Долгосрочное потенцирование

В нейробиологии ( долговременная потенциация ) — LTP это постоянное усиление синапсов на основе недавних моделей активности. Это паттерны синаптической активности, которые приводят к длительному усилению передачи сигнала между двумя нейронами . [ 2 ] Противоположностью LTP является длительная депрессия , которая приводит к длительному снижению синаптической силы.
Это одно из нескольких явлений, лежащих в основе синаптической пластичности — способности химических синапсов изменять свою силу. Поскольку считается, что воспоминания кодируются путем изменения силы синапсов , [ 3 ] LTP широко считается одним из основных клеточных механизмов, лежащих в основе обучения и памяти . [ 2 ] [ 3 ]
LTP был обнаружен в гиппокампе кролика Терье Лёмо в 1966 году и с тех пор остается популярным предметом исследований. Многие современные исследования LTP направлены на лучшее понимание ее базовой биологии, в то время как другие направлены на установление причинно-следственной связи между LTP и поведенческим обучением. Тем не менее, другие пытаются разработать методы, фармакологические или иные, усиления LTP для улучшения обучения и памяти. ЛТП также является предметом клинических исследований , например, в области медицины болезни Альцгеймера и наркозависимости .
История
Ранние теории обучения

В конце XIX века учёные вообще признали, что количество нейронов в мозге взрослого человека (примерно 100 миллиардов [ 4 ] ) существенно не увеличивается с возрастом, что дает нейробиологам веские основания полагать, что воспоминания, как правило, не являются результатом образования новых нейронов. [ 5 ] С этим осознанием возникла необходимость объяснить, как воспоминания могут формироваться в отсутствие новых нейронов.
Испанский был одним из нейроанатом Сантьяго Рамон-и-Кахаль первых, кто предложил механизм обучения, не требующий образования новых нейронов. В своей Крунианской лекции 1894 года он предположил, что вместо этого воспоминания могут формироваться путем укрепления связей между существующими нейронами для повышения эффективности их общения. [ 5 ] Теория Хебба , представленная Дональдом Хеббом в 1949 году, перекликалась с идеями Рамона-и-Кахаля, предполагая далее, что клетки могут создавать новые связи или претерпевать метаболические и синаптические изменения, которые повышают их способность общаться и создают нейронную сеть опыта: [ 6 ]
Давайте предположим, что сохранение или повторение реверберационной активности (или «следа») имеет тенденцию вызывать длительные клеточные изменения, которые повышают ее стабильность... Когда аксон клетки А находится достаточно близко, чтобы возбудить клетку В и неоднократно или постоянно участвует в его активации, в одной или обеих клетках происходит некоторый процесс роста или метаболические изменения, в результате чего эффективность A как одной из клеток, запускающих B, увеличивается. [ 7 ]
Эрик Кандел (1964) и его коллеги были одними из первых исследователей, обнаруживших долговременное потенцирование во время работы с морскими слизняками Aplysia. Они попытались применить поведенческую обусловленность к различным клеткам нейронной сети слизняка. Их результаты показали изменения в синаптической силе, и исследователи предположили, что это может быть связано с базовой формой обучения, происходящей внутри слизняка. [ 8 ] [ 9 ]
Хотя эти теории формирования памяти сейчас хорошо известны, для своего времени они были дальновидными: нейробиологи и психологи конца 19-го и начала 20-го века не были оснащены нейрофизиологическими методами, необходимыми для выяснения биологических основ обучения у животных. Эти навыки появились только во второй половине 20-го века, примерно в то же время, когда было открыто долговременное потенцирование.
Открытие
LTP впервые наблюдался Терье Лёмо в 1966 году в Осло , Норвегия , в лаборатории Пера Андерсена . [ 10 ] [ 11 ] Там Лёмо провел серию нейрофизиологических экспериментов на наркотизированных кроликах, чтобы изучить роль гиппокампа в кратковременной памяти .
Эксперименты Лёмо были сосредоточены на соединениях или синапсах от перфорантного пути к зубчатой извилине . Эти эксперименты проводились путем стимуляции пресинаптических волокон перфорантного пути и регистрации ответов от совокупности постсинаптических клеток зубчатой извилины. Как и ожидалось, одиночный импульс электрической стимуляции волокон перфорантного пути вызывал возбуждающие постсинаптические потенциалы (ВПСП) в клетках зубчатой извилины. Лёмо неожиданно обнаружил, что реакция постсинаптических клеток на эти одиночные импульсные стимулы может быть усилена на длительный период времени, если он сначала доставит высокочастотную последовательность стимулов к пресинаптическим волокнам. Когда применялась такая серия стимулов, последующие одиночные импульсные стимулы вызывали более сильные и продолжительные ВПСП в популяции постсинаптических клеток. Это явление, при котором высокочастотный стимул может вызывать долговременное усиление реакции постсинаптических клеток на последующие одноимпульсные стимулы, первоначально называлось «длительной потенциацией». [ 12 ] [ 13 ]
Тимоти Блисс , пришедший в лабораторию Андерсена в 1968 году, [ 10 ] сотрудничал с Лёмо, и в 1973 году они опубликовали первую характеристику длительной потенциации в гиппокампе кролика . [ 12 ] Блисс и Тони Гарднер-Медвин опубликовали аналогичный отчет о длительной потенциации у бодрствующих животных, который появился в том же выпуске, что и отчет Блисс и Лёмо. [ 13 ] В 1975 году Дуглас и Годдард предложили «долговременную потенциацию» как новое название феномена длительной потенциации. [ 14 ] [ 15 ] Андерсен предположил, что авторы выбрали «долговременное потенцирование», возможно, из-за его легко произносимой аббревиатуры «LTP». [ 16 ]
Модели и теория

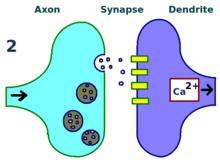
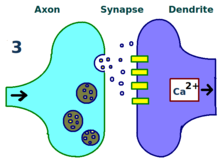

Физический и биологический механизм ЛТП до сих пор не изучен, но было разработано несколько успешных моделей. [1] Исследования дендритных шипиков , выступающих структур на дендритах, которые физически растут и втягиваются в течение минут или часов, предположили взаимосвязь между электрическим сопротивлением шипов и эффективной силой синапсов из-за их связи с внутриклеточными кальциевыми переходными процессами. . Математические модели, такие как теория BCM , которая также зависит от внутриклеточного кальция по отношению к рецептора NMDA воротам напряжения , разрабатываются с 1980-х годов и модифицируют традиционную априорную модель обучения Хебба, имея как биологическое, так и экспериментальное обоснование. Тем не менее, другие предложили перестроить или синхронизировать отношения между регуляцией рецепторов, LTP и синаптической силой. [ 17 ]
Типы
С момента своего первоначального открытия в гиппокампе кролика ДП наблюдалась во множестве других нервных структур, включая кору головного мозга . [ 18 ] мозжечок , [ 19 ] миндалевидное тело , [ 20 ] и многие другие. Роберт Маленка , известный исследователь ДП, предположил, что ДП может возникать даже во всех возбуждающих синапсах мозга млекопитающих. [ 21 ]
