1892). «Характер плечевого пояса и ключичной дуги при Sauropterygia» . Труды Лондонского королевского общества . 51 : 119–151. Бибкод : 1892RSPS...51..119S .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д Эндрюс, CW (1913). Описательный каталог морских рептилий Оксфордской глины. На основе коллекции Лидса в Британском музее (естественной истории) в Лондоне . Том. 2. Лондон: Британский музей.
- ^ Эндрюс, CW (1895). «О строении черепа Peloneustes philarchus , плиозавра из Оксфордской глины» . Анналы и журнал естественной истории; Зоология, ботаника и геология . 16 (93): 242–256.
- ^ Жаккар, Ф. (1907). «Заметки о Peloneustes philarchus Seeley из Палеонтологического музея Лозанны» . Бюллетень Водуазского общества естественных наук (на французском языке). 43 (160): 395–398.
- ^ Перейти обратно: а б с д Линдер, Х. (1913). «Вклад в знание родов плезиозавров Peloneustes и Pliosaurus » . Геологические и палеонтологические трактаты (на немецком языке). 11 :339-409.
- ^ Перейти обратно: а б Эндрюс, CW (1910). «Заметка о скелете небольшого плиозавра Peloneustes philarchus Seeley» . Геологический журнал . 7 (3): 110–112. дои : 10.1017/S0016756800132960 . S2CID 130045734 .
- ^ Перейти обратно: а б Филлипс, Дж. (1871). Геология Оксфорда и долины Темзы . Оксфорд: Кларендон Пресс.
- ^ Перейти обратно: а б Лидеккер, Р. (1890). Каталог ископаемых рептилий и амфибий в Британском музее (Естественная история). Часть IV. Включает отряды Anomodontia, Ecaudata, Caudata, Labyrinthodonta; и доп . Лондон: Попечители Британского музея. п. 273.
- ^ Перейти обратно: а б Кнутсен, Э.М. (2012). «Таксономический пересмотр рода Pliosaurus (Owen, 1841a) Owen, 1841b» (PDF) . Норвежский геологический журнал . 92 : 259–276.
- ^ Novozhilov, N. (1948). "Два новых плиозавра из нижнего волжского яруса Поволжья" [Two new pliosaurs from the Lower Volga beds Povolzhe (right bank of Volga)] (PDF) . Doklady Akademii Nauk SSSR (in Russian). 60 : 115–118.
- ^ Перейти обратно: а б Сторрс, Г.В.; Архангельский, М.С.; Ефимов, В.М. (2000). «Мезозойские морские рептилии России и других бывших советских республик». В Бентоне, MJ; Шишкин, М.А.; Анвин, DM; Курочкин Е.Н. (ред.). Эпоха динозавров в России и Монголии . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 187–209.
- ^ Перейти обратно: а б с О'Киф, Франция (2001). «Кладистический анализ и таксономический пересмотр Plesiosauria (Reptilia: Sauropterygia)» . АктаЗоология Фенника . 213 : 1–63.
- ^ Гаспарини, З.; Итурральде-Винент, Массачусетс (2006). «Кубинская оксфордская герпетофауна в Карибском море» (PDF) . Новогодний ежегодник геологии и палеонтологии - Трактаты . 240 (3): 343–371. дои : 10.1127/njgpa/240/2006/343 . S2CID 55346096 . Архивировано из оригинала (PDF) 5 июня 2020 г.
- ^ Гаспарини, З. (2009). «Новый оксфордский плиозаврид (Plesiosauria, Pliosauridae) в Карибском море» . Палеонтология . 52 (3): 661–669. Бибкод : 2009Palgy..52..661G . дои : 10.1111/j.1475-4983.2009.00871.x . S2CID 55353949 .
- ^ Перейти обратно: а б с д МакГенри, ЧР (2009). Пожиратель богов: палеоэкология мелового плиозавра Kronosaurus queenslandicus (Диссертация). Университет Ньюкасла. hdl : 1959.13/935911 . S2CID 132852950 .
- ^ Перейти обратно: а б с О'Киф, Франция (2001). «Экоморфология геометрии ласт плезиозавра» . Журнал эволюционной биологии . 14 (6): 987–991. дои : 10.1046/j.1420-9101.2001.00347.x . S2CID 53642687 .
