Частица распознавания сигнала РНК
| РН7СЛ1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RN7SL1 , 7L1a, 7SL, RN7SL, RNSRP1, частица распознавания сигнала РНК, РНК, 7SL, цитоплазматическая 1, компонент РНК частицы распознавания сигнала 7SL1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 612177 ; Генные карты : RN7SL1 ; ОМА : RN7SL1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||



РНК частицы распознавания сигнала (также известная как 7SL, 6S, ffs или 4.5S РНК) является частью комплекса частицы распознавания сигнала (SRP) рибонуклеопротеинового . SRP распознает сигнальный пептид и связывается с рибосомой , останавливая синтез белка. SRP-рецептор представляет собой белок, встроенный в мембрану и содержащий трансмембранную пору . Когда комплекс SRP-рибосома связывается с SRP-рецептором , SRP высвобождает рибосому и уходит. Рибосома возобновляет синтез белка, но теперь белок движется через трансмембранную пору SRP-рецептора .
Таким образом, SRP направляет движение белков внутри клетки для связывания с трансмембранной порой, что позволяет белку пересекать мембрану туда, где он необходим. РНК и белковые компоненты этого комплекса высококонсервативны , но различаются в разных царствах жизни.
Общее SINE семейство Alu , вероятно, произошло от гена 7SL РНК после делеции центральной последовательности. [3]
Эукариотический и шести белков: SRP 72, 68, 54 SRP состоит из 300-нуклеотидной 7S РНК , 19, 14 и 9. Архейный SRP состоит из 7S РНК и гомологов эукариотических белков SRP19 и SRP54. 7S РНК эукариот и архей имеют очень схожие вторичные структуры. [4]
У большинства бактерий SRP состоит из молекулы РНК (4,5S) и белка Ffh (гомолога эукариотического белка SRP54). Некоторые грамположительные бактерии (например, Bacillus subtilis ) имеют более длинную эукариотоподобную SRP-РНК, которая включает домен Alu . [5]
У эукариот и архей восемь спиральных элементов складываются в домены Alu и S, разделенные длинной линкерной областью. [6] [7] Считается, что домен Alu опосредует функцию замедления удлинения пептидной цепи SRP. [6] Универсально консервативная спираль, которая взаимодействует с М-доменом SRP54, опосредует распознавание сигнальной последовательности. [7] [8] Считается, что комплекс SRP19-спираль 6 участвует в сборке SRP и стабилизирует спираль 8 для SRP54. обязательность [6] У людей есть три функциональных гена SRP РНК, которые удобно называть RN7SL1, RN7SL2 и RN7SL3. В частности, известно, что геном человека содержит большое количество последовательностей, связанных с SRP-РНК, включая повторы Alu . [5]
Открытие
[ редактировать ]РНК SRP была впервые обнаружена в онкогенной РНК (окорны) частицах вируса птиц и мышей . [9] Впоследствии было обнаружено, что SRP РНК является стабильным компонентом неинфицированных клеток HeLa, где она связывается с мембранными и полисомальными фракциями. [10] [11] В 1980 году клеточные биологи выделили из собак поджелудочной железы 11S «белок распознавания сигналов» (по счастливой случайности также сокращенно «SRP»), который способствовал транслокации секреторных белков через мембрану эндоплазматического ретикулума . [12] Затем было обнаружено, что SRP содержит компонент РНК . [13] SRP РНК Сравнение генов разных видов показало, что спираль 8 SRP РНК высококонсервативна