Нервный гребень
| Нервный гребень | |
|---|---|
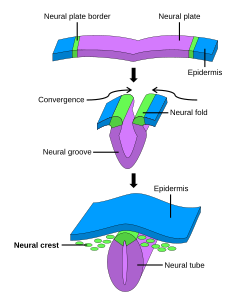 Формирование нервного гребня в процессе нейруляции. Нервный гребень сначала индуцируется в области границы нервной пластинки . После закрытия нервной трубки нервный гребень отделяется от области между дорсальной нервной трубкой и окружающей эктодермой и мигрирует к периферии. | |
| Идентификаторы | |
| МеШ | D009432 |
| ТО | crest_by_E5.0.2.1.0.0.2 E5.0.2.1.0.0.2 |
| ФМА | 86666 |
| Анатомическая терминология | |
Клетки нервного гребня представляют собой временную группу клеток, которые возникают из эмбриональной эктодермы зародышевого листка и, в свою очередь, дают начало разнообразному клеточному клону, включая меланоциты , черепно-лицевой хрящ и кости, гладкие мышцы , периферические и кишечные нейроны и глию . [1] [2]
После гаструляции на границе нервной пластинки и ненейральной эктодермы определяются клетки нервного гребня . Во время нейруляции границы нервной пластинки, также известные как нервные складки , сходятся на дорсальной срединной линии, образуя нервную трубку . [3] Впоследствии клетки нервного гребня из пластинки крыши нервной трубки претерпевают эпителиально-мезенхимальный переход , отслаиваясь от нейроэпителия и мигрируя через периферию, где они дифференцируются в различные типы клеток. [1] Появление нервного гребня сыграло важную роль в эволюции позвоночных позвоночных , поскольку многие из его структурных производных являются определяющими особенностями клады . [4]
В основе развития нервного гребня лежит регуляторная сеть генов , описываемая как набор взаимодействующих сигналов, факторов транскрипции и нижестоящих эффекторных генов, которые придают клеткам такие характеристики, как мультипотентность и миграционные способности. [5] Понимание молекулярных механизмов формирования нервного гребня важно для наших знаний о болезнях человека из-за его вклада в множественные клеточные линии . Аномалии развития нервного гребня вызывают нейрокристопатии , которые включают такие состояния, как лобно-носовая дисплазия , синдром Ваарденбурга-Шаха и синдром ДиДжорджа . [1]
Следовательно, определение механизмов развития нервного гребня может раскрыть ключевую информацию об эволюции позвоночных и нейрокристопатиях.
История
[ редактировать ]Нервный гребень был впервые описан у куриного эмбриона Вильгельмом Хисом-старшим в 1868 году как «промежуточный шнур» (Zwischenstrang) из-за его происхождения между нервной пластинкой и ненейральной эктодермой. [1] Он назвал тканевый ганглиозный гребень, поскольку его конечным пунктом назначения была каждая боковая сторона нервной трубки, где он дифференцировался в спинальные ганглии. [6] В первой половине 20 века большая часть исследований нервного гребня проводилась с использованием эмбрионов амфибий, обзор которых был сделан Хёрстадиусом (1950) в известной монографии. [7]
Методы маркировки клеток продвинули область исследования нервного гребня, поскольку позволили исследователям визуализировать миграцию ткани по развивающимся эмбрионам. В 1960-х годах Уэстон и Чибон использовали радиоизотопное мечение ядра тритиевым тимидином у эмбрионов кур и амфибий соответственно. Однако этот метод страдает недостатками стабильности, поскольку каждый раз, когда меченая клетка делится, сигнал ослабляется. Современные методы мечения клеток, такие как декстран, лизированный родамином, и витальный краситель diI также были разработаны для временной маркировки линий нервного гребня. [6]
Система маркировки перепелок-цыплят, разработанная Николь Ле Дуарен в 1969 году, была еще одним инструментальным методом, используемым для отслеживания клеток нервного гребня. [8] [9] Химеры , возникшие в результате трансплантации, позволили исследователям отличить клетки нервного гребня одного вида от окружающей ткани другого вида. С помощью этого метода поколения ученых смогли надежно отметить и изучить онтогенез клеток нервного гребня.