В разных областях мозга наблюдаются разные формы ДП. Конкретный тип LTP, проявляющийся между нейронами, зависит от ряда факторов. Одним из таких факторов является возраст организма, в котором наблюдается ДП. Например, молекулярные механизмы ДП в незрелом гиппокампе отличаются от тех механизмов, которые лежат в основе ДП взрослого гиппокампа. [ 22 ] Сигнальные пути, используемые конкретной клеткой, также способствуют появлению определенного типа LTP. Например, некоторые типы LTP гиппокампа зависят от рецептора NMDA , другие могут зависеть от метаботропного рецептора глутамата (mGluR), а третьи вообще зависят от другой молекулы. [ 21 ] Разнообразие сигнальных путей, которые способствуют LTP, и широкое распространение этих различных путей в мозге являются причинами того, что тип LTP, проявляемый между нейронами, лишь частично зависит от анатомического местоположения, в котором наблюдается LTP. Например, ДП в коллатеральном пути Шаффера гиппокампа является NMDA-рецептор-зависимой - это было доказано применением AP5 , антагониста NMDA-рецептора, который предотвращал ДП в этом пути. [ 23 ] И наоборот, LTP в пути мшистых волокон не зависит от рецептора NMDA, хотя оба пути находятся в гиппокампе. [ 24 ]
Пре- и постсинаптическая активность, необходимая для индукции ДП, являются другими критериями, по которым классифицируется ДП. В общих чертах это позволяет классифицировать LTP на хеббианские, нехеббианские и антихеббовские механизмы. Заимствуя свое название из постулата Хебба , резюмируемого максимой о том, что «клетки, которые срабатывают вместе, соединяются вместе», Хеббианская ДП требует одновременной пре- и постсинаптической деполяризации для ее индукции. [ 25 ] Нехеббианская LTP — это тип LTP, который не требует такой одновременной деполяризации пре- и постсинаптических клеток; пример этого происходит в гиппокампальном пути мшистых волокон. [ 26 ] Особый случай нехеббианской LTP, антихеббианская LTP явно требует одновременной пресинаптической деполяризации и относительной постсинаптической гиперполяризации для ее индукции. [ 27 ]
Благодаря своей предсказуемой организации и легко индуцируемой ДП гиппокамп CA1 стал прототипным местом исследования ДП млекопитающих. В частности, NMDA-рецептор-зависимая ДП в гиппокампе СА1 взрослых является наиболее широко изученным типом ДП. [ 21 ] и поэтому находится в центре внимания этой статьи.
Характеристики
Зависимая от NMDA-рецептора LTP проявляет несколько свойств, включая специфичность ввода, ассоциативность, кооперативность и устойчивость.
- Специфика ввода
- После индукции ДП в одном синапсе не распространяется на другие синапсы; скорее LTP зависит от ввода . Долговременная потенциация распространяется на эти синапсы только в соответствии с правилами ассоциативности и кооперативности. Однако входная специфичность LTP может быть неполной на коротких расстояниях. [ нужна ссылка ] Одна модель, объясняющая входную специфичность LTP, была представлена Фреем и Моррисом в 1997 году и называется гипотезой синаптического мечения и захвата . [ 28 ]
- Ассоциативность
- Ассоциативность относится к наблюдению, что, когда слабая стимуляция одного пути недостаточна для индукции ДП, одновременная сильная стимуляция другого пути будет вызывать ДП в обоих путях. [ 29 ]
- Кооперативность
- ДП может быть вызвана либо сильной тетанической стимуляцией одного пути к синапсу, либо совместно посредством более слабой стимуляции многих. Когда один путь в синапс стимулируется слабо, он вызывает недостаточную постсинаптическую деполяризацию для индукции ДП. Напротив, когда слабые стимулы применяются ко многим путям, которые сходятся на одном участке постсинаптической мембраны, отдельные генерируемые постсинаптические деполяризации могут коллективно деполяризовать постсинаптическую клетку в достаточной степени, чтобы совместно индуцировать ДП. Синаптическая маркировка, обсуждаемая позже, может быть распространенным механизмом, лежащим в основе ассоциативности и кооперативности. Брюс Макнотон утверждает, что любое различие между ассоциативностью и кооперативностью строго семантическое. [ 30 ] Эксперименты, проведенные путем стимуляции множества отдельных дендритных шипов, показали, что синаптическая кооперативность всего лишь двух соседних дендритных шипов предотвращает долговременную депрессию (LTD), позволяя использовать только LTP. [ 31 ]
- Упорство
- ДП носит постоянный характер и длится от нескольких минут до многих месяцев, и именно эта устойчивость отличает ДП от других форм синаптической пластичности . [ 32 ]
Ранняя фаза
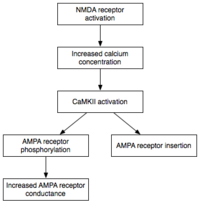

Обслуживание
В то время как индукция влечет за собой временную активацию CaMKII и PKC , поддержание E-LTP (ранней формы LTP) характеризуется их постоянной активацией. На этом этапе ПКМзета (ПКМζ), не зависящая от кальция, становится автономно активной. Следовательно, они способны осуществлять события фосфорилирования, которые лежат в основе экспрессии E-LTP. [ 34 ]
Выражение
Фосфорилирование — это химическая реакция, в которой небольшая фосфатная группа добавляется к другой молекуле, чтобы изменить активность этой молекулы. Автономно активные CaMKII и PKC используют фосфорилирование для реализации двух основных механизмов, лежащих в основе экспрессии E-LTP. Во-первых, и это наиболее важно, они фосфорилируют существующие АМРА-рецепторы, повышая их активность. [ 21 ] Во-вторых, они опосредуют или модулируют вставку дополнительных АМРА-рецепторов в постсинаптическую мембрану. [ 21 ] Важно отметить, что доставка АМРА-рецепторов в синапс во время E-LTP не зависит от синтеза белка . Это достигается за счет наличия несинаптического пула АМРА-рецепторов, прилегающего к постсинаптической мембране. Когда поступает соответствующий стимул, индуцирующий LTP, несинаптические АМРА-рецепторы быстро перемещаются в постсинаптическую мембрану под влиянием протеинкиназ. [ 35 ] Как упоминалось ранее, АМРА-рецепторы являются наиболее распространенными глутаматными рецепторами мозга и опосредуют большую часть его возбуждающей активности. Увеличивая эффективность и количество АМРА-рецепторов в синапсе, будущие возбуждающие стимулы вызывают более сильные постсинаптические реакции.
Хотя приведенная выше модель E-LTP полностью описывает постсинаптические механизмы индукции, поддержания и экспрессии, дополнительный компонент экспрессии может возникать пресинаптически. [ 36 ] Одна из гипотез этого пресинаптического облегчения состоит в том, что постоянная активность CaMKII в постсинаптической клетке во время E-LTP может привести к синтезу «ретроградного мессенджера», который обсуждается позже. Согласно этой гипотезе, вновь синтезированный мессенджер перемещается через синаптическую щель от постсинаптической клетки к пресинаптической, что приводит к цепочке событий, которые облегчают пресинаптический ответ на последующие стимулы. Такие события могут включать увеличение количества везикул нейромедиатора, вероятность высвобождения везикул или и то, и другое. В дополнение к ретроградному мессенджеру, лежащему в основе пресинаптической экспрессии ранней ДП , ретроградный мессенджер может также играть роль в экспрессии поздней ДП.