во всех сферах жизни . [14] Области вблизи 5'- и 3'-концов SRP РНК млекопитающих подобны доминантному семейству Alu средних повторяющихся последовательностей генома человека . [15] Теперь понятно, что ДНК Alu возникла из SRP РНК путем вырезания центрального SRP-РНК-специфичного (S) фрагмента с последующей обратной транскрипцией человека и интеграцией во множество участков хромосом . [3] SRP-РНК были идентифицированы также в некоторых органеллах , например, в пластидных SRP многих фотосинтезирующих организмов. [16] области ядерных рибосом и во внутренней транскрибируемой спейсерной некоторых эктомикоризных грибов. [17]
Транскрипция и обработка
[ редактировать ]Эукариотические SRP-РНК транскрибируются с ДНК с помощью РНК-полимеразы III (Pol III). [18] РНК-полимераза III также транскрибирует гены 5S- рибосомальной РНК, тРНК , 7SK-РНК и сплайсосомальной РНК U6 . Промоторы генов SRP РНК человека включают элементы , расположенные ниже места начала транскрипции. РНК SRP растений Промоторы содержат вышестоящий стимулирующий элемент (USE) и ТАТА-бокс . [ нужна ссылка ] Гены РНК SRP дрожжей имеют ТАТА-бокс и дополнительные внутригенные промоторные последовательности (называемые A- и B-блоками), которые играют роль в регуляции транскрипции гена SRP с помощью Pol III. [19] У бактерий полимеразой гены организованы в опероны и транскрибируются РНК- . [ нужна ссылка ] 5'-конец малой (4,5S) SRP РНК многих бактерий расщепляется P. РНКазой [20] Концы SRP-РНК Bacillus subtilis процессируются РНКазой III . SRP РНК до сих пор не Интронов наблюдалось. [ нужна ссылка ]
Функция
[ редактировать ]
Котрансляционная транслокация
[ редактировать ]РНК SRP является неотъемлемой частью малого и большого домена SRP. Функция малого домена заключается в задержке трансляции белка до тех пор, пока связанный с рибосомой SRP не получит возможность ассоциироваться с мембрано-резидентным рецептором SRP (SR). Внутри большого домена SRP-РНК SRP, заряженного сигнальным пептидом, способствует гидролизу двух молекул гуанозинтрифосфата (GTP). Эта реакция высвобождает SRP из рецептора SRP и рибосомы , позволяя трансляцию продолжить и белку войти в транслокон . [21] Белок пересекает мембрану котрансляционно (во время трансляции) и попадает в другой клеточный компартмент или внеклеточное пространство. У эукариот мишенью является мембрана эндоплазматической сети (ЭР). У архей SRP доставляет белки к плазматической мембране . [22] У бактерий SRP в первую очередь включает белки во внутреннюю мембрану. [23]
Посттрансляционный транспорт
[ редактировать ]SRP участвует также в сортировке белков после завершения их синтеза (посттрансляционная сортировка белков). У эукариот белки, закрепленные на хвосте и имеющие гидрофобную инсерционную последовательность на С-конце, доставляются в эндоплазматический ретикулум (ЭР) с помощью SRP. [24] Аналогичным образом, SRP способствует посттрансляционному импорту белков, кодируемых в ядре, в тилакоидную мембрану хлоропластов . [25]
Структура
[ редактировать ]
В 2005 году в номенклатуре всех SRP-РНК была предложена система нумерации из 12 спиралей. Секции спирали обозначаются суффиксом строчной буквы ( например, 5a). Вставкам или «ветвям» спирали присвоены номера, обозначенные точками (например, 9.1 и 12.1).