- ^ Сассун, Дж.; Фоффа, Д.; Марек, Р. (2015). «Онтогенез и замена зубов у Pliosauridae» . Королевское общество открытой науки . 2 (11): 150384. Бибкод : 2015RSOS....250384S . дои : 10.1098/rsos.150384 . ПМЦ 4680613 . ПМИД 26715998 .
- ^ Перейти обратно: а б Смит, А.С. (2013). «Морфология хвостовых позвонков Rhomaleosaurus zetlandicus и обзор доказательств существования хвостового плавника у Plesiosauria» (PDF) . Палудикола . 9 (3): 144–158.
- ^ Сили, Х.Г. (1874 г.). «Обратите внимание на некоторые общие модификации плезиозавровой грудной дуги» . Ежеквартальный журнал Геологического общества . 30 (1–4): 436–449. дои : 10.1144/gsl.jgs.1874.030.01-04.48 . S2CID 128746688 .
- ^ Уайт, Т.Е. (1940). «Голотип Plesiosaurus longirostris Blake и классификация плезиозавров». Журнал палеонтологии . 14 (5): 451–467.
- ^ Перссонс, ПО (1963). «Пересмотр классификации Plesiosauria с кратким описанием стратиграфического и географического распространения группы» (PDF) . Лундский университет Арсскрифт . 59 (1): 1–59.
- ^ Перейти обратно: а б Смит, А.С.; Дайк, Дж.Дж. (2008). «Череп гигантского хищного плиозавра Rhomaleosaurus cramptoni : значение для филогенетики плезиозавров» (PDF) . Naturwissenschaften . 95 (10): 975–980. Бибкод : 2008NW.....95..975S . дои : 10.1007/s00114-008-0402-z . ПМИД 18523747 . S2CID 12528732 .
- ^ Карпентер, К. (1997). «Сравнительная черепная анатомия двух североамериканских плезиозавров мелового периода». В Каллауэе, ИМ; Николлс, Э.Л. (ред.). Древние морские рептилии . Академическая пресса. стр. 191–216.
- ^ Перейти обратно: а б Бенсон, РБЖ; Эванс, М.; Друкенмиллер, PS (2012). «Высокое разнообразие, низкая разница и небольшой размер тела у плезиозавров (Reptilia, Sauropterygia) на границе триаса и юры» . ПЛОС ОДИН . 7 (3): e31838. Бибкод : 2012PLoSO...731838B . дои : 10.1371/journal.pone.0031838 . ПМК 3306369 . ПМИД 22438869 .
- ^ Перейти обратно: а б с Бенсон, РБЖ; Дракенмиллер, PS (2013). «Фаунистический оборот морских четвероногих во время юрско-мелового перехода». Биологические обзоры . 89 (1): 1–23. дои : 10.1111/brv.12038 . ПМИД 23581455 . S2CID 19710180 .
- ^ Бенсон, РБЖ; Эванс, М.; Смит, А.С.; Сассун, Дж.; Мур-Фэй, С.; Кетчум, HF; Форрест, Р. (2013). «Гигантский череп плиозавра из поздней юры Англии» . ПЛОС ОДИН . 8 (5): e65989. Бибкод : 2013PLoSO...865989B . дои : 10.1371/journal.pone.0065989 . ПМК 3669260 . ПМИД 23741520 .
- ^ Фишер, В.; Архангельский, М.С.; Стеньшин И.М.; Успенский Г.Н.; Зверков Н.Г.; Бенсон, RBJ (2015). «Особенные макрофагальные адаптации у нового мелового плиозаврид» . Королевское общество открытой науки . 2 (12): 150552. Цифровой код : 2015RSOS....250552F . дои : 10.1098/rsos.150552 . ПМЦ 4807462 . ПМИД 27019740 .
- ^ Фишер, В.; Бенсон, РБЖ; Зверков Н.Г.; Соул, ЖК; Архангельский, М.С.; Ламберт, О.; Стеньшин И.М.; Успенский Г.Н.; Друкенмиллер, PS (2017). «Пластичность и конвергенция в эволюции плезиозавров с короткой шеей» (PDF) . Современная биология . 27 (11): 1667–1676. дои : 10.1016/j.cub.2017.04.052 . ПМИД 28552354 . S2CID 39217763 .
- ^ Перейти обратно: а б с Кетчум, HF; Бенсон, RBJ (2022). «Новый плиозаврид из формации Оксфорд-Клей в Оксфордшире, Великобритания» . Acta Palaeontologica Polonica . 67 . дои : 10.4202/app.00887.2021 . S2CID 249034986 .
