Зеленые серные бактерии
| Зеленые серные бактерии | |
|---|---|
 | |
| Зеленые серобактерии в колонке Виноградского | |
| Научная классификация | |
| Домен: | Бактерии |
| (без рейтинга): | Группа Бактероидота-Хлоробиота. |
| Тип: | Хлоробиота Иино и др. 2021 год [3] |
| Сорт: | «Хлоробия» Гаррити и Холт, 2001 г. [2] |
| Заказ: | Хлоробиалесы Гиббонс и Мюррей 1978 г. (утвержденные списки 1980 г.) [1] |
| Семьи и роды | |
| Синонимы | |
| |
Зеленые серобактерии относятся типу Chlorobiota к . [4] облигатно анаэробных фотоавтотрофных бактерий , метаболизирующих серу. [5]
Зеленые серные бактерии неподвижны (за исключением Chloroherpeton thalassium , который может скользить) и способны к аноксигенному фотосинтезу . [5] [6] Они живут в анаэробной водной среде. [7] В отличие от растений зеленые серобактерии в качестве доноров электронов используют преимущественно сульфид-ионы. [8] Они являются автотрофами , которые используют обратный цикл трикарбоновых кислот для фиксации углерода . [9] Они также являются миксотрофами и восстанавливают азот. [10] [11]
Характеристики [ править ]
Зеленые серобактерии представляют собой грамотрицательные палочковидные или шаровидные бактерии. Некоторые виды зеленых серобактерий имеют газовые вакуоли, обеспечивающие движение. Они являются фотолитоавтотрофами и используют энергию света и восстановленные соединения серы в качестве источника электронов. [12] К донорам электронов относятся H 2 , H 2 S, S. Основным фотосинтетическим пигментом этих бактерий являются бактериохлорофиллы c или d у зеленых видов и е у коричневых видов, которые расположены в хлоросомах и плазматических мембранах. [7] Хлоросомы — уникальная особенность, позволяющая им улавливать свет в условиях низкой освещенности. [13]
Среда обитания [ править ]
Большинство зеленых серобактерий мезофильны , предпочитают умеренные температуры и живут в водной среде. Они требуют анаэробных условий и пониженного содержания серы; они обычно находятся в верхних миллиметрах отложений. Они способны к фотосинтезу в условиях низкой освещенности. [7]
Было обнаружено, что в Черном море , чрезвычайно бескислородной среде, на глубине около 100 м обитает большая популяция зеленых серных бактерий. Из-за нехватки света в этом районе моря большинство бактерий были фотосинтетически неактивны. Фотосинтетическая активность, обнаруженная в сульфидном хемоклине, позволяет предположить, что бактериям требуется очень мало энергии для поддержания клеток. [14]
Вид зеленых серобактерий был обнаружен живущим рядом с черным курильщиком у побережья Мексики на глубине 2500 м в Тихом океане . На этой глубине бактерия, получившая обозначение GSB1, живет за счет тусклого свечения теплового источника, поскольку солнечный свет не может проникнуть на эту глубину. [15]
Зеленые серные бактерии также были обнаружены живущими на колониях коралловых рифов на Тайване. Они составляют большую часть «зеленого слоя» этих колоний. Вероятно, они играют роль в коралловой системе, и между бактериями и кораллом-хозяином могут существовать симбиотические отношения. [16] Коралл может обеспечить анаэробную среду и источник углерода для бактерий. Бактерии могут обеспечивать питательные вещества и очищать кораллы от токсинов, окисляя сульфид. [17]
один тип зеленых серобактерий, Chlorobaculum tepidum В серных источниках был обнаружен . Эти организмы термофильны , в отличие от большинства других зеленых серобактерий. [7]
Филогения [ править ]
| на основе 16S рРНК ДП _08_2023 [18] [19] [20] | 120 маркерных белков на основе GTDB 08-RS214 [21] [22] [23] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Таксономия [ править ]
- Семейство Chlorobiaceae Copeland 1956 ["Chlorobacteriaceae" Geitler & Pascher 1925 ]
- ? Ancalochloris Gorlenko and Lebedeva 1971
- Хлоробакул Имхофф 2003
- Хлоробий Надсон 1906 г.