Поздняя фаза
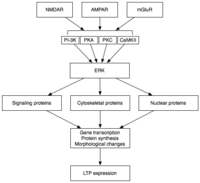
Поздняя LTP (L-LTP) является естественным продолжением E-LTP. В отличие от E-LTP, который не зависит от синтеза белка, L-LTP требует транскрипции генов. [ 37 ] и синтез белка [ 38 ] в постсинаптической клетке. Существуют две фазы L-LTP: первая зависит от синтеза белка, а вторая зависит как от транскрипции гена, так и от синтеза белка. [ 33 ] Эти фазы иногда называют LTP2 и LTP3 соответственно, а E-LTP в этой номенклатуре обозначается как LTP1.
Индукция
Поздняя LTP индуцируется изменениями в экспрессии генов и синтезе белка, вызванными постоянной активацией протеинкиназ, активированных во время E-LTP, таких как MAPK. [ 33 ] [ 34 ] [ 39 ] Фактически, MAPK — в частности, подсемейство MAPK , регулируемых внеклеточными сигнальными киназами (ERK), — может быть молекулярным связующим звеном между E-LTP и L-LTP, поскольку многие сигнальные каскады, участвующие в E-LTP, включая CaMKII и PKC, могут конвергентироваться. на ЭРК. [ 39 ] Недавние исследования показали, что индукция L-LTP может зависеть от совпадающих молекулярных событий, а именно от активации PKA и притока кальция, которые сходятся на CRTC1 (TORC1), мощном коактиваторе транскрипции белка, связывающего элемент ответа цАМФ (CREB). [ 40 ] Это требование молекулярного совпадения прекрасно объясняет ассоциативную природу LTP и, предположительно, природы обучения.
Обслуживание
После активации ERK может фосфорилировать ряд цитоплазматических и ядерных молекул, что в конечном итоге приводит к синтезу белка и морфологическим изменениям, наблюдаемым в L-LTP. [ 33 ] Эти цитоплазматические и ядерные молекулы могут включать факторы транскрипции, такие как CREB. [ 34 ] ERK-опосредованные изменения активности транскрипционных факторов могут запускать синтез белков, которые лежат в основе поддержания L-LTP. Одной из таких молекул может быть протеинкиназа Mζ (PKMζ), постоянно активная киназа, синтез которой увеличивается после индукции LTP. [ 41 ] [ 42 ] PKMζ представляет собой атипичную изоформу PKC, которая лишена регуляторной субъединицы и поэтому остается конститутивно активной. [ 41 ] В отличие от других киназ, которые опосредуют ДП, PKMζ активен не только в течение первых 30 минут после индукции ДП; скорее, PKMζ становится требованием для поддержания LTP только на поздней стадии LTP. [ 41 ] Таким образом, PKMζ важен для сохранения памяти и, как ожидается, будет важен для поддержания долговременной памяти . Действительно, введение ингибитора PKMζ в гиппокамп крысы приводит к ретроградной амнезии с неповрежденной кратковременной памятью ; PKMζ не играет роли в формировании кратковременной памяти. [ 42 ] Недавно было показано, что PKMζ лежит в основе поддержания L-LTP. [ 41 ] [ 42 ] направляя транспортировку и реорганизацию белков в синаптическом каркасе, которые лежат в основе экспрессии L-LTP. [ 41 ] Совсем недавно трансгенные мыши, лишенные PKMζ, демонстрируют нормальную LTP, что ставит под сомнение необходимость PKMζ. [ 43 ]
Долгосрочная стабилизация синаптических изменений определяется также параллельным увеличением пре- и постсинаптических структур, таких как аксональный бутон , дендритный шип и постсинаптическая плотность . [ 44 ] Было показано , что на молекулярном уровне увеличение постсинаптических каркасных белков PSD-95 и Homer1c коррелирует со стабилизацией синаптического расширения. [ 44 ]
Выражение
Известна идентичность лишь нескольких белков, синтезируемых во время L-LTP. Независимо от их личности, считается, что они способствуют увеличению количества дендритных шипов , площади поверхности и постсинаптической чувствительности к нейромедиаторам, связанным с экспрессией L-LTP. [ 33 ] Последнее может быть частично вызвано усиленным синтезом АМРА-рецепторов во время L-LTP. [ 33 ] Поздняя ДП также связана с пресинаптическим синтезом синаптотагмина и увеличением количества синаптических пузырьков , что позволяет предположить, что L-ДП индуцирует синтез белка не только в постсинаптических клетках, но и в пресинаптических клетках. [ 33 ] Как упоминалось ранее, для того, чтобы постсинаптическая индукция LTP привела к пресинаптическому синтезу белка, должна существовать связь между постсинаптической клеткой и пресинаптической клеткой. Это может произойти посредством синтеза ретроградного посланника, о котором речь пойдет позже.
Даже в исследованиях, посвященных постсинаптическим событиям, исследователи не определили место синтеза белка, лежащего в основе L-LTP. В частности, неясно, происходит ли синтез белка в теле постсинаптической клетки или в ее дендритах . [ 39 ] были обнаружены в дендритах еще в 1960-х годах, преобладало мнение, что тело клетки является преобладающим местом синтеза белка в нейронах. Несмотря на то, что рибосомы (основные компоненты механизма синтеза белка) [ 39 ] Это рассуждение не подвергалось серьезному сомнению до 1980-х годов, когда исследователи сообщили о наблюдении синтеза белка в дендритах, связь которых с телом клетки была разорвана. [ 39 ] Совсем недавно исследователи продемонстрировали, что этот тип локального синтеза белка необходим для некоторых типов LTP. [ 45 ] [ 46 ]
Одной из причин популярности гипотезы локального синтеза белка является то, что она обеспечивает возможный механизм специфичности, связанной с LTP. [ 39 ] В частности, если действительно локальный синтез белка лежит в основе L-LTP, только дендритные шипики, получающие стимулы, индуцирующие LTP, будут подвергаться LTP; потенцирование не будет распространяться на соседние синапсы. Напротив, глобальный синтез белка, который происходит в теле клетки, требует, чтобы белки доставлялись во все области клетки, включая синапсы, которые не получили стимулов, индуцирующих LTP. В то время как локальный синтез белка обеспечивает механизм специфичности, глобальный синтез белка, по-видимому, напрямую его ставит под угрозу. Однако, как обсуждается позже, гипотеза синаптического мечения успешно согласовывает глобальный синтез белка, специфичность синапсов и ассоциативность.