Индукция
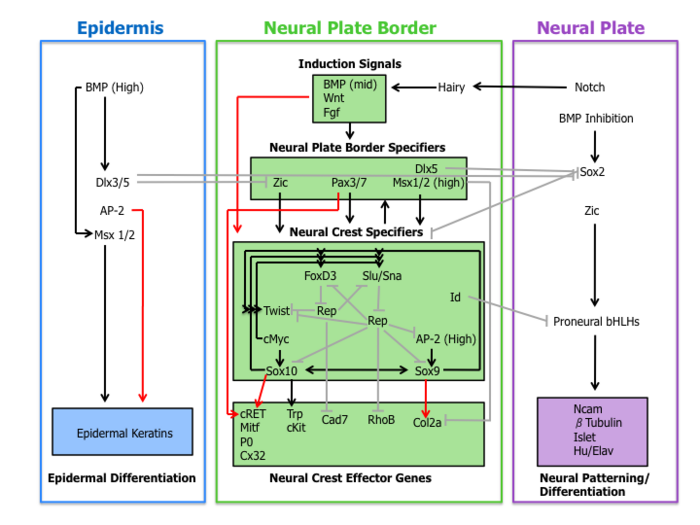
[ редактировать ]Молекулярный каскад событий участвует в установлении миграционных и мультипотентных характеристик клеток нервного гребня. Эту сеть регуляции генов можно разделить на следующие четыре подсети, описанные ниже.
Индуктивные сигналы
[ редактировать ]Во-первых, внеклеточные сигнальные молекулы, секретируемые из прилежащего эпидермиса и подлежащей мезодермы, такие как Wnts , BMPs и Fgfs, отделяют ненейральную эктодерму (эпидермис) от нервной пластинки во время нервной индукции . [1] [4]
Передача сигналов Wnt была продемонстрирована при индукции нервного гребня у нескольких видов посредством экспериментов по приобретению и потере функции. В соответствии с этим наблюдением, промоторная область слизняка (ген, специфичный для нервного гребня) содержит сайт связывания для транскрипционных факторов, участвующих в активации Wnt-зависимых генов-мишеней, что указывает на прямую роль передачи сигналов Wnt в спецификации нервного гребня. [10]
Современная роль BMP в формировании нервного гребня связана с индукцией нервной пластинки. Антагонисты BMP, диффундирующие из эктодермы, создают градиент активности BMP. Таким образом, линия нервного гребня формируется из промежуточных уровней передачи сигналов BMP, необходимых для развития нервной пластинки (низкий BMP) и эпидермиса (высокий BMP). [1]
Fgf из параксиальной мезодермы был предложен в качестве источника индуктивного сигнала нервного гребня. Исследователи продемонстрировали, что экспрессия доминантно-негативного рецептора Fgf в эксплантатах эктодермы блокирует индукцию нервного гребня при рекомбинации с параксиальной мезодермой. [11] Понимание роли путей BMP, Wnt и Fgf в экспрессии спецификаторов нервного гребня остается неполным.
Спецификаторы границ нервной пластинки
[ редактировать ]Сигнальные события, которые устанавливают границу нервной пластинки, приводят к экспрессии набора транскрипционных факторов, описанных здесь как спецификаторы границы нервной пластинки. Эти молекулы включают факторы Zic, Pax3/7, Dlx5, Msx1/2, которые могут опосредовать влияние Wnts, BMP и Fgfs. Эти гены широко экспрессируются в пограничной области нервной пластинки и предшествуют экспрессии достоверных маркеров нервного гребня. [4]
Экспериментальные данные помещают эти факторы транскрипции выше спецификаторов нервного гребня. Например, у Xenopus Msx1 необходим и достаточен для экспрессии Slug, Snail и FoxD3. [12] Более того, Pax3 необходим для экспрессии FoxD3 в эмбрионах мышей. [13]
Спецификаторы нервного гребня
[ редактировать ]За экспрессией спецификаторов границ нервной пластинки следует набор генов, включая Slug/Snail, FoxD3, Sox10, Sox9, AP-2 и c-Myc. Этот набор генов, обозначенный здесь как спецификаторы нервного гребня, активируется в возникающих клетках нервного гребня. По крайней мере, у Xenopus каждый спецификатор нервного гребня необходим и/или достаточен для экспрессии всех других спецификаторов, демонстрируя существование обширной перекрестной регуляции. [4] Более того, этот модельный организм сыграл важную роль в выяснении роли сигнального пути Hedgehog в спецификации нервного гребня, при этом транскрипционный фактор Gli2 играет ключевую роль. [14]