- ^ Перейти обратно: а б Нинан, Дж. М.; Райх, Т.; Эверс, Юго-Запад; Дракенмиллер, PS; Воетен, DFAE; Шуаньер, Ж.Н.; Барретт, премьер-министр; Пирс, ЮВ; Бенсон, RBJ (2017). «Эволюция зауроптеригического лабиринта с все более пелагическим образом жизни» (PDF) . Современная биология . 27 (24): 3852–3858. дои : 10.1016/j.cub.2017.10.069 . ПМИД 29225027 . S2CID 207053689 .
- ^ Перейти обратно: а б Флейшле, резюме; Сандер, премьер-министр; Винтрич, Т.; Каспар, КР (2019). «Гематологическая конвергенция между мезозойскими морскими рептилиями (Sauropterygia) и современными водными амниотами проясняет адаптацию плезиозавров к нырянию» . ПерДж . 7 : е8022. дои : 10.7717/peerj.8022 . ПМК 6873879 . ПМИД 31763069 .
- ^ Перейти обратно: а б Флейшле, резюме; Винтрич, Т.; Сандер, премьер-министр (2018). «Количественные гистологические модели предполагают эндотермию у плезиозавров» . ПерДж . 6 : е4955. дои : 10.7717/peerj.4955 . ПМЦ 5994164 . ПМИД 29892509 .
- ^ Хусай, А. (2013). «Гистология костей водных рептилий: что она говорит нам о вторичной адаптации к водной жизни?». Биологический журнал Линнеевского общества . 108 (1): 3–21. дои : 10.1111/j.1095-8312.2012.02002.x .
- ^ Перейти обратно: а б с д и ж Ноэ, LF (2001). Таксономическое и функциональное исследование келловейских (среднеюрских) Pliosauroidea (Reptilia, Sauropterygia) (доктор философии). Чикаго: Университет Дерби.
- ^ Перейти обратно: а б с д Круикшанк, ARI; Мартилл, DM; Ноэ, Л.Ф. (1996). «Плиозавр (Reptilia, Sauropterygia) с пахиостозом из средней юры Англии» . Журнал Геологического общества . 153 (6): 873–879. Бибкод : 1996JGSoc.153..873C . дои : 10.1144/gsjgs.153.6.0873 . S2CID 129602868 .
- ^ Перейти обратно: а б Массаре, Дж. А. (1988). «Плавательные способности мезозойских морских рептилий: значение для метода хищничества». Палеобиология . 14 (2): 187–205. Бибкод : 1988Pbio...14..187M . дои : 10.1017/S009483730001191X . S2CID 85810360 .
- ^ Гутарра, С.; Стаббс, ТЛ; Мун, Британская Колумбия; Палмер, К.; Бентон, MJ (2022). «Большой размер водных четвероногих компенсирует высокое сопротивление, вызванное чрезмерными пропорциями тела» . Коммуникационная биология . 5 (1): 380. doi : 10.1038/s42003-022-03322-y . ПМК 9051157 . ПМИД 35484197 .
- ^ Ротшильд, Б.М.; Кларк, НДЛ; Кларк, CM (2018). «Доказательства выживания среднеюрского плезиозавра с патологией плечевой кости: какой мы можем сделать вывод о поведении плезиозавра?» . Электронная палеонтология . 21 (16): 1–11. дои : 10.26879/719 . S2CID 133806762 .
- ^ Перейти обратно: а б с д Дафф, К.Л. «Палеоэкология битуминозных сланцев – глина Нижнего Оксфорда центральной Англии» . Палеонтология . 18 (3): 443–482.
- ^ Перейти обратно: а б с д и ж г Меттам, К.; Джонсон, Алабама; Нанн, EV; Шёне, БР (2014). «Стабильный изотоп (δ 18 О и δ 13 C) склерохронология келловейских (среднеюрских) двустворчатых моллюсков ( Gryphaea (Bilobissa) dilobotes ) и белемнитов ( Cylindroteuthis puzosiana ) из пачки Питерборо формации Оксфорд-Клэй (Кембриджшир, Англия): свидетельства палеоклимата, глубины воды и поведения белемнитов » (PDF) ) . Палеогеография, Палеоклиматология, Палеоэкология . 399 : 187–201. Бибкод : 2014PPP... . doi : j.palaeo.2014.01.010 . hdl : 10545/592777 . / 10.1016 399..187M
- ^ Перейти обратно: а б Хадсон, доктор медицинских наук; Мартилл, Д. (1994). «Пачка Питерборо (келловей, средняя юра) формации Оксфордская глина в Питерборо, Великобритания» . Журнал Геологического общества . 151 (1): 113–124. Бибкод : 1994JGSoc.151..113H . дои : 10.1144/gsjgs.151.1.0113 . S2CID 130058981 .