- « Хлороплана » Дубинина и Горленко 1975 г.
- « Клатрохлорис » Гейтлер 1925 г.
- Простекохлорис Горленко 1970 г.
- Семейство « Термохлоробактериевые » , корриг. Лю и др. 2012 ["Chloroherpetonaceae" Белло и др. 2022 ]
- Хлорогерпетон Гибсон и др. 1985 год
- « Ca. Thermochromobacter » Liu et al. 2012 год
Видовые характеристики родов [ править ]
Зеленые серобактерии относятся к семейству Chlorobiaceae. Есть четыре рода; Хлорогерпетон , Простекохлорис , Хлоробий и Хлоробакулум . Характеристики, используемые для различия этих родов, включают некоторые метаболические свойства, пигменты, морфологию клеток и спектры поглощения. Однако различить эти свойства сложно, поэтому таксономическое деление иногда неясно. [24]
Обычно Chlorobium имеют палочковидную или вибровидную форму, а некоторые виды содержат газовые пузырьки. Они могут развиваться как одиночные или совокупные клетки. Они могут быть зелеными или темно-коричневыми. Зеленые штаммы используют фотосинтетические пигменты Bchl c или d с хлоробактеновыми каротиноидами, а коричневые штаммы используют фотосинтетический пигмент Bchl e с изорениератиновыми каротиноидами. Для роста требуется небольшое количество соли. [24]
Простекохлоры состоят из виброидных, яйцевидных или палочковидных клеток. Они начинаются с одиночных клеток, образующих неветвящиеся придатки, называемые неветвящимися протезами. Они также могут образовывать газовые пузырьки. Присутствующие фотосинтетические пигменты включают Bchl c, d или e. Кроме того, соль необходима для роста. [24]
Хлоробакулум развивается в виде одиночных клеток и обычно имеет виброидную или палочковидную форму. Некоторые из них могут образовывать газовые пузырьки. Фотосинтетические пигменты этого рода — Bchl c, d или e . Некоторым видам для роста требуется NaCl (хлорид натрия). Представители этого рода раньше входили в состав рода Chlorobium, но образовали отдельную линию. [24]
Род Chloroherpeton уникален, поскольку представители этого рода подвижны. Они представляют собой гибкие длинные стержни и могут передвигаться, скользя. Они зеленого цвета и содержат фотосинтетический пигмент Bchl c, а также γ-каротин . Соль необходима для роста. [24]
Метаболизм [ править ]
Фотосинтез [ править ]
Зеленые серные бактерии используют для фотосинтеза реакционный центр типа I. Реакционные центры типа I являются бактериальным гомологом фотосистемы I (PSI) у растений и цианобактерий . Реакционные центры GSB содержат бактериохлорофилл а и известны как реакционные центры P840 из-за длины волны возбуждения 840 нм, которая обеспечивает поток электронов. У зеленых серобактерий реакционный центр связан с большим комплексом антенн, называемым хлоросомой , который захватывает и направляет световую энергию в реакционный центр. Хлоросомы имеют пик поглощения в дальней красной области спектра между 720 и 750 нм, поскольку они содержат бактериохлорофиллы c, d и e. [25] Белковый комплекс, называемый комплексом Фенны-Мэтьюза-Олсона (FMO), физически расположен между хлоросомами и RC P840. Комплекс ФМО помогает эффективно передавать поглощенную антенной энергию в реакционный центр.
Реакционные центры PSI и типа I способны восстанавливать ферредоксин (Fd), сильный восстановитель, который можно использовать для фиксации CO.