Ретроградная сигнализация
Ретроградная передача сигналов — это гипотеза, которая пытается объяснить, что, хотя ДП индуцируется и выражается постсинаптически, некоторые данные свидетельствуют о том, что она выражается также и пресинаптически. [ 21 ] [ 36 ] [ 47 ] Гипотеза получила свое название потому, что нормальная синаптическая передача носит направленный характер и идет от пресинаптической клетки к постсинаптической. Чтобы индукция происходила постсинаптически и частично выражалась пресинаптически, сообщение должно пройти от постсинаптической клетки к пресинаптической в ретроградном (обратном) направлении. Оказавшись там, сообщение, по-видимому, инициирует каскад событий, который приводит к пресинаптическому компоненту экспрессии, например, к увеличению вероятности высвобождения везикул нейромедиатора . [ 48 ]
Ретроградная передача сигналов в настоящее время является спорным вопросом, поскольку некоторые исследователи не считают, что пресинаптические клетки вообще способствуют экспрессии LTP. [ 21 ] Даже среди сторонников гипотезы существуют разногласия по поводу личности посланника. Первые мысли были сосредоточены на оксиде азота , тогда как самые последние данные указывают на клеточной адгезии . белки [ 21 ]
Синаптическая маркировка
До того, как гипотеза локального синтеза белка получила значительную поддержку, существовало общее мнение, что синтез белка, лежащий в основе L-LTP, происходит в теле клетки. Кроме того, считалось, что продукты этого синтеза неспецифически доставляются по всей клетке. Таким образом, возникла необходимость объяснить, как синтез белка может происходить в теле клетки без ущерба для входной специфичности LTP. Гипотеза синаптической маркировки пытается решить сложную проблему клетки, связанную с синтезом белков в теле клетки, но гарантируя, что они достигают только тех синапсов, которые получили стимулы, индуцирующие LTP.
Гипотеза синаптической метки предполагает, что «синаптическая метка» синтезируется в синапсах, которые получили стимулы, индуцирующие LTP, и что эта синаптическая метка может служить для захвата белков, связанных с пластичностью, доставляемых по всей клетке из тела клетки. [ 49 ] Исследования LTP у морской улитки Aplysia Californica показали, что синаптическая маркировка является механизмом входной специфичности LTP. [ 50 ] [ 51 ] Есть некоторые доказательства того, что при наличии двух широко разделенных синапсов стимул, индуцирующий LTP в одном синапсе, запускает несколько сигнальных каскадов (описанных ранее), которые инициируют экспрессию генов в ядре клетки. В том же самом синапсе (но не в нестимулированном синапсе) локальный синтез белка создает короткоживущую (менее трех часов) синаптическую метку. Продукты экспрессии генов разносятся по всей клетке, но захватываются только синапсами, экспрессирующими синаптические метки. Таким образом, потенцируется только тот синапс, который получает стимулы, индуцирующие ДП, что демонстрирует специфичность входного сигнала ДП.
Гипотеза синаптических тегов также может объяснять ассоциативность и кооперативность LTP. Ассоциативность ( см . «Свойства ») наблюдается, когда один синапс возбуждается стимуляцией, индуцирующей LTP, в то время как отдельный синапс стимулируется лишь слабо. Хотя можно было бы ожидать, что только сильно стимулированный синапс подвергнется ДП (поскольку одной лишь слабой стимуляции недостаточно для индукции ДП в любом из синапсов), оба на самом деле ДП будут подвергаться синапса. Хотя слабые стимулы не способны индуцировать синтез белка в теле клетки, они могут стимулировать синтез синаптической метки. Одновременная сильная стимуляция отдельного пути, способного индуцировать синтез белка клеточного тела, может затем вызвать выработку белков, связанных с пластичностью, которые доставляются по всей клетке. Поскольку оба синапса экспрессируют синаптическую метку, оба будут захватывать белковые продукты, что приводит к экспрессии LTP как в сильно стимулируемых, так и в слабостимулируемых путях.
Кооперативность наблюдается, когда два синапса активируются слабыми стимулами, неспособными вызвать ДП при индивидуальной стимуляции. Но при одновременной слабой стимуляции оба синапса совместно подвергаются ДП. Синаптическое мечение не объясняет, как множественные слабые стимулы могут привести к образованию коллективного стимула, достаточного для индукции ДП (это объясняется постсинаптическим суммированием ВПСП, описанным ранее). Скорее, синаптическое мечение объясняет способность слабостимулированных синапсов, ни один из которых не способен самостоятельно генерировать ДП, получать продукты синтеза белка, инициируемого коллективно. Как и раньше, это может быть достигнуто посредством синтеза локальной синаптической метки после слабой синаптической стимуляции.
Модуляция
| Модулятор | Цель |
|---|---|
| β-адренергический рецептор | амплификация цАМФ, МАРК |
| Синтаза оксида азота | Гуанилилциклаза, ПКГ, NMDAR |
| Дофаминовый рецептор | амплификация цАМФ, МАРК |
| Метаботропный глутаматный рецептор | ПКС, МАПК усиление |
Как описано ранее, молекулы, лежащие в основе LTP, можно классифицировать как медиаторы или модуляторы. Медиатором LTP является молекула, такая как рецептор NMDA или кальций, присутствие и активность которого необходимы для генерации LTP практически при всех условиях. Напротив, модулятор — это молекула, которая может изменять ДП, но не является необходимой для его генерации или экспрессии. [ 21 ]
Помимо описанных выше сигнальных путей, LTP гиппокампа может быть изменен различными модуляторами. Например, стероидный гормон эстрадиол может усиливать LTP, стимулируя фосфорилирование CREB и последующий рост дендритных шипов . [ 52 ] Кроме того, агонисты β-адренергических рецепторов, такие как норадреналин, могут изменять зависимую от синтеза белка позднюю фазу ДП. [ 53 ] Активность синтазы оксида азота также может приводить к последующей активации гуанилилциклазы и ПКГ. [ 54 ] Аналогично, активация дофаминовых рецепторов может усиливать ДП посредством сигнального пути цАМФ/ПКА. [ 55 ] [ 56 ]
Связь с поведенческой памятью
Хотя долговременное усиление синапсов в клеточной культуре, по-видимому, обеспечивает элегантный субстрат для обучения и памяти, вклад ДП в поведенческое обучение, то есть обучение на уровне всего организма, нельзя просто экстраполировать из in vitro исследований . . По этой причине значительные усилия были направлены на установление того, является ли LTP необходимым условием обучения и памяти у живых животных. По этой причине LTP также играет решающую роль в обработке страха .
Пространственная память

В 1986 году Ричард Моррис предоставил одни из первых доказательств того, что ДП действительно необходима для формирования воспоминаний in vivo . [ 57 ] Он проверил пространственную память крыс, фармакологически модифицировав их гиппокамп — структуру мозга, роль которой в пространственном обучении хорошо известна. Крыс обучали водному лабиринту Морриса — задаче на пространственную память, в которой крысы плавают в луже с мутной водой, пока не находят платформу, скрытую под ее поверхностью. Ожидается, что во время этого упражнения нормальные крысы будут ассоциировать расположение скрытой платформы с заметными сигналами, расположенными в определенных местах по окружности лабиринта. После тренировки гиппокамп одной группы крыс окунали в блокатор рецепторов NMDA APV , а другая группа служила контролем . Обе группы затем были подвергнуты задаче на пространственную память в водном лабиринте. Крысы в контрольной группе смогли найти платформу и выбраться из бассейна, в то время как работоспособность крыс, получавших APV, была значительно снижена. Более того, когда у обеих групп были взяты срезы гиппокампа, LTP легко индуцировался в контрольной группе, но не мог индуцироваться в мозге крыс, обработанных APV. Это дало ранние доказательства того, что рецептор NMDA — и, как следствие, LTP — необходим, по крайней мере, для некоторых типов обучения и памяти.