РНК SRP охватывает широкий филогенетический спектр по размеру и количеству структурных особенностей (см. Примеры вторичной структуры РНК SRP ниже). Наименьшие функциональные SRP-РНК обнаружены у микоплазмы и родственных видов. РНК SRP Escherichia coli (также называемая 4,5S РНК) состоит из 114 нуклеотидных РНК остатков и образует стебель-петлю . бактерия Грамположительная РНК Bacillus subtilis кодирует более крупную 6S SRP РНК, которая напоминает архей, гомологи но лишена 6-й спирали РНК SRP. SRP архей имеют спирали от 1 до 8, лишены спирали 7 и характеризуются третичной структурой , включающей апикальные петли. спирали 3 и спирали 4. Эукариотические SRP РНК лишены спирали 1 и содержат спираль 7 вариабельного размера. Некоторые SRP-РНК простейших имеют редуцированные спирали 3 и 4. SRP-РНК ascomycota имеют полностью уменьшенный малый домен и лишены спиралей 3 и 4. Самые крупные SRP-РНК, известные на сегодняшний день, обнаружены у дрожжей ( Saccharomycetes ), которые приобрели спирали с 9 по 12 в виде вставки в спираль 5, а также расширенную спираль 7. растения экспрессируют многочисленные сильно дивергентные SRP РНК. [4]
Узоры
[ редактировать ]Были идентифицированы четыре консервативных признака (мотива) (показаны на рисунке темно-серым цветом): (1) мотив связывания SRP54, (2) мотив тетрапетли Helix 6 GNAR, (3) мотив 5e и (4) UGU(NR). мотив. [ нужна ссылка ]
SRP54 привязка
[ редактировать ]Асимметричная петля между спиральными участками 8a и 8b и прилегающим участком 8b с парными основаниями является важным свойством каждой SRP РНК. Спиральный участок 8b содержит пары оснований, отличные от Уотсона-Крика , которые способствуют образованию уплощенной малой бороздки в РНК, пригодной для связывания белка SRP54 (называемого у бактерий Ffh). [7] Апикальная от петля спирали 8 содержит четыре, пять или шесть остатков в зависимости вида . Он имеет высококонсервативный гуанозин в качестве первого остатка петли и аденозин в качестве последнего остатка петли. Эта особенность необходима для взаимодействия с третьим аденозиновым остатком тетрапетлевого мотива спирали 6 GNAR. [26]
Тетрапетля Helix 6 GNAR
[ редактировать ]РНК SRP эукариот и архей имеют тетрапетлю GNAR (N — любой нуклеотид , R — пурин ) в спирали 6. Ее консервативный остаток аденозина важен для связывания белка SRP19. [27] Этот аденозин осуществляет третичное взаимодействие с другим остатком аденозина, расположенным в апикальной петле спирали 8. [28]
5е
[ редактировать ]11 нуклеотидов мотива 5e образуют четыре пары оснований , которые прерываются петлей из трех нуклеотидов . [5] У эукариот первый нуклеотид петли представляет собой аденозин , необходимый для связывания белка SRP72. [29]
ПЕРВЫЙ(НР)
[ редактировать ]Мотив UGU(NR) соединяет спирали 3 и 4 в малом (Alu) домене SRP. Грибные SRP-РНК, лишенные спиралей 3 и 4, содержат мотив внутри петли спирали 2. [5] Это важно для связывания белка гетеродимера SRP9/14 как части РНК разворота . [30]
вторичный
[ редактировать ]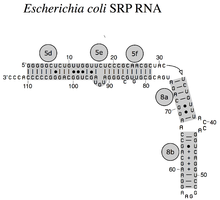 Бактериальная SRP РНК (4,5S РНК) из E. coli
Бактериальная SRP РНК (4,5S РНК) из E. coli Бактериальная SRP РНК (6S РНК) из Bacillus subtilis
Бактериальная SRP РНК (6S РНК) из Bacillus subtilis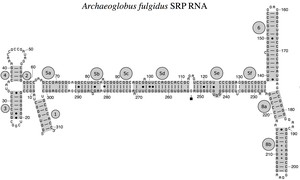 Архейная SRP РНК Archaeoglobus fulgidus
Архейная SRP РНК Archaeoglobus fulgidus эукариотических протистов РНК SRP Trypanosoma brucei
эукариотических протистов РНК SRP Trypanosoma brucei РНК SRP эукариотических дрожжей из Saccharomyces cerevisiae
РНК SRP эукариотических дрожжей из Saccharomyces cerevisiae





Третичный
[ редактировать ]| SRP РНК | |
|---|---|
| Идентификаторы | |
| Рфам | CL00003 |
| Другие данные | |
| PDB Структуры | PDBe 2IY3 , 1Z43 , 1RY1 , 1QZW , 1MFQ , 1L9A , 1LNG , 1JID , 1E8S , 1E8O , 1DUL , 1DUH , 1D4R , 28SR , 28SP |
Рентгеновская кристаллография , ядерный магнитный резонанс (ЯМР) и криоэлектронная микроскопия (крио-ЭМ] использовались для определения молекулярной структуры частей РНК SRP различных видов . Доступные структуры PDB показывают, что молекула РНК либо свободна, либо свободна. или когда он связан с одним или несколькими белками SRP .