2 и уменьшить НАД + . Как только реакционный центр (RC) отдал электрон Fd, он становится окислителем (P840 + ) с восстановительным потенциалом около +300 мВ. Хотя этого недостаточно, чтобы отобрать электроны у воды и синтезировать O.
2 ( Э
0 = +820 мВ), он может принимать электроны из других источников, таких как H
2 S , тиосульфат или Fe 2+
ионы. [26] Этот транспорт электронов от доноров, таких как H
2 S к акцептору Fd называется линейным потоком электронов или линейным транспортом электронов. Окисление сульфид-ионов приводит к образованию серы в качестве побочного продукта, который накапливается в виде глобул на внеклеточной стороне мембраны. Эти шарики серы дали название зеленым серным бактериям. Когда сульфид истощается, шарики серы расходуются и далее окисляются до сульфата. Однако путь окисления серы недостаточно изучен. [8]
Вместо передачи электронов на Fd кластеры Fe-S в реакционном центре P840 могут передавать электроны менахинону (MQ: MQH
2 ), который возвращает электроны в P840 + через электрон-транспортную цепь (ЭТЦ). На обратном пути в РЦ электроны от MQH2 проходят через комплекс цитохрома bc 1 (аналогичный комплексу III митохондрий), который перекачивает H +
ионы через мембрану. Электрохимический потенциал протонов через мембрану используется для синтеза АТФ F o F 1 с помощью АТФ-синтазы . Этот циклический транспорт электронов отвечает за преобразование энергии света в клеточную энергию в форме АТФ. [25]
Метаболизм серы [ править ]
Зеленые серные бактерии окисляют неорганические соединения серы, чтобы использовать их в качестве доноров электронов для анаэробного фотосинтеза, в частности, для фиксации углекислого газа. В качестве донора электронов они обычно предпочитают использовать сульфид, а не другие соединения серы, однако они могут использовать тиосульфат или H 2 . [27] Промежуточным продуктом обычно является сера, которая откладывается вне клетки. [28] и конечный продукт - сульфат. Сера, откладывающаяся внеклеточно, имеет форму серных глобул, которые впоследствии могут быть полностью окислены. [27]
Механизмы окисления серы у зеленых серобактерий изучены недостаточно. Некоторые ферменты, которые, как полагают, участвуют в окислении сульфидов, включают флавоцитохром с, сульфид: хинон оксидоредуктазу и SO.
система х . Флавоцитохром может катализировать перенос электронов на цитохромы из сульфида, и эти цитохромы могут затем перемещать электроны в реакционный центр фотосинтеза. Однако не все зеленые серобактерии производят этот фермент, что свидетельствует о том, что он не необходим для окисления сульфида. Сульфид:хиноноксидоредуктаза (SQR) также способствует транспорту электронов, но, как было обнаружено, она снижает скорость окисления сульфидов у зеленых серных бактерий, что позволяет предположить, что существует другой, более эффективный механизм. [27] Однако большинство зеленых серных бактерий содержат гомолог гена SQR. [29] Окисление тиосульфата до сульфата может катализироваться ферментами SO .