Точно так же Сусуму Тонегава продемонстрировал в 1996 году, что область CA1 гиппокампа имеет решающее значение для формирования пространственной памяти у живых мышей. [ 58 ] Так называемые клетки места , расположенные в этой области, становятся активными только тогда, когда крыса находится в определенном месте — называемом полем места — в окружающей среде. Поскольку эти поля мест распределены по всей окружающей среде, одна из интерпретаций состоит в том, что группы клеток мест образуют карты в гиппокампе. Точность этих карт определяет, насколько хорошо крыса изучает окружающую среду и, следовательно, насколько хорошо она может в ней ориентироваться. Тонегава обнаружил, что при повреждении рецептора NMDA, в частности, путем генетического удаления субъединицы NR1 в регионе CA1, генерируемые поля места были существенно менее специфичными, чем у контрольной группы. То есть мыши создавали ошибочные пространственные карты, когда их NMDA-рецепторы были повреждены. Как и ожидалось, эти мыши очень плохо справлялись с пространственными задачами по сравнению с контрольной группой, что еще раз подтверждает роль LTP в пространственном обучении.
Также было показано, что повышенная активность рецепторов NMDA в гиппокампе приводит к усилению ДП и общему улучшению пространственного обучения. В 1999 году Тан и др . создали линию мышей с усиленной функцией рецептора NMDA за счет сверхэкспрессии субъединицы NR2B в гиппокампе. [ 59 ] [ 60 ] Получившиеся в результате умные мыши, получившие прозвище «мыши Дуги» в честь вымышленного выдающегося доктора Дуги Хаузера , имели более крупный LTP и превосходно справлялись с задачами пространственного обучения, что еще раз подчеркивало важность LTP в формировании памяти, зависящей от гиппокампа.
Тормозное избегание
В 2006 году Джонатан Уитлок и его коллеги сообщили о серии экспериментов, которые предоставили, возможно, самые убедительные доказательства роли LTP в поведенческой памяти, утверждая, что для того, чтобы сделать вывод о том, что LTP лежит в основе поведенческого обучения, эти два процесса должны одновременно имитировать и перекрывать друг друга. [ 61 ] Используя парадигму обучения тормозящему избеганию, исследователи обучали крыс в двухкамерном аппарате со светлой и темной камерами, причем последняя была оснащена устройством, которое при входе в нее наносило удар по ноге крысы. Анализ синапсов гиппокампа CA1 показал, что тренировка тормозного избегания индуцирует in vivo фосфорилирование рецептора AMPA того же типа, что и наблюдаемое при LTP in vitro ; то есть тренировка тормозного избегания имитировала LTP. Кроме того, синапсы, потенцируемые во время обучения, не могли быть дополнительно усилены экспериментальными манипуляциями, которые в противном случае вызвали бы ДП; то есть тренировка тормозного избегания блокировала LTP. В ответ на статью Тимоти Блисс и его коллеги отметили, что эти и подобные эксперименты «существенно подтверждают теорию ДП как нейронного механизма памяти». [ 62 ]
Клиническое значение
Роль LTP при заболевании менее ясна, чем ее роль в основных механизмах синаптической пластичности . Однако изменения в ДП могут способствовать ряду неврологических заболеваний , включая депрессию , болезнь Паркинсона , эпилепсию и нейропатическую боль . [ 63 ] Нарушение LTP также может играть роль в развитии болезни Альцгеймера и наркозависимости .
болезнь Альцгеймера

ЛТП привлекла большое внимание среди тех, кто изучает болезнь Альцгеймера (БА), нейродегенеративное заболевание , вызывающее выраженное снижение когнитивных функций и деменцию . Большая часть этого ухудшения происходит в связи с дегенеративными изменениями в гиппокампе и других медиальных структурах височной доли . Из-за хорошо известной роли гиппокампа в LTP некоторые предполагают, что снижение когнитивных функций, наблюдаемое у людей с AD, может быть результатом нарушения LTP.
В обзоре литературы 2003 г. Rowan et al. предложил одну модель того, как LTP может повлиять на AD. [ 64 ] БА, по-видимому, является результатом, по крайней мере частично, неправильной обработки белка-предшественника амилоида (АРР). Результатом этой аномальной обработки является накопление фрагментов этого белка, называемого амилоидом β (Aβ). Aβ существует как в растворимой, так и в фибриллярной формах. Неправильная обработка APP приводит к накоплению растворимого Aβ, который, согласно гипотезе Роуэна, нарушает LTP гиппокампа и может привести к снижению когнитивных функций, наблюдаемому на ранних стадиях болезни Альцгеймера.
AD может также нарушать LTP посредством механизмов, отличных от Aβ. Например, одно исследование показало, что фермент PKMζ накапливается в нейрофибриллярных клубках , которые являются патологическим маркером БА. PKMζ представляет собой фермент, имеющий решающее значение для поддержания поздней LTP . [ 65 ]
Наркомания
исследования в области медицины наркологии. Недавно были проведены [ когда? ] сосредоточил свое внимание на LTP из-за гипотезы о том, что наркомания представляет собой мощную форму обучения и памяти. [ 66 ] Зависимость — это сложное нейроповеденческое явление, затрагивающее различные части мозга, такие как вентральная покрышка (VTA) и прилежащее ядро (NAc). Исследования показали, что синапсы VTA и NAc способны подвергаться LTP. [ 66 ] и что этот LTP может быть ответственным за поведение, характеризующее зависимость. [ 67 ]
См. также
- Нейропластичность
- Актиновое ремоделирование нейронов
- Транскраниальная стимуляция постоянным током
- Посттетаническая потенциация
Ссылки
- ^ Парадизо М.А., Медведь М.Ф., Коннорс Б.В. (2007). Нейронаука: исследование мозга . Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. п. 718 . ISBN 978-0-7817-6003-4 .
- ^ Jump up to: а б Кук С.Ф., Bliss TV (июль 2006 г.). «Пластичность центральной нервной системы человека» . Мозг . 129 (Часть 7): 1659–73. дои : 10.1093/brain/awl082 . ПМИД 16672292 .
- ^ Jump up to: а б Bliss TV, Collingridge GL (январь 1993 г.). «Синаптическая модель памяти: долговременная потенциация в гиппокампе». Природа . 361 (6407): 31–9. Бибкод : 1993Natur.361...31B . дои : 10.1038/361031a0 . ПМИД 8421494 . S2CID 4326182 .
- ^ Уильямс Р.В., Херруп К. (1988). «Контроль количества нейронов». Ежегодный обзор неврологии . 11 (1): 423–53. дои : 10.1146/annurev.ne.11.030188.002231 . ПМИД 3284447 .
- ^ Jump up to: а б Сантьяго RC (1894 г.). «Крунская лекция: Тонкая структура нервных центров» . Труды Лондонского королевского общества . 55 (331–335): 444–468. Бибкод : 1894RSPS...55..444C . дои : 10.1098/rspl.1894.0063 .
- ^ Хебб Д. (1949). Организация поведения: НЕЙРОПСИХОЛОГИЧЕСКАЯ ТЕОРИЯ . Нью-Йорк: ДЖОН УАЙЛИ, если SONS, Inc. ISBN 978-0805843002 .
- ^ Хебб Д.О. (1949). Организация поведения: нейропсихологическая теория . Нью-Йорк: Джон Уайли. ISBN 978-0-471-36727-7 .
- ^ Кандель Э.Р., Таук Л. (ноябрь 1965 г.). «Гетеросинапсическое облегчение в нейронах брюшного ганглия Aplysia depilans» . Журнал физиологии . 181 (1): 1–27. doi : 10.1113/jphysicalol.1965.sp007742 . ПМЦ 1357435 . ПМИД 5866283 .
