Мезенхима
| Мезенхима | |
|---|---|
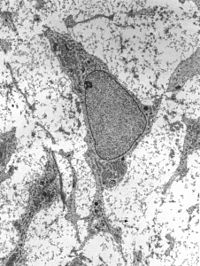 Просвечивающая электронная микрофотография мезенхимы, показывающая ультраструктуру типичной клетки и матрикса . | |
 Мезенхима (указатель), окрашенная H&E | |
| Подробности | |
| Этап Карнеги | 6б |
| Предшественник | Латеральная мезодерма |
| Идентификаторы | |
| ТО | Е5.16.4.0.3.0.18 |
| Анатомическая терминология | |
Мезенхима ( / ˈ m ɛ s ə n k aɪ m ˈ m iː z ən - / [1] ) — тип рыхло организованной эмбриональной соединительной ткани животных , состоящей из недифференцированных клеток , дающих начало большинству тканей, таких как кожа , кровь или кости . [2] [3] Взаимодействия между мезенхимой и эпителием помогают формировать почти каждый орган развивающегося эмбриона. [4]
Позвоночные животные
[ редактировать ]Структура
[ редактировать ]Мезенхима морфологически характеризуется выраженным основного вещества, матриксом содержащим рыхлый агрегат ретикулярных волокон и неспециализированных мезенхимальных стволовых клеток . [5] Мезенхимальные клетки могут легко мигрировать (в отличие от эпителиальных клеток , которые лишены подвижности, организованы в плотно прилегающие листы и поляризованы в апикально-базальной ориентации).
Разработка
[ редактировать ]Мезенхима происходит из мезодермы . [6] Из мезодермы мезенхима выглядит как эмбриологически примитивный «суп». Этот «суп» существует как комбинация мезенхимальных клеток, серозной жидкости и множества различных тканевых белков. Серозная жидкость обычно содержит множество серозных элементов, таких как натрий и хлорид. Мезенхима развивается в ткани лимфатической и кровеносной систем, а также опорно-двигательного аппарата. Эта последняя система характеризуется наличием соединительных тканей по всему телу, таких как кость и хрящ . Злокачественный рак мезенхимальных клеток — разновидность саркомы . [7] [8]
Эпителиально-мезенхимальный переход
[ редактировать ]Первое появление мезенхимы происходит во время гаструляции в результате процесса эпителиально-мезенхимального перехода (ЕМТ). Этот переход происходит за счет потери эпителиального кадгерина , плотных и слипчивых соединений на клеточных мембранах эпителиальных клеток . [9] Поверхностные молекулы подвергаются эндоцитозу , а цитоскелет микротрубочек теряет форму, позволяя мезенхиме мигрировать вдоль внеклеточного матрикса (ECM). Эпителиально-мезенхимальный переход происходит в эмбриональных клетках, которым требуется миграция через ткань или поверх нее, и за ним может следовать мезенхимально-эпителиальный переход с образованием вторичных эпителиальных тканей .Эмбриологические мезенхимальные клетки экспрессируют белок S100-A4 ( S100A4 ). [10] также известный как белок, специфичный для фибробластов , [11] что указывает на их общие свойства с мигрирующими взрослыми фибробластами и c-Fos , онкогеном, связанным с подавлением эпителиального кадгерина. [12] [13] Как формирование примитивной полоски , так и мезенхимальной ткани зависит от пути Wnt/β-catenin . [14] Специфические маркеры мезенхимальной ткани включают дополнительную экспрессию факторов ЕСМ, таких как фибронектин и витронектин . [15]
Имплантация
[ редактировать ]Первыми клетками эмбриона, подвергающимися ЭМП и образующими мезенхиму, являются внеэмбриональные клетки трофэктодермы . Они мигрируют из тела бластоцисты в слой эндометрия матки , чтобы способствовать образованию прикрепленной плаценты . [16]
Первичная мезенхима
[ редактировать ]Первичная мезенхима — это первая возникающая эмбриональная мезенхимальная ткань, которая образуется из ЕМТ в эпибласта клетках . В эпибласте он индуцируется примитивной полоской посредством передачи сигналов Wnt и образует энтодерму и мезодерму из переходной ткани, называемой мезендодермой, во время процесса гаструляции . [17]
Формирование первичной мезенхимы зависит от экспрессии WNT3 . Другие недостатки в сигнальных путях, например, в Nodal (белок TGF-бета), приведут к дефектному формированию мезодермы . [9]
Слои ткани, образовавшиеся из примитивной полоски, вместе инвагинируются в эмбрион, и индуцированные мезенхимальные стволовые клетки проникают в него и образуют мезодерму . Мезодермальная ткань будет продолжать дифференцироваться и/или мигрировать по всему эмбриону, в конечном итоге образуя большинство слоев соединительной ткани тела. [18]
Нейральная мезенхима