За пределами жестко регулируемой сети спецификаторов нервного гребня находятся два других фактора транскрипции Twist и Id. Twist, транскрипционный фактор bHLH , необходим для мезенхимной дифференцировки структур глоточной дуги . [15] Id является непосредственной мишенью c-Myc и, как известно, важен для поддержания стволовых клеток нервного гребня. [16]
Эффекторные гены нервного гребня
[ редактировать ]Наконец, спецификаторы нервного гребня включают экспрессию эффекторных генов, которые придают определенные свойства, такие как миграция и мультипотентность. Два эффектора нервного гребня, Rho GTPases и кадгерины , действуют при расслаивании, регулируя морфологию клеток и адгезивные свойства. Sox9 и Sox10 регулируют дифференцировку нервного гребня путем активации многих эффекторов, специфичных для типа клеток, включая Mitf, P0, Cx32, Trp и cKit. [4]
Миграция
[ редактировать ]
Миграция клеток нервного гребня включает высокоскоординированный каскад событий, который начинается с закрытия дорсальной нервной трубки .
Расслаивание
[ редактировать ]После слияния нервных складок с образованием нервной трубки клетки, первоначально расположенные на границе нервной пластинки, нервного гребня становятся клетками . [17] Чтобы миграция началась, клетки нервного гребня должны подвергнуться процессу, называемому расслаиванием, который включает полный или частичный эпителиально-мезенхимальный переход (ЕМТ) . [18] Расслоение определяется как разделение ткани на различные популяции, в данном случае клетки нервного гребня отделяются от окружающей ткани. [19] И наоборот, ЭМП представляет собой серию событий, координирующих изменение эпителиального на мезенхимальный фенотипа . [18] Например, деламинация у куриных эмбрионов запускается каскадом / Wnt , BMP который индуцирует экспрессию EMT, способствующую транскрипционным факторам, таким как SNAI2 и FoxD3 . [19] Хотя все клетки нервного гребня подвергаются ЭМП, время расслаивания происходит на разных стадиях у разных организмов: у эмбрионов Xenopus laevis происходит массивное расслоение, которое происходит, когда нервная пластинка не полностью срослась, тогда как у куриного эмбриона расслаивание происходит во время слияния нервных пластинок. нервная складка . [19]
Перед расслоением презумптивные клетки нервного гребня первоначально прикрепляются к соседним клеткам с помощью белков плотного соединения, таких как окклюдин , и молекул клеточной адгезии, таких как NCAM и N - кадгерин . [20] Дорсально экспрессируемые BMPs инициируют расслоение, индуцируя экспрессию цинкового пальца факторов транскрипции белка улитки , слизняка и твиста . [17] Эти факторы играют непосредственную роль в индукции эпителиально-мезенхимального перехода за счет снижения экспрессии окклюдина и N-кадгерина а также способствуют модификации NCAM , остатками полисиаловой кислоты для уменьшения адгезивности. [17] [21] Клетки нервного гребня также начинают экспрессировать протеазы, способные разрушать кадгерины, такие как ADAM10. [22] и секретируют матриксные металлопротеиназы (ММП), которые разрушают вышележащую базальную пластинку нервной трубки, позволяя клеткам нервного гребня выйти наружу. [20] Кроме того, клетки нервного гребня во время миграции начинают экспрессировать интегрины , которые связываются с внеклеточного матрикса белками , включая коллаген , фибронектин и ламинин . [23] Как только базальная пластинка становится проницаемой, клетки нервного гребня могут начать мигрировать по эмбриону.