система х . [27]
Считается, что ферменты и гены, связанные с метаболизмом серы, были получены путем горизонтального переноса генов в ходе эволюции зеленых серобактерий. [29]
Фиксация углерода [ править ]
Зеленые серные бактерии являются фотоавтотрофами : они не только получают энергию от света, но и могут расти, используя углекислый газ в качестве единственного источника углерода. Они фиксируют углекислый газ, используя обратный цикл трикарбоновых кислот (rTCA). [9] где энергия расходуется на восстановление углекислого газа, а не на окисление, как это видно в прямом цикле ТСА , [9] для синтеза пирувата и ацетата . Эти молекулы используются в качестве сырья для синтеза всех строительных блоков, необходимых клетке для создания макромолекул . Цикл rTCA очень энергоэффективен, позволяя бактериям расти в условиях низкой освещенности. [30] Однако он содержит несколько чувствительных к кислороду ферментов, что ограничивает его эффективность в аэробных условиях. [30]
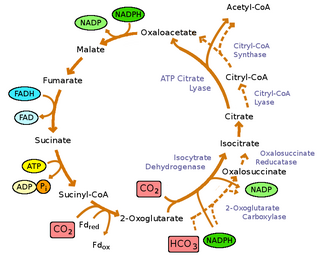
Реакции обращения окислительного цикла трикарбоновых кислот катализируются четырьмя ферментами: [9]
- пируват:ферредоксин (Fd)оксидоредуктаза:
- ацетил-КоА + СО 2 + 2Fdred + 2H+ ⇌ пируват + КоА + 2Fdox
- АТФ-цитрат-лиаза:
- ACL, ацетил-КоА + оксалоацетат + АДФ + Пи ⇌ цитрат + КоА + АТФ
- α-кетоглутарат:ферредоксиноксидоредуктаза:
- сукцинил-КоА + СО 2 + 2Fdred + 2H+ ⇌ α-кетоглутарат + КоА + 2Fdox
- фумаарередуктаза
- сукцинат + акцептор ⇌ фумарат + восстановленный акцептор
Однако окислительный цикл ТСА (ОТЦА) все еще присутствует у зеленых серобактерий. OTCA может ассимилировать ацетат, однако OTCA, по-видимому, является неполным у зеленых серных бактерий из-за расположения и подавления гена во время фототрофного роста. [9]
Миксотрофия [ править ]
Зеленые серные бактерии часто называют облигатными фотоавтотрофами, поскольку они не могут расти в отсутствие света, даже если им обеспечены органические вещества. [9] [26] Однако они проявляют форму миксотрофии , при которой могут потреблять простые органические соединения в присутствии света и CO 2 . [9] В присутствии CO 2 или HCO 3 − Некоторые зеленые серобактерии могут использовать ацетат или пируват. [9]
Миксотрофию зеленых серобактерий лучше всего моделирует представительная зеленая серная бактерия Chlorobaculum tepidum . [31] Миксотрофия возникает во время биосинтеза аминокислот/утилизации углерода и энергетического метаболизма. [32] Бактерия использует электроны, образующиеся в результате окисления серы, и энергию света, которую она захватывает, для запуска rTCA. C. tepidum также демонстрирует использование пирувата и ацетата в качестве источника органического углерода. [32]
Примером миксотрофии C. tepidum , сочетающей автотрофность и гетеротрофию, является синтез ацетил-КоА. C. tepidum может автотрофно генерировать ацетил-КоА посредством цикла rTCA или гетеротрофно генерировать его за счет поглощения ацетата. Аналогичная миксотрофная активность возникает, когда пируват используется для биосинтеза аминокислот, но миксотрофный рост с использованием ацетата дает более высокие скорости роста. [31] [32]
В энергетическом метаболизме C. tepidum полагается на световые реакции для производства энергии (НАДФН и НАДН), поскольку пути, обычно отвечающие за производство энергии (окислительный пентозофосфатный путь и нормальный цикл ТСА), функционируют лишь частично. [32] Фотоны, поглощаемые светом, используются для производства НАДФН и НАДН, кофакторов энергетического обмена. C. tepidum также генерирует энергию в форме АТФ, используя движущую силу протонов, образующуюся в результате окисления сульфида. [31] Производство энергии как за счет окисления сульфидов, так и за счет поглощения фотонов бактериохлорофиллами . [32]
Фиксация азота [ править ]