[ редактировать ]Эмбриологическая мезенхима особенно преходяща и вскоре дифференцируется после миграции. Нейральная мезенхима формируется вскоре после образования первичной мезенхимы. [19]
Взаимодействие с эктодермой и морфогенными факторами, образующими сомиты, приводит к тому, что некоторые первичные мезенхимы образуют нервную мезенхиму или параксиальную мезодерму и способствуют образованию сомитов . Нейральная мезенхима вскоре претерпевает мезенхимально-эпителиальный переход под влиянием WNT6, продуцируемого эктодермой , с образованием сомитов . [20] Эти структуры подвергаются вторичной ЭМП, поскольку ткань сомитов на более позднем этапе развития мигрирует с образованием структурной соединительной ткани, такой как хрящ и скелетные мышцы . [21]
Клетки нервного гребня (NCC) образуются из нейроэктодермы , а не из первичной мезенхимы, из морфогенных сигналов нервного гребня . ЕМТ происходит в результате передачи сигналов Wnt , влияния генов Sox и потери E-кадгерина с поверхности клетки. NCC дополнительно требуют репрессии N-кадгерина и молекулы адгезии нервных клеток . НКС проникают в эмбрион из эпителиального нейроэктодермального слоя и мигрируют по всему организму, образуя множественные периферической нервной системы клетки (ПНС) и меланоциты . Миграция NCCs в первую очередь индуцируется передачей сигналов BMP и его ингибитором Noggin . [22] [23]
Беспозвоночные
[ редактировать ]У некоторых беспозвоночных , таких как Porifera , Cnidaria , Ctenophora и некоторых триплобластов (а именно ацеломатов ), термин «мезенхима» относится к более или менее твердой, но рыхло организованной ткани, состоящей из гелевого матрикса ( мезоглеи ) с различные клеточные и волокнистые включения, расположенные между эпидермисом и гастродермой (у нетриплобластных животных обычно считается отсутствие «соединительной» ткани). В некоторых случаях мезоглея бесклеточная. [24]
- У губок мезенхима называется мезогилом . [25]
- В диплобластах (Cnidaria и Ctenophora) мезенхима полностью происходит из эктодермы. Этот вид мезенхимы называется эктомезодермальным и не считается настоящей мезодермой .
- У триблобластных ацеломатов (таких как плоские черви ) термин паренхима иногда используется для обозначения среднего (мезенхимального) слоя, в котором плотный слой включает ткани, происходящие как из эктодермы, так и из энтомезодермы (истинная мезодерма, происходящая из энтодермы ).
Когда клеточный материал редкий или плотно упакован, как у книдарий, мезенхиму иногда называют колленхимой или паренхимой у плоских червей. [25] Когда клеточный материал отсутствует, как у Hydrozoa ), этот слой правильно называется мезоглеей . [25]
У некоторых колониальных книдарий мезенхима перфорирована гастроваскулярными каналами, проходящими между членами колонии. Вся эта матрица общего базального материала называется коенхимой . [25]
Ссылки
[ редактировать ]- ^ «Мезенхима, английское определение и значение | Lexico.com» . Архивировано из оригинала 29 сентября 2019 года.
- ^ Сэдлер, Т.В. (2010). Медицинская эмбриология Лангмана (11-е изд.). Филадельфия: Липпинкотт Уильям и Уилкинс. п. 70. ИСБН 9780781790697 .
- ^ «Определение МЕЗЕНХИМЫ» . Мерриам-Вебстер . Архивировано из оригинала 4 февраля 2024 года.
- ^ МакКорд, Кейт (14 сентября 2012 г.). «Мезенхима» . Энциклопедия проекта «Эмбрион» . Университет штата Аризона . Архивировано из оригинала 20 января 2024 года.
- ^ Сломянка, Лутц. «Голубая гистология – соединительные ткани» . Школа анатомии и биологии человека — Университет Западной Австралии . Архивировано из оригинала 7 марта 2020 г.
- ^ Кирзенбаум, Авраам Л.; Трес, Лаура (2015). Электронная книга «Гистология и клеточная биология: введение в патологию» (4-е изд.). Elsevier Науки о здоровье. п. 123. ИСБН 9780323313353 .
- ^ Страм, Джуди М.; Гартнер, Лесли П.; Хайатт, Джеймс Л. (2007). Клеточная биология и гистология . Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. п. 83 . ISBN 978-0-7817-8577-8 .
- ^ Сэдлер, Т.В. (2006). Медицинская эмбриология Лангмана . Липпинкотт Уильямс и Уилкинс . стр. 68–70. ISBN 978-0-7817-9485-5 .
- ^ Jump up to: а б Каллури, Рагху; Вайнберг, Роберт А. (2009). «Основы эпителиально-мезенхимального перехода» . Журнал клинических исследований . 119 (6): 1420–8. дои : 10.1172/JCI39104 . ПМК 2689101 . ПМИД 19487818 .
- ^ «S100A4 - Белок S100-A4 - Homo sapiens (Человек)» . ЮниПрот . Архивировано из оригинала 21 ноября 2021 г.