Миграция
[ редактировать ]
Миграция клеток нервного гребня происходит в ростральном направлении к каудальному без необходимости использования нейронного каркаса, например, вдоль радиальных глиальных клеток . По этой причине процесс миграции клеток гребня называется «свободной миграцией». Вместо каркаса на клетках-предшественниках миграция нервного гребня является результатом отталкивающего направления через EphB / EphrinB и передачу сигналов семафорина / нейропилина , взаимодействия с внеклеточным матриксом и контактного ингибирования друг с другом. [17] В то время как белки эфрин и Eph обладают способностью передавать двунаправленный сигнал, отталкивание клеток нервного гребня использует преимущественно прямую передачу сигналов, чтобы инициировать ответ внутри клетки нервного гребня, несущей рецептор . [23] Растущие клетки нервного гребня экспрессируют EphB, рецепторную тирозинкиназу , которая связывает трансмембранный лиганд EphrinB, экспрессируемый в каудальной половине каждого сомита . Когда эти два домена взаимодействуют, это вызывает фосфорилирование тирозина рецептора, активацию rhoGTPase и возможные цитоскелетные перестройки внутри клеток гребня, вызывающие их отталкивание. Это явление позволяет клеткам нервного гребня проходить через ростральную часть каждого сомита. [17]
Передача отталкивающих сигналов семафорин-нейропилин работает синергетически с передачей сигналов EphB, направляя клетки нервного гребня вниз по ростральной половине сомитов у мышей. У куриных эмбрионов семафорин действует в головной области, направляя клетки нервного гребня через глоточные дуги . Помимо отталкивающей передачи сигналов, клетки нервного гребня экспрессируют интегрины β1 и α4 , которые обеспечивают связывание и управляемое взаимодействие с коллагеном , ламинином и фибронектином внеклеточного матрикса во время их перемещения. Кроме того, клетки гребня обладают внутренним ингибированием контакта друг с другом, свободно проникая в ткани различного происхождения, такие как мезодерма . [17] Клетки нервного гребня, мигрирующие через ростральную половину сомитов, дифференцируются в сенсорные и симпатические нейроны периферической нервной системы . Другой основной путь, по которому идут клетки нервного гребня, — дорсолатеральный между эпидермисом и дермамиотомом . дифференцируются в пигментные клетки дермы Клетки, мигрирующие по этому пути , . Дальнейшая дифференцировка клеток нервного гребня и спецификация их окончательного типа клеток смещены в результате их пространственно-временной подчиненности морфогенным сигналам, таким как BMP, Wnt, FGF, Hox и Notch . [20]
Клиническое значение
[ редактировать ]Нейрокристопатии возникают в результате аномальной спецификации, миграции, дифференцировки или гибели клеток нервного гребня на протяжении эмбрионального развития. [24] [25] Эта группа заболеваний включает широкий спектр врожденных пороков развития, поражающих многих новорожденных. Кроме того, они возникают из-за генетических дефектов, влияющих на формирование нервного гребня, и из-за действия тератогенов. [26]
синдром Ваарденбурга
[ редактировать ]Синдром Ваарденбурга — нейрокристопатия , возникающая в результате нарушения миграции клеток нервного гребня. Основные характеристики этого заболевания включают пегие глаза и врожденную глухоту . В случае пегости бесцветные участки кожи вызваны полным отсутствием пигмент вырабатывающих меланоцитов, нервного гребня . [27] Существует четыре различных типа синдрома Ваарденбурга, каждый из которых имеет различные генетические и физиологические особенности. Типы I и II различают в зависимости от того, страдают ли члены семьи больного человека антиутопией канторум . [28] Тип III приводит к аномалиям верхних конечностей. Наконец, тип IV также известен как синдром Ваарденбурга-Шаха, и у больных наблюдаются как синдром Ваарденбурга, так и болезнь Гиршпрунга . [29] Типы I и III наследуются по аутосомно-доминантному типу. [27] тогда как II и IV имеют аутосомно-рецессивный тип наследования. В целом синдром Ваарденбурга встречается редко, его частота в США составляет ~ 2/100 000 человек. Все расы и полы страдают в равной степени. [27] В настоящее время не существует лекарства или лечения синдрома Ваарденбурга.