![SRP19-7S.S Комплекс РНК SRP из M. jannaschii[28]](//upload.wikimedia.org/wikipedia/commons/thumb/1/16/PDB_1lng_EBI.jpg/267px-PDB_1lng_EBI.jpg) SRP19-7S.S Комплекс РНК SRP из M. jannaschii [28]
SRP19-7S.S Комплекс РНК SRP из M. jannaschii [28]![S-домен SRP человека[31]](//upload.wikimedia.org/wikipedia/commons/thumb/0/08/PDB_1mfq_EBI.jpg/267px-PDB_1mfq_EBI.jpg) S-домен SRP человека [31]
S-домен SRP человека [31]
![SRP19-7S.S Комплекс РНК SRP из M. jannaschii[28]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/16/PDB_1lng_EBI.jpg/267px-PDB_1lng_EBI.jpg)
![S-домен SRP человека[31]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/08/PDB_1mfq_EBI.jpg/267px-PDB_1mfq_EBI.jpg)
Связывающие белки
[ редактировать ]SRP Один или несколько белков SRP, связываются с РНК образуя функциональный SRP. Белки SRP названы в соответствии с их приблизительной молекулярной массой, измеряемой в килодальтонах . [32] Большинство бактериальных SRP состоят из SRP РНК и SRP54 (также называемого Ffh, что означает « гомолог четвертый » ) пятьдесят . SRP архей содержит белки SRP54 и SRP19. У эукариот РНК SRP сочетается с импортированными белками SRP SRP9/14, SRP19 и SRP68/72 в области ядрышка . Этот пре-SRP транспортируется в цитозоль , где связывается с белком SRP54. [33] Молекулярные структуры свободных или SRP-РНК-связанных белков SRP9/14, SRP19 или SRP54 известны с высоким разрешением.
SRP9 и SRP14
[ редактировать ]SRP9 и SRP14 структурно SRP9/14, родственны и образуют гетеродимер который связывается с РНК SRP малого (Alu) домена. [30] В дрожжевом SRP отсутствует SRP9, но он содержит структурно родственный связывающий белок SRP21. Дрожжевой SRP14 образует гомодимеры в кристалле и не связывает Alu. [34] SRP9/14 отсутствует в SRP трипаносомы , которая вместо этого содержит тРНК -подобную молекулу. [35]
СРП19
[ редактировать ]SRP19 обнаружен в SRP эукариот и архей . Его основная роль заключается в подготовке РНК SRP для связывания SRP54, SRP68 и SRP72 путем правильного расположения спиралей 6 и 8 SRP РНК. [31] Дрожжевой SRP содержит Sec65p, более крупный гомолог SRP19. [36]
СРП54
[ редактировать ]Белок SRP54 (называемый у бактерий Ffh ) является важным компонентом каждого SRP. Он состоит из трех функциональных доменов : N-концевого (N) домена, домена ГТФазы (G) и богатого метионином (M) домена. [37] [38]
СРП68 и СРП72
[ редактировать ]Белки SRP68 и SRP72 являются структурно несвязанными компонентами большого домена эукариотического SRP. Они образуют стабильный гетеродимер SRP68/72. Было показано, что около одной трети белка SRP68 человека связывается с РНК SRP. [39] Относительно небольшая область, расположенная вблизи С-конца SRP72, связывается с мотивом 5e SRP РНК. [29] [40]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000276168 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Уллу Э, Чуди С (1984). «Последовательности Alu обрабатываются генами 7SL РНК». Природа . 312 (5990): 171–172. Бибкод : 1984Natur.312..171U . дои : 10.1038/312171a0 . ПМИД 6209580 . S2CID 4328237 .
- ^ Jump up to: а б Розенблад М.А., Ларсен Н., Самуэльссон Т., Цвиб С. (2009). «Родство в семействе SRP РНК» . Биология РНК . 6 (5): 508–516. дои : 10.4161/rna.6.5.9753 . ПМИД 19838050 .
- ^ Jump up to: а б с д Регалия М., Розенблад М.А., Самуэльссон Т. (август 2002 г.). «Прогнозирование сигналов распознавания генов РНК частиц» . Исследования нуклеиновых кислот . 30 (15): 3368–3377. дои : 10.1093/nar/gkf468 . ПМК 137091 . ПМИД 12140321 .