Большинство зеленых серобактерий являются диазотрофами : они способны восстанавливать азот до аммиака, который затем используется для синтеза аминокислот. [33] Азотфиксация среди зеленых серобактерий обычно типична для аноксигенных фототрофов и требует присутствия света. Зеленые серные бактерии проявляют активность системы секреции типа 1 и ферредоксин-НАДФ + оксидоредуктазы, производя восстановленное железо - признак, который развился для поддержки фиксации азота. [34] Подобно пурпурным серным бактериям, они могут регулировать активность нитрогеназы посттрансляционно в ответ на концентрацию аммиака. Обладание ими генами nif , даже несмотря на то, что они эволюционно различны, может свидетельствовать о том, что их способности к фиксации азота возникли в результате двух разных событий или от общего очень далекого предка. [33]
Примеры зеленых серобактерий, способных к фиксации азота, включают род Chlorobium и Pelodictyon , за исключением P. phaeoclathratiforme . Prosthecochromis aestuarii и Chloroherpeton thalassium также попадают в эту категорию. [33] Их фиксация N 2 широко распространена и играет важную роль в общей доступности азота для экосистем. Зеленые серные бактерии, живущие на коралловых рифах, такие как Prosthecochromis , играют решающую роль в производстве доступного азота в и без того ограниченной питательными веществами среде. [16]
См. также [ править ]
- Аноксическое событие
- Пурпурные серобактерии
- Зеленые несерные бактерии
- Список родов бактерий
- Список порядка бактерий
Ссылки [ править ]
- ^ Гиббонс Н.Э., Мюррей Р.Г.Е. (1978). «Предложения о высших таксонах бактерий» . Международный журнал систематической бактериологии . 28 : 1–6. дои : 10.1099/00207713-28-1-1 .
- ^ Гэррити, Холт Дж.Г. (2001). «Тип BXI. Chlorobi phy. nov.». В Boone DR, Castenholz RW, Garrity GM (ред.). Руководство Берджи по систематической бактериологии . Том. 1 (Археи и глубоко ветвящиеся и фототрофные бактерии) (2-е изд.). Нью-Йорк, штат Нью-Йорк: Спрингер – Верлаг. стр. 601–623.
- ^ Орен А., генеральный директор Гаррити (октябрь 2021 г.). «Действительная публикация названий сорока двух типов прокариот» . Международный журнал систематической и эволюционной микробиологии . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . ПМИД 34694987 . S2CID 239887308 .
- ^ «Тип Хлоробиота » . Список названий прокариот, стоящих в номенклатуре . 25907 . Проверено 22 августа 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Брайант Д.А., Фригаард Н.У. (ноябрь 2006 г.). «Освещение фотосинтеза и фототрофии прокариот». Тенденции в микробиологии . 14 (11): 488–496. дои : 10.1016/j.tim.2006.09.001 . ПМИД 16997562 .
- ^ Зеленый БР (2003). Светособирающие антенны в фотосинтезе . Спрингер. п. 8. ISBN 0792363353 .
- ↑ Перейти обратно: Перейти обратно: а б с д Кушкевич И., Прохазка Ю., Гайдач М., Риттманн С.К., Витезова М. (июнь 2021 г.). «Молекулярная физиология анаэробных фототрофных пурпурных и зеленых серных бактерий» . Международный журнал молекулярных наук . 22 (12): 6398. doi : 10.3390/ijms22126398 . ПМЦ 8232776 . ПМИД 34203823 .
- ↑ Перейти обратно: Перейти обратно: а б Сакураи Х., Огава Т., Сига М., Иноуэ К. (июнь 2010 г.). «Неорганическая система окисления серы в зеленых серобактериях». Исследования фотосинтеза . 104 (2–3): 163–176. дои : 10.1007/s11120-010-9531-2 . ПМИД 20143161 . S2CID 1091791 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Тан К.Х., Бланкеншип Р.Э. (ноябрь 2010 г.). «В зеленых серобактериях действуют как прямой, так и обратный циклы ТСА» . Журнал биологической химии . 285 (46): 35848–35854. дои : 10.1074/jbc.M110.157834 . ПМК 2975208 . ПМИД 20650900 .
- ^ Валунд Т.М., Мэдиган М.Т. (январь 1993 г.). «Азотфиксация термофильной зеленой серной бактерией Chlorobium tepidum» . Журнал бактериологии . 175 (2): 474–478. дои : 10.1128/jb.175.2.474-478.1993 . ЧВК 196162 . ПМИД 8093448 .