Болезнь Гиршпрунга
[ редактировать ]К дефектам, связанным с развитием и миграцией клеток нервного гребня, также причастна болезнь Гиршпрунга (HD или HSCR), характеризующаяся отсутствием иннервации в участках кишечника . Отсутствие иннервации может привести к дальнейшим физиологическим отклонениям, таким как увеличение толстой кишки ( мегаколон ), непроходимость кишечника или даже замедление роста. При здоровом развитии клетки нервного гребня мигрируют в кишечник и образуют кишечные ганглии . Гены, играющие роль в здоровой миграции этих клеток нервного гребня в кишечник, включают RET , GDNF , GFRα , EDN3 и EDNRB . RET , рецепторная тирозинкиназа (RTK), образует комплекс с GDNF и GFRα . EDN3 и EDNRB тогда участвуют в одной и той же сигнальной сети. Когда эта передача сигналов нарушается у мышей, возникает аганглионоз или отсутствие этих кишечных ганглиев. [30]
Расстройство алкогольного спектра плода
[ редактировать ]Пренатальное воздействие алкоголя (ПАЭ) является одной из наиболее частых причин пороков развития . [31] В зависимости от степени воздействия и тяжести возникающих нарушений у пациентов диагностируется ряд расстройств, широко называемых расстройством фетального алкогольного спектра (ФАСН). нервного гребня Тяжелая форма ФАСН может нарушать миграцию , о чем свидетельствуют характерные черепно-лицевые аномалии , включая короткие глазные щели , удлиненную верхнюю губу и сглаженный желобок . Однако из-за беспорядочного характера этанола связывания механизмы возникновения этих нарушений до сих пор неясны. клеточных культур Эксплантаты клеток нервного гребня, а также in vivo развивающиеся рыбок данио, эмбрионы подвергнутые воздействию этанола, демонстрируют уменьшенное количество мигрирующих клеток и уменьшение расстояний, которые проходят мигрирующие клетки нервного гребня. Механизмы этих изменений не совсем понятны, но данные свидетельствуют о том, что ПАЭ может усиливать апоптоз из-за повышения уровня цитозольного кальция , вызванного IP3 -опосредованным высвобождением кальция из внутриклеточных депо. Было также высказано предположение, что снижение жизнеспособности клеток нервного гребня, подвергшихся воздействию этанола, вызвано увеличением окислительный стресс . Несмотря на эти и другие достижения, еще многое предстоит выяснить о том, как этанол влияет на развитие нервного гребня. Например, оказывается, что этанол по-разному влияет на одни клетки нервного гребня по сравнению с другими; то есть, хотя черепно-лицевые аномалии часто встречаются при ПАЭ, пигментные клетки нервного гребня поражаются минимально. [32]
Синдром ДиДжорджа
[ редактировать ]Синдром ДиДжорджа связан с делециями или транслокациями небольшого сегмента хромосомы человека 22 . Эта делеция может нарушить ростральных клеток миграцию или развитие нервного гребня . Некоторые наблюдаемые дефекты связаны с системой глоточного мешка , которая получает вклад от клеток рострального мигрирующего гребня. Симптомы дефекты синдрома ДиДжорджа включают врожденные пороки сердца , лица , а также некоторые неврологические нарушения и нарушения обучаемости . Сообщалось также, что у пациентов с делецией 22q11 чаще встречается шизофрения и биполярное расстройство . [33]
Синдром Тричера Коллинза
[ редактировать ]Синдром Тричера-Коллинза (TCS) возникает в результате нарушения развития первой и второй глоточных дуг на ранней эмбриональной стадии, что в конечном итоге приводит к аномалиям средней и нижней части лица. TCS вызван миссенс-мутацией гена TCOF1 , которая заставляет клетки нервного гребня подвергаться апоптозу во время эмбриогенеза . Хотя мутации гена TCOF1 являются одними из наиболее хорошо изученных по своей роли при СТО, мутации в POLR1C и POLR1D генах также связаны с патогенезом СТО. [34]
Клеточные линии
[ редактировать ]Клетки нервного гребня, возникающие из разных положений по передне -задней оси, развиваются в различные ткани. Эти области нервного гребня можно разделить на четыре основных функциональных домена, которые включают краниальный нервный гребень, туловищный нервный гребень, блуждающий и крестцовый нервный гребень и сердечный нервный гребень.