- ^ Jump up to: а б с Уайлд К., Вайхенридер О., Струб К., Синнинг И., Кьюсак С. (февраль 2002 г.). «К структуре частицы распознавания сигналов млекопитающих». Современное мнение в области структурной биологии . 12 (1): 72–81. дои : 10.1016/S0959-440X(02)00292-0 . ПМИД 11839493 .
- ^ Jump up to: а б с Бэти РТ, Рэмбо Р.П., Лукаст Л., Ра Б., Дудна Дж.А. (февраль 2000 г.). «Кристаллическая структура рибонуклеопротеинового ядра частицы распознавания сигнала». Наука . 287 (5456): 1232–1239. Бибкод : 2000Sci...287.1232B . дои : 10.1126/science.287.5456.1232 . ПМИД 10678824 .
- ^ Бэти Р.Т., Сагар М.Б., Дудна Дж.А. (март 2001 г.). «Структурный и энергетический анализ распознавания РНК универсально консервативным белком из частицы распознавания сигнала». Журнал молекулярной биологии . 307 (1): 229–246. дои : 10.1006/jmbi.2000.4454 . ПМИД 11243816 .
- ^ Епископ Дж.М., Левинсон В.Е., Салливан Д., Фаншир Л., Квинтрелл Н., Джексон Дж. (декабрь 1970 г.). «Низкомолекулярные РНК вируса саркомы Рауса. II. 7S РНК». Вирусология . 42 (4): 927–937. дои : 10.1016/0042-6822(70)90341-7 . ПМИД 4321311 .
- ^ Уокер Т.А., Пейс Н.Р., Эриксон Р.Л., Эриксон Э., Бер Ф. (сентябрь 1974 г.). «7S РНК, общая для онкорнавирусов и нормальных клеток, связана с полирибосомами» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (9): 3390–3394. Бибкод : 1974PNAS...71.3390W . дои : 10.1073/pnas.71.9.3390 . ПМЦ 433778 . ПМИД 4530311 .
- ^ Зиве Г., Пенман С. (май 1976 г.). «Малые виды РНК клетки HeLa: метаболизм и субклеточная локализация». Клетка . 8 (1): 19–31. дои : 10.1016/0092-8674(76)90181-1 . ПМИД 954090 . S2CID 26928799 .
- ^ Уолтер П., Ибрагими И., Блобель Дж. (ноябрь 1981 г.). «Транслокация белков через эндоплазматический ретикулум. I. Белок распознавания сигнала (SRP) связывается с полисомами, собранными in vitro, синтезирующими секреторный белок» . Журнал клеточной биологии . 91 (2 ч. 1): 545–550. дои : 10.1083/jcb.91.2.545 . ПМК 2111968 . ПМИД 7309795 .
- ^ Уолтер П., Блобель Дж. (октябрь 1982 г.). «Частица распознавания сигнала содержит 7S РНК, необходимую для транслокации белка через эндоплазматический ретикулум». Природа . 299 (5885): 691–698. Бибкод : 1982Natur.299..691W . дои : 10.1038/299691a0 . ПМИД 6181418 . S2CID 4237513 .
- ^ Ларсен Н., Звиб С. (январь 1991 г.). «Выравнивание последовательности SRP-РНК и вторичная структура» . Исследования нуклеиновых кислот . 19 (2): 209–215. дои : 10.1093/нар/19.2.209 . ПМЦ 333582 . ПМИД 1707519 .
- ^ Уллу Э., Мерфи С., Мелли М. (май 1982 г.). «Человеческая 7SL РНК состоит из 140-нуклеотидной среднеповторяющейся последовательности, вставленной в последовательность alu». Клетка . 29 (1): 195–202. дои : 10.1016/0092-8674(82)90103-9 . ПМИД 6179628 . S2CID 12709599 .
- ^ Розенблад М.А., Самуэльссон Т. (ноябрь 2004 г.). «Идентификация генов РНК частиц распознавания сигналов хлоропластов» . Физиология растений и клеток . 45 (11): 1633–1639. дои : 10.1093/pcp/pch185 . ПМИД 15574839 .