- ^ Фэн Икс, Тан К.Х., Бланкеншип Р.Э., Тан Ю.Дж. (декабрь 2010 г.). «Анализ метаболических потоков миксотрофного метаболизма зеленой серной бактерии Chlorobaculum tepidum» . Журнал биологической химии . 285 (50): 39544–39550. дои : 10.1074/jbc.M110.162958 . ПМК 2998096 . ПМИД 20937805 .
- ^ «Зеленые серные бактерии - обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 22 апреля 2022 г.
- ^ ЭЛС (1-е изд.). Джон Вили и сыновья, ООО, 30 мая 2001 г. дои : 10.1002/9780470015902.a0000458.pub2 . ISBN 978-0-470-01617-6 . S2CID 82067054 .
- ^ Маршалл Э., Джоглер М., Хессге У., Оверманн Дж. (май 2010 г.). «Крупномасштабное распространение и модели активности чрезвычайно адаптированной к слабому освещению популяции зеленых серных бактерий в Черном море». Экологическая микробиология . 12 (5): 1348–1362. дои : 10.1111/j.1462-2920.2010.02178.x . ПМИД 20236170 .
- ^ Битти Дж.Т., Оверманн Дж., Линс М.Т., Манске А.К., Ланг А.С., Бланкеншип Р.Э. и др. (июнь 2005 г.). «Облигатно фотосинтезирующий бактериальный анаэроб из глубоководных гидротермальных источников» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (26): 9306–9310. Бибкод : 2005PNAS..102.9306B . дои : 10.1073/pnas.0503674102 . ПМК 1166624 . ПМИД 15967984 .
- ↑ Перейти обратно: Перейти обратно: а б Ян Ш., Ли С.Т., Хуан Ч.Р., Ценг Ч.Х., Чан П.В., Чен К.П. и др. (26 февраля 2016 г.). «Распространенность потенциальных азотфиксирующих зеленых серных бактерий в скелете рифообразующего коралла Isopora palifera» . Лимнология и океанография . 61 (3): 1078–1086. Бибкод : 2016LimOc..61.1078Y . дои : 10.1002/lno.10277 . ISSN 0024-3590 . S2CID 87463811 .
- ^ Цай Л., Чжоу Г., Тянь Р.М., Тонг Х., Чжан В., Сунь Дж. и др. (август 2017 г.). «Метагеномный анализ показывает, что зеленая серная бактерия является потенциальным симбионтом кораллов» . Научные отчеты . 7 (1): 9320. Бибкод : 2017NatSR...7.9320C . дои : 10.1038/s41598-017-09032-4 . ПМЦ 5571212 . ПМИД 28839161 .
- ^ «ЛТП» . Проверено 20 ноября 2023 г.
- ^ «Дерево LTP_all в формате Ньюика» . Проверено 20 ноября 2023 г.
- ^ «Примечания к выпуску LTP_08_2023» (PDF) . Проверено 20 ноября 2023 г.
- ^ «Выпуск GTDB 08-RS214» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ "bac120_r214.sp_label" . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ «История таксонов» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и Брянцева И.А., Тарасов А.Л., Кострикина Н.А., Гайсин В.А., Гроуздев Д.С., Горленко В.М. (декабрь 2019). «Prosthecochromis marina sp. nov., новая зеленая серная бактерия из прибрежной зоны Южно-Китайского моря». Архив микробиологии . 201 (10): 1399–1404. дои : 10.1007/s00203-019-01707-y . ПМИД 31338544 . S2CID 198190182 .
- ↑ Перейти обратно: Перейти обратно: а б Хауска Г., Шодль Т., Ремиги Х., Циотис Г. (октябрь 2001 г.). «Реакционный центр зеленых серобактерий(1)» . Биохимика и биофизика Acta . 1507 (1–3): 260–277. дои : 10.1016/S0005-2728(01)00200-6 . ПМИД 11687219 .