Краниальный нервный гребень
[ редактировать ]Краниальный нервный гребень мигрирует дорсолатерально, образуя черепно-лицевую мезенхиму, которая дифференцируется в различные краниальные ганглии, черепно-лицевые хрящи и кости. [21] Эти клетки проникают в глоточные карманы и дуги, где вносят вклад в вилочковую железу , кости среднего уха и челюсти, а также в одонтобласты зачатков зубов. [35]
Нервный гребень туловища
[ редактировать ]Ствол нервного гребня дает начало двум популяциям клеток. [36] Одна группа клеток, которым суждено стать меланоцитами, мигрирует дорсолатерально в эктодерму по направлению к вентральной срединной линии. Вторая группа клеток мигрирует вентролатерально через переднюю часть каждого склеротома . Клетки, которые остаются в склеротоме, образуют ганглии дорсальных корешков , тогда как те, которые продолжаются вентрально, образуют симпатические ганглии, мозговое вещество надпочечников и нервы, окружающие аорту. [35]
Вагусный и крестцовый нервный гребень
[ редактировать ]Клетки блуждающего и сакрального нервного гребня развиваются в ганглии кишечной нервной системы и парасимпатические ганглии. [35]
Сердечный нервный гребень
[ редактировать ]Сердечный нервный гребень развивается в меланоциты, хрящи, соединительную ткань и нейроны некоторых глоточных дуг. Кроме того, этот домен дает начало таким областям сердца, как мышечно-соединительная ткань крупных артерий и части перегородки , которая отделяет малый круг кровообращения от аорты. [35] Согласно новому исследованию, полулунные клапаны сердца связаны с клетками нервного гребня. [37]
Эволюция
[ редактировать ]Ряд структур, отличающих позвоночных от других хордовых, образован из производных клеток нервного гребня. В своей теории «новой головы» Ганс и Норткат утверждают, что наличие нервного гребня было основой специфических особенностей позвоночных, таких как сенсорные ганглии и черепной скелет. Более того, появление этих особенностей сыграло решающую роль в эволюции позвоночных, поскольку позволило им вести хищнический образ жизни. [38] [39]
Однако признание нервного гребня инновацией позвоночных не означает, что он возник de novo . Вместо этого новые структуры часто возникают в результате модификации существующих программ регулирования развития. Например, программы регулирования могут быть изменены за счет привлечения новых вышестоящих регуляторов или за счет использования новых нижестоящих генов-мишеней, помещая таким образом существующие сети в новый контекст. [40] [41] Эта идея подтверждается данными гибридизации in situ , которые показывают сохранение спецификаторов границ нервной пластинки у протохордовых , что позволяет предположить, что часть сети предшественников нервного гребня присутствовала у общего предка хордовых. [5] У некоторых хордовых беспозвоночных, таких как оболочники, была идентифицирована линия клеток (меланоцитов), которые похожи на клетки нервного гребня у позвоночных. Это означает, что рудиментарный нервный гребень существовал у общего предка позвоночных и оболочников. [42]
Производные нервного гребня
[ редактировать ]Эктомезенхима (также известная как мезэктодерма ): [43] одонтобласты , зубные сосочки , хондрокраниум ( носовая капсула , меккелев хрящ , склеральные косточки , квадратная, суставная, подъязычная и колумелла), трахеальный и гортанный хрящи, дерматокраний (перепончатые кости), спинные плавники и пластрон черепахи (низшие позвоночные), перициты и гладкие мышцы жаберных артерий и вен, сухожилия глазных и жевательных мышц, соединительная ткань желез головы и шеи (гипофиз, слюнные, слезные, тимус, щитовидная железа), дерма и жировая ткань свода черепа, вентральной части шеи и лица.
Эндокринные клетки : хромаффинные клетки мозгового слоя надпочечников, гломусные клетки I/II типа.
Периферическая нервная система : Сенсорные нейроны и глия ганглиев дорсальных корешков , головных ганглиев (VII и частично, V, IX и X), клеток Рохона-Борода , некоторых клеток Меркеля в усе, [44] [45] Сателлитные глиальные клетки всех вегетативных и сенсорных ганглиев, шванновские клетки всех периферических нервов.
Кишечные клетки : Энтерохромаффинные клетки . [46]
Меланоциты , мышечные и пигментные клетки радужной оболочки и даже ассоциированы с некоторыми опухолями (например, меланотической нейроэктодермальной опухолью младенчества ).
См. также
[ редактировать ]- Синдром первой дуги
- DGCR2 — может контролировать миграцию клеток нервного гребня.