- ^ Альм Розенблад М., Мартин М.П., Тедерсо Т., Райберг М.Р., Ларссон Э., Вурцбахер С., Абаренков К., Нильссон Р.Х. (2016). «Обнаружение РНК сигнальных частиц (SRP) в ядерном рибосомальном внутреннем транскрибируемом спейсере 1 (ITS1) трех линий эктомикоризных грибов» . Микокейс . 13 : 21–33. дои : 10.3897/mycokeys.13.8579 . hdl : 10261/163935 .
- ^ Диечи Г., Фиорино Г., Кастельнуово М., Тейхманн М., Пагано А. (декабрь 2007 г.). «Расширяющийся транскриптом РНК-полимеразы III». Тенденции в генетике . 23 (12): 614–622. дои : 10.1016/j.tig.2007.09.001 . hdl : 11381/1706964 . ПМИД 17977614 .
- ^ Диечи Дж., Джулиодори С., Кателлани М., Перкудани Р., Оттонелло С. (март 2002 г.). «Адаптация внутригенного промотора и облегчение рециркуляции РНК-полимеразы III при транскрипции SCR1, гена РНК 7SL Saccharomyces cerevisiae» . Журнал биологической химии . 277 (9): 6903–6914. дои : 10.1074/jbc.M105036200 . ПМИД 11741971 .
- ^ Ботвелл, Алабама; Гарбер, РЛ; Альтман, С. (10 декабря 1976 г.). «Нуклеотидная последовательность и процессинг in vitro молекулы-предшественника 4,5 S РНК Escherichia coli» . Журнал биологической химии . 251 (23): 7709–16. дои : 10.1016/S0021-9258(17)32909-5 . ПМИД 794064 .
- ^ Шан С.О., Уолтер П. (февраль 2005 г.). «Котрансляционное нацеливание на белок с помощью частицы распознавания сигнала» . Письма ФЭБС . 579 (4): 921–926. дои : 10.1016/j.febslet.2004.11.049 . ПМИД 15680975 . S2CID 46046514 .
- ^ Звиеб С., Эйхлер Дж. (март 2002 г.). «Добираемся до цели: архейская частица распознавания сигналов» . Архея . 1 (1): 27–34. дои : 10.1155/2002/729649 . ПМЦ 2685543 . ПМИД 15803656 .
- ^ Ульбрандт Н.Д., Ньюитт Дж.А., Бернштейн Х.Д. (январь 1997 г.). «Частица распознавания сигнала E. coli необходима для внедрения подмножества белков внутренней мембраны» . Клетка . 88 (2): 187–196. дои : 10.1016/S0092-8674(00)81839-5 . ПМИД 9008159 . S2CID 15246619 .
- ^ Абелл Б.М., Пул М.Р., Шленкер О., Синнинг И., Хай С. (июль 2004 г.). «Частица распознавания сигнала опосредует посттрансляционное нацеливание у эукариот» . Журнал ЭМБО . 23 (14): 2755–2764. дои : 10.1038/sj.emboj.7600281 . ПМК 514945 . ПМИД 15229647 .
- ^ Шунеманн Д., Гупта С., Перселло-Картье Ф., Климюк В.И., Джонс Дж.Д., Нуссауме Л., Хоффман Н.Е. (август 1998 г.). «Новая частица, распознающая сигнал, направляет светособирающие белки на мембраны тилакоидов» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 10312–10316. Бибкод : 1998PNAS...9510312S . дои : 10.1073/pnas.95.17.10312 . ПМК 21505 . ПМИД 9707644 .
- ^ Звиеб С., ван Нуэс Р.В., Розенблад М.А., Браун Дж.Д., Самуэльссон Т. (январь 2005 г.). «Номенклатура для всех РНК частиц распознавания сигналов» . РНК . 11 (1): 7–13. дои : 10.1261/rna.7203605 . ПМК 1370686 . ПМИД 15611297 .
- ^ Звиб С. (август 1992 г.). «Распознавание тетрануклеотидной петли РНК частицы распознавания сигнала белком SRP19» . Журнал биологической химии . 267 (22): 15650–15656. дои : 10.1016/S0021-9258(19)49585-9 . ПМИД 1379233 .