- ↑ Перейти обратно: Перейти обратно: а б Лигроне Р. (2019). «Движение к свету: эволюция фотосинтеза» . В Лигроне Р (ред.). Биологические инновации, которые построили мир: путешествие длиной в четыре миллиарда лет по жизни и истории Земли . Чам: Международное издательство Springer. стр. 99–127. дои : 10.1007/978-3-030-16057-9_4 . ISBN 978-3-030-16057-9 . S2CID 189992218 . Проверено 29 января 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Фригаард Н.У., Даль К. (январь 2008 г.). «Обмен серы у фототрофных серных бактерий» . В Пуле РК (ред.). Достижения микробной физиологии . Том. 54. Академическая пресса. стр. 103–200 . Проверено 22 апреля 2022 г.
- ^ ван Гемерден Х (октябрь 1986 г.). «Продукция элементарной серы зелеными и пурпурными серобактериями» . Архив микробиологии . 146 (1): 52–56. дои : 10.1007/BF00690158 . ISSN 1432-072X . S2CID 30812886 .
- ↑ Перейти обратно: Перейти обратно: а б Грегерсен Л.Х., Брайант Д.А., Фригаард Н.У. (2011). «Механизмы и эволюция окислительного обмена серы у зеленых серобактерий» . Границы микробиологии . 2 : 116. дои : 10.3389/fmicb.2011.00116 . ПМК 3153061 . ПМИД 21833341 .
- ↑ Перейти обратно: Перейти обратно: а б Бар-Эвен А., Нур Э., Майло Р. (март 2012 г.). «Обзор путей фиксации углерода через количественную призму» . Журнал экспериментальной ботаники . 63 (6): 2325–2342. дои : 10.1093/jxb/err417 . ПМИД 22200662 .
- ↑ Перейти обратно: Перейти обратно: а б с Фригаард Н.У., Чу А.Г., Ли Х., Мареска Дж.А., Брайант Д.А. (2003). «Chlorobium tepidum: понимание структуры, физиологии и метаболизма зеленой серной бактерии, полученное на основе полной последовательности генома». Исследования фотосинтеза . 78 (2): 93–117. дои : 10.1023/B:PRES.0000004310.96189.b4 . ПМИД 16245042 . S2CID 30218833 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Фэн Икс, Тан К.Х., Бланкеншип Р.Э., Тан Ю.Дж. (декабрь 2010 г.). «Анализ метаболических потоков миксотрофного метаболизма зеленой серной бактерии Chlorobaculum tepidum» . Журнал биологической химии . 285 (50): 39544–39550. дои : 10.1074/jbc.M110.162958 . ПМК 2998096 . ПМИД 20937805 .
- ↑ Перейти обратно: Перейти обратно: а б с Мэдиган М.Т. (1995). «Микробиология фиксации азота аноксигенными фотосинтезирующими бактериями». В Бланкеншипе Р.Э. , Мэдигане М.Т., Бауэре С.Э. (ред.). Аноксигенные фотосинтезирующие бактерии . Достижения в области фотосинтеза и дыхания. Том. 2. Дордрехт: Springer Нидерланды. стр. 915–928. дои : 10.1007/0-306-47954-0_42 . ISBN 978-0-306-47954-0 .
- ^ Мус Ф., Колман Д.Р., Питерс Дж.В., Бойд Э.С. (август 2019 г.). «Геобиологические обратные связи, кислород и эволюция нитрогеназы» . Свободно-радикальная биология и медицина . Ранняя жизнь на Земле и окислительный стресс. 140 : 250–259. doi : 10.1016/j.freeradbiomed.2019.01.050 . PMID 30735835 . S2CID 73433517 .
Внешние ссылки [ править ]
- «Семейство Хлоробиевые» . Прокариоты . Архивировано из оригинала 17 ноября 2003 года . Проверено 5 июля 2005 г.