- Список типов клеток человека, полученных из зародышевых листков
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Хуанг, X.; Сен-Жанне, JP (2004). «Индукция нервного гребня и возможности жизни на грани». Дев. Биол . 275 (1): 1–11. дои : 10.1016/j.ydbio.2004.07.033 . ПМИД 15464568 .
- ^ Шахова, Ольга; Соммер, Лукас (2008). «Стволовые клетки, полученные из нервного гребня» . СтемБук . Гарвардский институт стволовых клеток. дои : 10.3824/stembook.1.51.1 . ПМИД 20614636 . Проверено 27 декабря 2019 г.
- ^ Брукер, RJ 2014, Биология, 3-е изд., McGraw-Hill, Нью-Йорк, Нью-Йорк, 1084.
- ^ Jump up to: а б с д и Меулеманс, Д.; Броннер-Фрейзер, М. (2004). «Генно-регуляторные взаимодействия в эволюции и развитии нервного гребня» . Ячейка разработчиков . 7 (3): 291–9. дои : 10.1016/j.devcel.2004.08.007 . ПМИД 15363405 .
- ^ Jump up to: а б Саука-Шпенглер, Т.; Меулеманс, Д.; Джонс, М.; Броннер-Фрейзер, М. (2007). «Древнее эволюционное происхождение регуляторной сети генов нервного гребня» . Ячейка разработчиков . 13 (3): 405–20. дои : 10.1016/j.devcel.2007.08.005 . ПМИД 17765683 .
- ^ Jump up to: а б Ле Дуарен, Нью-Мексико (2004). «Птичий эмбрион как модель для изучения развития нервного гребня: долгая и продолжающаяся история» . Мех. Дев . 121 (9): 1089–102. дои : 10.1016/j.mod.2004.06.003 . ПМИД 15296974 .
- ^ Хёрстадиус , С. (1950). Нервный гребень: его свойства и производные в свете экспериментальных исследований . Издательство Оксфордского университета, Лондон, 111 стр.
- ^ Ле Дуарен, Нью-Мексико (1969). интерфазного ядра японского перепела (Coturnix coturnix japonica). Использование этих особенностей в качестве «биологической маркировки» в исследованиях тканевых взаимодействий и миграции клеток во время онтогенеза» Bull Belg . « Особенности . Fr Biol .
- ^ Ле Дуарен, Нью-Мексико (1973). «Методика биологической маркировки клеток и ее использование в экспериментальной эмбриологии». Дев Биол . 30 (1): 217–22. дои : 10.1016/0012-1606(73)90061-4 . ПМИД 4121410 .
- ^ Валлин, Дж.; и др. (2001). «Клонирование и характеристика трех промоторов слизней Xenopus выявили прямую регуляцию посредством передачи сигналов Lef/бета-катенин» . J Биол Хим . 276 (32): 30350–8. дои : 10.1074/jbc.M103167200 . ПМИД 11402039 .
- ^ Мэр Р.; Герреро, Н.; Мартинес, К. (1997). «Роль FGF и ноггина в индукции нервного гребня» . Дев Биол . 189 (1): 1–12. дои : 10.1006/dbio.1997.8634 . ПМИД 9281332 .
- ^ Трибуло, К.; и др. (2003). «Регуляция генов Msx с помощью градиента Bmp важна для спецификации нервного гребня» . Разработка . 130 (26): 6441–52. дои : 10.1242/dev.00878 . hdl : 11336/95313 . ПМИД 14627721 .
- ^ Доттори, М.; Гросс, МК; Лабоски, П.; Гулдинг, М. (2001). «Фактор транскрипции крылатой спирали Foxd3 подавляет дифференцировку межнейронов и способствует судьбе клеток нервного гребня». Разработка . 128 (21): 4127–4138. дои : 10.1242/dev.128.21.4127 . ПМИД 11684651 .
- ^ Серрисуэла, Сантьяго; Вега-Лопес, Гильермо А.; Паласио, Мария Белен; Трибуло, Селеста; Айбар, Мануэль Дж. (01 декабря 2018 г.). «Gli2 необходим для индукции и миграции нервного гребня Xenopus laevis» . Механизмы развития . 154 : 219–239. дои : 10.1016/j.mod.2018.07.010 . hdl : 11336/101714 . ISSN 0925-4773 . ПМИД 30086335 .