- ^ Патихис Л. (октябрь 2018 г.). «Историческое значение открытия долговременного потенцирования: обзор и оценка для неспециалистов» . Американский журнал психологии . 131 (3): 369–80. дои : 10.5406/amerjpsyc.131.3.0369 .
- ^ Jump up to: а б Лёмо Т. (апрель 2003 г.). «Открытие долговременного потенцирования» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 617–20. дои : 10.1098/rstb.2002.1226 . ПМК 1693150 . ПМИД 12740104 .
- ^ Лёмо Т. (1966). «Потенцирование частоты возбуждающей синаптической активности в зубчатой области гиппокампальной формации». Acta Physiologica Scandinavica . 68 (Приложение 277): 128.
- ^ Jump up to: а б Блисс ТВ, Ломо Т (июль 1973 г.). «Длительное усиление синаптической передачи в зубчатой области анестезированного кролика после стимуляции перфорантного пути» . Журнал физиологии . 232 (2): 331–56. doi : 10.1113/jphysicalol.1973.sp010273 . ПМК 1350458 . ПМИД 4727084 .
- ^ Jump up to: а б Bliss TV, Гарднер-Медвин, Арканзас (июль 1973 г.). «Длительная потенциация синаптической передачи в зубчатой области неанестезированного кролика после стимуляции перфорантного пути» . Журнал физиологии . 232 (2): 357–74. doi : 10.1113/jphysicalol.1973.sp010274 . ПМЦ 1350459 . ПМИД 4727085 .
- ↑ Хотя термин «долговременное потенцирование» однажды появился в оригинальной статье Блисс и Лёмо, официально он не был предложен для этого явления до статьи Дугласа и Годдарда.
- ^ Дуглас Р.М., Годдард Г.В. (март 1975 г.). «Длительная потенциация синапса перфорантных клеток-зерен в гиппокампе крысы». Исследования мозга . 86 (2): 205–15. дои : 10.1016/0006-8993(75)90697-6 . ПМИД 163667 . S2CID 43260928 .
- ^ Андерсен П. (апрель 2003 г.). «Прелюдия к долговременному потенцированию» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 613–5. дои : 10.1098/rstb.2002.1232 . ПМЦ 1693144 . ПМИД 12740103 .
- ^ Макихерн Дж. К., Шоу, Калифорния (июнь 1996 г.). «Альтернатива ортодоксальности LTP: модель континуума пластичности-патологии». Исследования мозга. Обзоры исследований мозга . 22 (1): 51–92. дои : 10.1016/0165-0173(96)00006-9 . ПМИД 8871785 . S2CID 41680613 . 8871785.
- ^ Медведь МФ (1996). «Синаптическая основа хранения памяти в коре головного мозга» . Труды Национальной академии наук . 93 (24): 13453–13459. Бибкод : 1996PNAS...9313453B . дои : 10.1073/pnas.93.24.13453 . ПМК 33630 . ПМИД 8942956 .
- ^ Уардуз М., Састри Б.Р. (2000). «Механизмы, лежащие в основе LTP тормозной синаптической передачи в глубоких ядрах мозжечка». Журнал нейрофизиологии . 84 (3): 1414–1421. дои : 10.1152/jn.2000.84.3.1414 . ПМИД 10980014 . S2CID 16972473 .
- ^ Clugnet MC, LeDoux JE (август 1990 г.). «Синаптическая пластичность в цепях кондиционирования страха: индукция ДП в латеральном ядре миндалевидного тела путем стимуляции медиального коленчатого тела» . Журнал неврологии . 10 (8): 2818–24. doi : 10.1523/JNEUROSCI.10-08-02818.1990 . ПМК 6570263 . ПМИД 2388089 .
- ^ Jump up to: а б с д и ж г час я Маленка Р.К., Медведь М.Ф. (сентябрь 2004 г.). «ЛТП и ЛТД: конфуз богатства» . Нейрон . 44 (1): 5–21. дои : 10.1016/j.neuron.2004.09.012 . ПМИД 15450156 . S2CID 79844 .
- ^ Ясуда Х., Барт А.Л., Стеллваген Д., Маленка Р.С. (январь 2003 г.). «Переключатель развития в сигнальных каскадах для индукции LTP». Природная неврология . 6 (1): 15–6. дои : 10.1038/nn985 . ПМИД 12469130 . S2CID 28913342 .
- ^ Коллингридж Г.Л., Кель С.Дж., МакЛеннан Х. (январь 1983 г.). «Возбуждающие аминокислоты в синаптической передаче в коллатерально-комиссуральном пути Шаффера гиппокампа крысы» . Журнал физиологии . 334 : 33–46. дои : 10.1113/jphysicalol.1983.sp014478 . ПМЦ 1197298 . ПМИД 6306230 .
- ^ Харрис EW, Котман CW (сентябрь 1986 г.). «Долгосрочное усиление реакции мшистых волокон морской свинки не блокируется антагонистами N-метил-D-аспартата». Письма по неврологии . 70 (1): 132–7. дои : 10.1016/0304-3940(86)90451-9 . ПМИД 3022192 . S2CID 42647125 .
- ^ Вигстрём Х, Густафссон Б (1986). «Постсинаптический контроль долговременной потенциации гиппокампа». Журнал физиологии . 81 (4): 228–36. ПМИД 2883309 .
- ^ Урбан НН, Баррионуево Г (июль 1996 г.). «Индукция долговременного потенциирования хеббианских и нехеббианских мшистых волокон с помощью различных моделей высокочастотной стимуляции» . Журнал неврологии . 16 (13): 4293–9. doi : 10.1523/JNEUROSCI.16-13-04293.1996 . ПМК 6579001 . ПМИД 8753890 .
- ^ Куллманн Д.М., Ламса К. (март 2008 г.). «Роль отдельных рецепторов глутамата в индукции долговременного анти-Хеббианского потенцирования» . Журнал физиологии . 586 (6): 1481–6. дои : 10.1113/jphysicalol.2007.148064 . ПМЦ 2375711 . ПМИД 18187472 .
- ^ Фрей Ю, Моррис Р.Г. (февраль 1997 г.). «Синаптическая маркировка и долгосрочная потенциация». Природа . 385 (6616): 533–536. Бибкод : 1997Natur.385..533F . дои : 10.1038/385533a0 . ПМИД 9020359 . S2CID 4339789 .
- ^ Хао Л., Ян З., Лэй Дж. (1 мая 2018 г.). «Основные механизмы кооперативности, входной специфичности и ассоциативности долгосрочного потенциирования посредством положительной обратной связи локального синтеза белка» . Границы вычислительной нейронауки . 12:25 . дои : 10.3389/fncom.2018.00025 . ПМЦ 5938377 . ПМИД 29765314 .
- ^ Макнотон Б.Л. (апрель 2003 г.). «Долгосрочная потенциация, кооперативность и клеточные сборки Хебба: личная история» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 629–34. дои : 10.1098/rstb.2002.1231 . ПМЦ 1693161 . ПМИД 12740107 .
- ^ Тазерарт С., Митчелл Д.Э., Миранда-Роттманн С., Арайя Р. (август 2020 г.). «Правило пластичности, зависящее от времени всплеска, для дендритных шипов» . Природные коммуникации . 11 (1): 4276. Бибкод : 2020NatCo..11.4276T . дои : 10.1038/s41467-020-17861-7 . ПМЦ 7449969 . ПМИД 32848151 .