- ^ Остеррайхер, Кристоф Х.; Пенц-Остеррайхер, Мелитта; Гривенников, Сергей И. (04 января 2011 г.). «Специфический для фибробластов белок 1 идентифицирует воспалительную субпопуляцию макрофагов в печени» . Труды Национальной академии наук . 108 (1): 308–313. Бибкод : 2011PNAS..108..308O . дои : 10.1073/pnas.1017547108 . ПМК 3017162 . ПМИД 21173249 .
- ^ Окада, Х; Данофф, ТМ; Каллури, Р; Нилсон, Э.Г. (1997). «Ранняя роль Fsp1 в эпителиально-мезенхимальной трансформации» . Американский журнал физиологии . 273 (4 ч. 2): F563–74. дои : 10.1152/ajprenal.1997.273.4.F563 . ПМИД 9362334 .
- ^ Эгер, А; Стокингер, А; Шаффхаузер, Б; Бег, Х; Фойснер, Р. (2000). «Эпителиально-мезенхимальный переход путем активации эстрогенового рецептора c-Fos включает ядерную транслокацию бета-катенина и активацию транскрипционной активности бета-катенина/лимфоидного энхансера, связывающего фактор-1» . Журнал клеточной биологии . 148 (1): 173–88. дои : 10.1083/jcb.148.1.173 . ПМК 3207144 . ПМИД 10629227 .
- ^ Мохамед, ОА; Кларк, HJ; Дюфорт, Д. (2004). «Передача сигналов бета-катенина отмечает предполагаемое место формирования примитивных полосок в эмбрионе мыши» . Динамика развития . 231 (2): 416–24. дои : 10.1002/dvdy.20135 . ПМИД 15366019 . S2CID 39908122 .
- ^ Тьери, JP; Слиман, JP (2006). «Сложные сети управляют эпителиально-мезенхимальными переходами» (PDF) . Nature Reviews Молекулярно-клеточная биология . 7 (2): 131–42. дои : 10.1038/nrm1835 . ПМИД 16493418 . S2CID 8435009 . Архивировано из оригинала (PDF) 26 июня 2013 г.
- ^ Беллерс, Р. (1986). «Примитивная полоса». Анатомия и эмбриология . 174 (1): 1–14. дои : 10.1007/bf00318331 . ПМИД 3518538 . S2CID 33629601 .
- ^ Хэй, ЭД (2005). «Мезенхимальная клетка, ее роль в эмбрионе и замечательные сигнальные механизмы, которые ее создают» . Динамика развития . 233 (3): 706–20. дои : 10.1002/dvdy.20345 . ПМИД 15937929 . S2CID 22368548 .
- ^ Марески, К; Новара, М; Рустичелли, Д; Ферреро, я; Гвидо, Д; Карбоне, Э; Медико, Э; Мэдон, Э; Верчелли, А; Фаджиоли, Ф (2006). «Нейральная дифференциация мезенхимальных стволовых клеток человека: данные об экспрессии нейронных маркеров и типов каналов K+ eag» . Экспериментальная гематология . 34 (11): 1563–72. дои : 10.1016/j.exphem.2006.06.020 . ПМИД 17046576 .
- ^ Шмидт, К; Штёкельхубер, М; Маккиннелл, я; Путц, Р; Христос, Б; Патель, К. (2004). «Wnt 6 регулирует процесс эпителизации сегментарной пластинки мезодермы, приводящий к образованию сомитов» . Биология развития . 271 (1): 198–209. дои : 10.1016/j.ydbio.2004.03.016 . ПМИД 15196961 .
- ^ Стокдейл, FE; Никовиц-младший, Вт; Христос, Б. (2000). «Молекулярная и клеточная биология развития сомитов птиц». Динамика развития . 219 (3): 304–21. doi : 10.1002/1097-0177(2000)9999:9999<::AID-DVDY1057>3.0.CO;2-5 . ПМИД 11066088 . S2CID 32342256 .
- ^ Броннер-Фрейзер, М. (1994). «Формирование и миграция клеток нервного гребня в развивающемся эмбрионе» . Журнал ФАСЭБ . 8 (10): 699–706. дои : 10.1096/fasebj.8.10.8050668 . ПМИД 8050668 . S2CID 12161494 .
- ^ Трейнор, Пенсильвания (2005). «Характеристика формирования и миграции клеток нервного гребня у эмбрионов мыши». Семинары по клеточной биологии и биологии развития . 16 (6): 683–93. дои : 10.1016/j.semcdb.2005.06.007 . ПМИД 16043371 .
- ^ Бруска, Колорадо; Бруска, Дж.Дж. (2003). Беспозвоночные (2-е изд.). Сандерленд, Массачусетс. п. 101 . ISBN 9780878930975 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Jump up to: а б с д Бруска, Колорадо; Бруска, Дж.Дж. (2003). Беспозвоночные (2-е изд.). Сандерленд, Массачусетс. п. 220 . ISBN 9780878930975 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка )
| Базы данных органов управления : Национальные |
|---|