- ^ Jump up to: а б Хайнцл Т., Хуанг С., Зауэр-Эрикссон А.Е. (июнь 2002 г.). «Структура комплекса РНК SRP19 и значение для сборки частиц распознавания сигналов». Природа . 417 (6890): 767–771. Бибкод : 2002Natur.417..767H . дои : 10.1038/nature00768 . ПМИД 12050674 . S2CID 2509475 .
- ^ Jump up to: а б Яхьяева Э., Вауэр Дж., Вауэр И.К., Звиб С. (июнь 2008 г.). «Мотив 5e РНК эукариотической частицы распознавания сигнала содержит консервативный аденозин для связывания SRP72» . РНК . 14 (6): 1143–1153. дои : 10.1261/rna.979508 . ПМК 2390789 . ПМИД 18441046 .
- ^ Jump up to: а б Вайхенридер О., Уайлд К., Струб К., Кьюсак С. (ноябрь 2000 г.). «Структура и сборка домена Alu частицы распознавания сигнала млекопитающих» . Природа . 408 (6809): 167–173. Бибкод : 2000Natur.408..167W . дои : 10.1038/35041507 . ПМИД 11089964 . S2CID 4427070 .
- ^ Jump up to: а б Куглстаттер А., Обридж С., Нагай К. (2002). «Индуцированные структурные изменения 7SL РНК во время сборки частицы распознавания сигналов человека». Нат Структ Биол . 9 (10): 740–744. дои : 10.1038/nsb843 . ПМИД 12244299 . S2CID 9543041 .
- ^ Уолтер П., Блобель Дж. (сентябрь 1983 г.). «Разборка и восстановление частицы распознавания сигнала». Клетка . 34 (2): 525–533. дои : 10.1016/0092-8674(83)90385-9 . ПМИД 6413076 . S2CID 17907778 .
- ^ Политц Дж.К., Яровой С., Килрой С.М., Гауда К., Звиб С., Педерсон Т. (январь 2000 г.). «Распознавание сигналов компонентов частиц в ядрышке» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (1): 55–60. Бибкод : 2000ПНАС...97...55П . дои : 10.1073/pnas.97.1.55 . ПМК 26615 . ПМИД 10618370 .
- ^ Брукс М.А., Равелли Р.Б., Маккарти А.А., Струб К., Кьюсак С. (май 2009 г.). «Структура SRP14 из частицы распознавания сигнала Schizosaccharomyces pombe» . Acta Crystallographica Раздел D. 65 (Часть 5): 421–433. дои : 10.1107/S0907444909005484 . ПМИД 19390147 .
- ^ Лю Л., Бен-Шломо Х., Сюй YX, Стерн М.З., Гончаров И., Чжан Ю., Михаэли С. (май 2003 г.). «Частица, распознающая сигнал трипаносоматида, состоит из двух молекул РНК, гомолога РНК 7SL и новой тРНК-подобной молекулы» . Журнал биологической химии . 278 (20): 18271–18280. дои : 10.1074/jbc.M209215200 . ПМИД 12606550 .
- ^ Ханн BC, Стирлинг CJ, Уолтер П. (апрель 1992 г.). «Продукт гена SEC65 представляет собой субъединицу частицы распознавания дрожжевого сигнала, необходимую для ее целостности». Природа . 356 (6369): 532–533. Бибкод : 1992Natur.356..532H . дои : 10.1038/356532a0 . ПМИД 1313947 . S2CID 4287636 .
- ^ Ремиш К., Уэбб Дж., Герц Дж., Прен С., Франк Р., Вингрон М., Добберштейн Б. (август 1989 г.). «Гомология белка 54K частицы распознавания сигнала, стыковочного белка и двух белков E. coli с предполагаемыми GTP-связывающими доменами» (PDF) . Природа . 340 (6233): 478–482. Бибкод : 1989Natur.340..478R . дои : 10.1038/340478a0 . ПМИД 2502717 . S2CID 4343347 .