- ^ Авраам В.К. (апрель 2003 г.). «Как долго будет длиться долговременное потенцирование?» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 735–44. дои : 10.1098/rstb.2002.1222 . ПМЦ 1693170 . ПМИД 12740120 .
- ^ Jump up to: а б с д и ж г час Линч М.А. (январь 2004 г.). «Долговременная потенциация и память» . Физиологические обзоры . 84 (1): 87–136. doi : 10.1152/physrev.00014.2003 . ПМИД 14715912 .
- ^ Jump up to: а б с д Суэтт Джей Ди (1999). «К молекулярному объяснению долговременного потенциирования» . Обучение и память . 6 (5): 399–416. дои : 10.1101/lm.6.5.399 . ПМИД 10541462 .
- ^ Малинов Р. (апрель 2003 г.). «Торговля рецепторами AMPA и долгосрочное потенцирование» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 707–14. дои : 10.1098/rstb.2002.1233 . ПМЦ 1693162 . ПМИД 12740116 .
- ^ Jump up to: а б Эмптадж, Нью-Джерси, Рид, Калифорния, Fine A, Bliss TV (июнь 2003 г.). «Оптически-квантовый анализ выявляет пресинаптический компонент LTP в гиппокампальных синапсах Шаффера» . Нейрон . 38 (5): 797–804. дои : 10.1016/S0896-6273(03)00325-8 . ПМИД 12797963 . S2CID 13629691 .
- ^ Фрей У, Фрей С, Шольмайер Ф, Круг М (февраль 1996 г.). «Влияние актиномицина D, ингибитора синтеза РНК, на долговременную потенциацию нейронов гиппокампа крысы in vivo и in vitro» . Журнал физиологии . 490. 490 (Часть 3): 703–11. doi : 10.1113/jphysicalol.1996.sp021179 . ПМЦ 1158708 . ПМИД 8683469 .
- ^ Фрей У., Круг М., Рейманн К.Г., Мэттис Х. (июнь 1988 г.). «Анизомицин, ингибитор синтеза белка, блокирует поздние фазы явлений LTP в области CA1 гиппокампа in vitro». Исследования мозга . 452 (1–2): 57–65. дои : 10.1016/0006-8993(88)90008-X . ПМИД 3401749 . S2CID 39245231 .
- ^ Jump up to: а б с д и ж Келлехер Р.Дж., Говиндараджан А., Тонегава С. (сентябрь 2004 г.). «Механизмы трансляционной регуляции в стойких формах синаптической пластичности» . Нейрон . 44 (1): 59–73. дои : 10.1016/j.neuron.2004.09.013 . ПМИД 15450160 . S2CID 1511103 .
- ^ Ковач К.А., Стеуллет П., Стейнманн М., До К.К., Маджистретти П.Дж., Халфон О. и др. (март 2007 г.). «TORC1 представляет собой чувствительный к кальцию и цАМФ детектор совпадений, участвующий в долгосрочной синаптической пластичности гиппокампа» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (11): 4700–5. Бибкод : 2007PNAS..104.4700K . дои : 10.1073/pnas.0607524104 . ПМЦ 1838663 . ПМИД 17360587 .
- ^ Jump up to: а б с д и Серрано П., Яо Й., Сактор Т.С. (февраль 2005 г.). «Постоянное фосфорилирование протеинкиназой Mzeta поддерживает долговременное потенцирование поздней фазы» . Журнал неврологии . 25 (8): 1979–84. doi : 10.1523/JNEUROSCI.5132-04.2005 . ПМК 6726070 . ПМИД 15728837 .
- ^ Jump up to: а б с Пасталькова Е., Серрано П., Пинхасова Д., Уоллес Э., Фентон А.А., Сактор Т.С. (август 2006 г.). «Хранение пространственной информации с помощью механизма обслуживания LTP». Наука . 313 (5790): 1141–4. Бибкод : 2006Sci...313.1141P . CiteSeerX 10.1.1.453.2136 . дои : 10.1126/science.1128657 . ПМИД 16931766 . S2CID 7260010 .
- ^ Волк Л.Дж., Бахман Дж.Л., Джонсон Р., Ю Ю, Хуганир Р.Л. (январь 2013 г.). «PKM-ζ не требуется для синаптической пластичности гиппокампа, обучения и памяти» . Природа . 493 (7432): 420–3. Бибкод : 2013Природа.493..420В . дои : 10.1038/nature11802 . ПМЦ 3830948 . ПМИД 23283174 .
- ^ Jump up to: а б Мейер Д., Бонхёффер Т., Шойсс В. (апрель 2014 г.). «Баланс и стабильность синаптических структур при синаптической пластичности» . Нейрон . 82 (2): 430–43. дои : 10.1016/j.neuron.2014.02.031 . ПМИД 24742464 .
- ^ Кан Х, Шуман Э.М. (сентябрь 1996 г.). «Необходимость локального синтеза белка для синаптической пластичности гиппокампа, индуцированной нейротрофинами». Наука . 273 (5280): 1402–6. Бибкод : 1996Sci...273.1402K . дои : 10.1126/science.273.5280.1402 . ПМИД 8703078 . S2CID 38648558 .
- ^ Стюард О, Уорли ПФ (июнь 2001 г.). «Клеточный механизм нацеливания вновь синтезированных мРНК на синаптические участки дендритов» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (13): 7062–8. Бибкод : 2001PNAS...98.7062S . дои : 10.1073/pnas.131146398 . ПМК 34623 . ПМИД 11416188 .
- ^ Павлидис П., Монтгомери Дж. , Мэдисон Д.В. (июнь 2000 г.). «Активность пресинаптической протеинкиназы поддерживает долговременное усиление синапсов между отдельными нейронами гиппокампа» . Журнал неврологии . 20 (12): 4497–505. doi : 10.1523/JNEUROSCI.20-12-04497.2000 . ПМК 6772468 . ПМИД 10844019 .
- ^ Захаренко С.С., Паттерсон С.Л., Драгацис И., Цейтлин С.О., Сигельбаум С.А., Кандель Э.Р. и др. (сентябрь 2003 г.). «Пресинаптический BDNF необходим для пресинаптического, но не постсинаптического компонента LTP в синапсах CA1-CA3 гиппокампа» . Нейрон . 39 (6): 975–90. дои : 10.1016/S0896-6273(03)00543-9 . ПМИД 12971897 . S2CID 7117772 .
- ^ Фрей Ю, Моррис Р.Г. (февраль 1997 г.). «Синаптическая маркировка и долгосрочная потенциация». Природа . 385 (6616): 533–6. Бибкод : 1997Natur.385..533F . дои : 10.1038/385533a0 . ПМИД 9020359 . S2CID 4339789 .
- ^ Мартин К.С., Касадио А., Чжу Х., Япин Э., Роуз Дж.К., Чен М. и др. (декабрь 1997 г.). «Синапс-специфическое, долгосрочное облегчение сенсорной аплизии моторных синапсов: функция локального синтеза белка в хранилищах памяти» . Клетка . 91 (7): 927–38. дои : 10.1016/S0092-8674(00)80484-5 . ПМИД 9428516 . S2CID 16423304 .