- ^ Перейти обратно: а б с Белин, С.; Кениг, Ф. (1994). «Петрографический анализ органо-минеральных связей: условия отложения оксфордской глиняной формации (юрский период), Великобритания». Журнал Геологического общества . 151 (1): 153–160. Бибкод : 1994JGSoc.151..153B . CiteSeerX 10.1.1.1001.7308 . дои : 10.1144/gsjgs.151.1.0153 . S2CID 131433536 .
- ^ Кениг, Ф; Хадсон, доктор медицинских наук; Дамсте, JSS; Попп, Б.Н. (2004). «Периодическая эвксиния: согласование юрских черных сланцев с их биофациями» . Геология . 32 (5): 421–424. Бибкод : 2004Geo....32..421K . дои : 10.1130/G20356.1 .
- ^ Перейти обратно: а б с д и ж г час я дж Мартилл, DM; Тейлор, Массачусетс; Дафф, КЛ; Райдинг, Джей Би; Баун, PR (1994). «Трофическая структура биоты члена Питерборо, формация Оксфордская глина (юрский период), Великобритания» . Журнал Геологического общества . 151 (1): 173–194. Бибкод : 1994JGSoc.151..173M . дои : 10.1144/gsjgs.151.1.0173 . S2CID 131200898 .
- ^ Мартилль, ДМ (1991), «Рыба», в Мартилле, ДМ; Хадсон, доктор медицинских наук (ред.), Окаменелости Оксфордской глины (PDF) , Лондон: Палеонтологическая ассоциация, стр. 197–225, ISBN 0901702463
- ^ Джонсон, ММ; Янг, Монтана; Брусатте, СЛ (2020). «Филогенетика Teleosauroidea (Crocodylomorpha, Thalattosuria) и значение их экологии и эволюции» . ПерДж . 8 : е9808. дои : 10.7717/peerj.9808 . ПМЦ 7548081 . ПМИД 33083104 .
- ^ Янг, Монтана; Бриньон, А.; Сакс, С.; Хорнунг, Джей-Джей; Фоффа, Д.; Китсон, JJN; Джонсон, ММ; Стил, Л. (2021). «Разрезание гордиева узла: исторический и таксономический пересмотр юрского крокодиломорфа Metriorhynchus » . Зоологический журнал Линнеевского общества . 192 (2): 510–553. doi : 10.1093/zoolinnean/zlaa092 .
- ^ Сакс, С.; Янг, Монтана; Абель, П.; Мэллисон, Х. (2019). «Новый вид метриоринхидного крокодиломорфа Cricosaurus из верхней юры южной Германии» (PDF) . Acta Palaeontologica Polonica . 64 (2): 343–356. дои : 10.4202/app.00541.2018 .
- ^ Перейти обратно: а б Кетчум, HF; Бенсон, RBJ (2011). «Новый плиозаврид (Sauropterygia, Plesiosauria) из формации Оксфордской глины (средняя юра, келловей) Англии: свидетельства изящного, лонгиростринового сорта плиозавридов ранней и средней юры». Специальные статьи по палеонтологии . 86 : 109–129. ISSN 0038-6804 . ОСЛК 2450768 .
- ^ Ноэ, LF; Листон, Дж.; Эванс, М. (2003). «Первый относительно полный экзоципитально-опистоз из черепной коробки келловейского плиозавра Лиоплевродона » (PDF) . Геологический журнал . 140 (4): 479–486. Бибкод : 2003ГеоМ..140..479Н . дои : 10.1017/S0016756803007829 . S2CID 22915279 . Архивировано из оригинала (PDF) 9 июня 2020 года.
- ^ Массаре, Дж. А. (1987). «Морфология зубов и предпочтения добычи мезозойских морских рептилий». Журнал палеонтологии позвоночных . 7 (2): 121–137. Бибкод : 1987JVPal...7..121M . дои : 10.1080/02724634.1987.10011647 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Пелоневстом, на Викискладе?
СМИ, связанные с Пелоневстом, на Викискладе?