- ^ Бернштейн Х.Д., Поритц М.А., Струб К., Хобен П.Дж., Бреннер С., Уолтер П. (август 1989 г.). «Модель распознавания сигнальной последовательности из аминокислотной последовательности субъединицы 54K частицы распознавания сигнала». Природа . 340 (6233): 482–486. Бибкод : 1989Natur.340..482B . дои : 10.1038/340482a0 . ПМИД 2502718 . S2CID 619959 .
- ^ Яхьяева Э, Бхуян Ш., Инь Дж, Звиб С (июнь 2006 г.). «Белок SRP68 частицы распознавания сигнала человека: идентификация связывающих доменов РНК и SRP72» . Белковая наука . 15 (6): 1290–1302. дои : 10.1110/ps.051861406 . ПМЦ 2242529 . ПМИД 16672232 .
- ^ Яхьяева Е, Инь Дж, Звиб С (январь 2005 г.). «Идентификация РНК-связывающего домена в SRP72 человека». Журнал молекулярной биологии . 345 (4): 659–666. дои : 10.1016/j.jmb.2004.10.087 . ПМИД 15588816 .
Дальнейшее чтение
[ редактировать ]- Куглстаттер А., Обридж С., Нагай К. (октябрь 2002 г.). «Индуцированные структурные изменения 7SL РНК во время сборки частицы распознавания сигналов человека». Структурная биология природы . 9 (10): 740–744. дои : 10.1038/nsb843 . ПМИД 12244299 . S2CID 9543041 .
- Ван Т, Тянь С, Чжан В, Луо К, Саркис П.Т., Ю Л, Лю Б, Ю Ю, Ю XF (декабрь 2007 г.). «7SL РНК опосредует упаковку вириона противовирусной цитидиндезаминазы APOBEC3G» . Журнал вирусологии . 81 (23): 13112–13124. дои : 10.1128/JVI.00892-07 . ПМК 2169093 . ПМИД 17881443 .
- Уллу Э., Вайнер А.М. (декабрь 1984 г.). «Человеческие гены и псевдогены компонента РНК 7SL частицы, распознающей сигнал» . Журнал ЭМБО . 3 (13): 3303–3310. дои : 10.1002/j.1460-2075.1984.tb02294.x . ПМЦ 557853 . ПМИД 6084597 .
- Энглерт М., Фелис М., Юнкер В., Бейер Х. (декабрь 2004 г.). «Новые восходящие и внутригенные элементы контроля РНК-полимеразы III-зависимой транскрипции генов 7SL РНК человека». Биохимия . 86 (12): 867–874. дои : 10.1016/j.biochi.2004.10.012 . ПМИД 15667936 .
- Оубридж С., Куглстаттер А., Джовин Л., Нагай К. (июнь 2002 г.). «Кристаллическая структура SRP19 в комплексе с S-доменом РНК SRP и ее значение для сборки частицы распознавания сигнала» . Молекулярная клетка . 9 (6): 1251–1261. дои : 10.1016/S1097-2765(02)00530-0 . ПМИД 12086622 .
- Ван Т, Тянь С, Чжан В, Саркис П.Т., Ю С.Ф. (январь 2008 г.). «Для упаковки вириона APOBEC3F необходимо взаимодействие с РНК 7SL, но не с геномной РНК ВИЧ-1 или P-тельцами». Журнал молекулярной биологии . 375 (4): 1098–1112. дои : 10.1016/j.jmb.2007.11.017 . ПМИД 18067920 .
Внешние ссылки
[ редактировать ]- База данных SRP (SRPDB): выравнивание РНК SRP и связанных с ними белков, вторичные структуры РНК SRP и трехмерные модели.
- Запись Rfam для РНК частицы распознавания сигнала типа Metazoan
- Запись Rfam для РНК бактериальных частиц, распознающих малые сигналы
- Запись Rfam для РНК бактериальной частицы распознавания большого сигнала
- Запись Rfam для РНК частицы распознавания грибкового сигнала
- Запись Rfam для РНК частицы распознавания сигналов растений
- Запись Rfam для РНК частицы распознавания сигнала простейших
- Запись Rfam для РНК частиц распознавания сигналов архей
- Фильм о частицах распознавания сигналов Dnatube