- ^ Касадио А., Мартин К.С., Джустетто М., Чжу Х., Чен М., Барч Д. и др. (октябрь 1999 г.). «Переходная, охватывающая весь нейрон форма долгосрочного облегчения, опосредованного CREB, может быть стабилизирована в специфических синапсах за счет локального синтеза белка» . Клетка . 99 (2): 221–37. дои : 10.1016/S0092-8674(00)81653-0 . ПМИД 10535740 .
- ^ Сигал М., Мерфи Д.Д. (1999). «Активация CREB опосредует пластичность культивируемых нейронов гиппокампа» . Нейронная пластичность . 6 (3): 1–7. дои : 10.1155/NP.1998.1 . ПМК 2565317 . ПМИД 9920677 .
- ^ Штраубе Т., Фрей Ю. (2003). «Участие бета-адренергических рецепторов в зависимой от синтеза белка поздней долгосрочной потенциации (ДП) в зубчатой извилине свободно движущихся крыс: решающая роль силы индукции ДП». Нейронаука . 119 (2): 473–9. дои : 10.1016/S0306-4522(03)00151-9 . ПМИД 12770561 . S2CID 23436714 .
- ^ Лу Ю.Ф., Кандел Э.Р., Хокинс Р.Д. (декабрь 1999 г.). «Передача сигналов оксида азота способствует поздней фазе фосфорилирования LTP и CREB в гиппокампе» . Журнал неврологии . 19 (23): 10250–61. doi : 10.1523/JNEUROSCI.19-23-10250.1999 . ПМК 6782403 . ПМИД 10575022 .
- ^ Фрей У., Мэттис Х., Рейманн К.Г., Мэттис Х. (август 1991 г.). «Влияние блокады дофаминергических рецепторов D1 во время тетанизации на проявление долговременной потенциации в области CA1 крысы in vitro». Письма по неврологии . 129 (1): 111–4. дои : 10.1016/0304-3940(91)90732-9 . ПМИД 1833673 . S2CID 45084596 .
- ^ Отмахова Н.А., Лисман Ю.Е. (декабрь 1996 г.). «Активация дофаминовых рецепторов D1/D5 увеличивает величину ранней долгосрочной потенциации в синапсах гиппокампа CA1» . Журнал неврологии . 16 (23): 7478–86. doi : 10.1523/JNEUROSCI.16-23-07478.1996 . ПМК 6579102 . ПМИД 8922403 .
- ^ Моррис Р.Г., Андерсон Э., Линч Г.С., Бодри М. (1986). «Селективное нарушение обучения и блокада долговременного потенциирования антагонистом рецептора N-метил-D-аспартата, AP5». Природа . 319 (6056): 774–6. Бибкод : 1986Natur.319..774M . дои : 10.1038/319774a0 . ПМИД 2869411 . S2CID 4356601 .
- ^ МакХью Т.Дж., Блюм К.И., Цьен Дж.З., Тонегава С., Уилсон М.А. (декабрь 1996 г.). «Нарушение представления пространства в гиппокампе у мышей с нокаутом NMDAR1, специфичных для CA1» . Клетка . 87 (7): 1339–49. дои : 10.1016/S0092-8674(00)81828-0 . ПМИД 8980239 . S2CID 5131226 .
- ^ Тан Ю.П., Симидзу Э., Дубе Г.Р., Рампон С., Кершнер Г.А., Чжуо М. и др. (1999). «Генетическое улучшение обучения и памяти у мышей» . Природа . 401 (6748): 63–69. Бибкод : 1999Natur.401...63T . дои : 10.1038/43432 . ПМИД 10485705 . S2CID 481884 .
- ^ Тан Ю, Ван Х, Фэн Р, Кьин М, Цянь Дж (2001). «Дифференциальное влияние обогащения на функции обучения и памяти у трансгенных мышей NR2B». Нейрофармакология . 41 (6): 779–90. дои : 10.1016/S0028-3908(01)00122-8 . ПМИД 11640933 . S2CID 23602265 .
- ^ Уитлок-младший, Хейнен А.Дж., Шулер М.Г., Медведь М.Ф. (август 2006 г.). «Обучение вызывает долговременную потенциацию в гиппокампе». Наука . 313 (5790): 1093–7. Бибкод : 2006Sci...313.1093W . дои : 10.1126/science.1128134 . ПМИД 16931756 . S2CID 612352 .
- ^ Bliss TV, Коллингридж Г.Л., Ларош С. (август 2006 г.). «Нейронаука. ZAP и ZIP, история, которую стоит забыть». Наука . 313 (5790): 1058–9. дои : 10.1126/science.1132538 . ПМИД 16931746 . S2CID 27735098 .
- ^ Кук С.Ф., Bliss TV (июль 2006 г.). «Пластичность центральной нервной системы человека» . Мозг . 129 (Часть 7): 1659–73. дои : 10.1093/brain/awl082 . ПМИД 16672292 .
- ^ Jump up to: а б Роуэн М.Дж., Клубин И., Каллен В.К., Анвил Р. (апрель 2003 г.). «Синаптическая пластичность на животных моделях ранней болезни Альцгеймера» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1432): 821–8. дои : 10.1098/rstb.2002.1240 . ПМК 1693153 . ПМИД 12740129 .
- ^ Крари Дж.Ф., Шао С.И., Мирра С.С., Эрнандес А.И., Сактор Т.К. (апрель 2006 г.). «Атипичная протеинкиназа C при нейродегенеративном заболевании I: PKMzeta агрегирует с лимбическими нейрофибриллярными клубками и рецепторами AMPA при болезни Альцгеймера». Журнал невропатологии и экспериментальной неврологии . 65 (4): 319–26. дои : 10.1097/01.jnen.0000218442.07664.04 . ПМИД 16691113 .
- ^ Jump up to: а б Кауэр Дж.А., Маленка Р.С. (ноябрь 2007 г.). «Синаптическая пластичность и зависимость» . Обзоры природы. Нейронаука . 8 (11): 844–58. дои : 10.1038/nrn2234 . ПМИД 17948030 . S2CID 38811195 .
- ^ Вольф М.Е. (август 2003 г.). «ЛТП может вызвать зависимость» . Молекулярные вмешательства . 3 (5): 248–52. дои : 10.1124/ми.3.5.248 . ПМИД 14993438 .
Дальнейшее чтение
- Блисс Т., Коллингридж Дж., Моррис Р., ред. (2004). Долгосрочное потенцирование: развитие нейробиологии в течение 30 лет . Издательство Оксфордского университета. дои : 10.1093/oso/9780198530305.001.0001 . ISBN 978-0-19-853030-5 .
- Андерсен П., Моррис Р., Амарал Д., Блисс Т., О'Киф Дж. (2007). «10. Синаптическая пластичность в гиппокампе» . Книга «Гиппокамп» . Издательство Оксфордского университета. стр. 350–474. doi : 10.1093/acprof:oso/9780195100273.003.0010 . ISBN 978-0-19-510027-3 .
Внешние ссылки
- Исследователи представили первые доказательства существования механизма обучения — отчет PhysOrg.com об исследовании Беара и его коллег, проведенном в 2006 году.
- Короткий документальный видеофильм о мышах Дуги. ( формат RealPlayer )
- «Умная мышь», серия телеканала Quantum ABC о мышах Дуги.
- Долгосрочное + потенцирование в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)