- ^ Винцентц, JW; и др. (2008). «Отсутствие Twist1 приводит к аберрантному морфогенезу сердечного нервного гребня» . Дев Биол . 320 (1): 131–9. дои : 10.1016/j.ydbio.2008.04.037 . ПМК 2572997 . ПМИД 18539270 .
- ^ Лайт, В.; и др. (2005). «Xenopus Id3 необходим ниже Myc для образования мультипотентных клеток-предшественников нервного гребня» . Разработка . 132 (8): 1831–41. дои : 10.1242/dev.01734 . ПМИД 15772131 .
- ^ Jump up to: а б с д и ж Санес, Дэн (2012). Развитие нервной системы, 3-е изд . Оксфорд: ELSEVIER INC., стр. 70–72. ISBN 978-0123745392 .
- ^ Jump up to: а б Ламуй, Сами (2014). «Молекулярные механизмы эпителиально-мезенхимального перехода» . Nature Reviews Молекулярно-клеточная биология . 15 (3): 178–196. дои : 10.1038/nrm3758 . ПМК 4240281 . ПМИД 24556840 .
- ^ Jump up to: а б с Тевено, Эрик (2012). «Расслоение и миграция нервного гребня: от перехода эпителия к мезенхиме к коллективной миграции клеток» (PDF) . Биология развития . 366 (1): 34–54. дои : 10.1016/j.ydbio.2011.12.041 . ПМИД 22261150 .
- ^ Jump up to: а б с Кандел, Эрик (2013). Принципы нейронауки . Нью-Йорк: The McGraw-Hill Companies, Inc., стр. 1197–1199. ISBN 978-0-07-139011-8 .
- ^ Jump up to: а б Тэнейхилл, Луизиана (2008). «Придерживаться или не придерживаться: роль кадгеринов в развитии нервного гребня». Cell Adh Миграция. 2, 223–30.
- ^ Мэр Роберто (2013 г.). «Нейральный гребень» . Разработка . 140 (11): 2247–2251. дои : 10.1242/dev.091751 . ПМИД 23674598 .
- ^ Jump up to: а б Сакука-Шпенглер, Татьяна (2008). «Сеть регуляции генов управляет формированием нервного гребня». Nature Reviews Молекулярно-клеточная биология . 9 (7): 557–568. дои : 10.1038/nrm2428 . ПМИД 18523435 . S2CID 10746234 .
- ^ Вега-Лопес, Гильермо А.; Серрисуэла, Сантьяго; Трибуло, Селеста; Айбар, Мануэль Дж. (01 декабря 2018 г.). «Нейрокристопатии: новые открытия спустя 150 лет после открытия нервного гребня» . Биология развития . Нейронный гребень: 150 лет после открытия Хиса. 444 : S110–S143. дои : 10.1016/j.ydbio.2018.05.013 . hdl : 11336/101713 . ISSN 0012-1606 . ПМИД 29802835 .
- ^ Боланде, Роберт П. (1 июля 1974 г.). «Нейрокристопатии: объединяющая концепция заболеваний, возникающих при неправильном развитии нервного гребня». Патология человека . 5 (4): 409–429. дои : 10.1016/S0046-8177(74)80021-3 . ISSN 0046-8177 .
- ^ Серрисуэла, Сантьяго; Вега-Лопес, Гильермо А.; Айбар, Мануэль Дж. (11 января 2020 г.). «Роль тератогенов в развитии нервного гребня». Исследование врожденных дефектов . 112 (8): 584–632. дои : 10.1002/bdr2.1644 . ISSN 2472-1727 . ПМИД 31926062 . S2CID 210151171 .
- ^ Jump up to: а б с Мэллори, SB; Винер, Э; Нордлунд, Джей-Джей (1986). «Синдром Ваарденбурга с болезнью Гиршпрунга: дефект нервного гребня». Детская дерматология . 3 (2): 119–124. дои : 10.1111/j.1525-1470.1986.tb00501.x . ПМИД 3952027 . S2CID 23858201 .
- ^ Ариас, С (1971). «Генетическая гетерогенность при синдроме Ваарденбурга». Врожденные дефекты Б. 07 (4): 87–101. ПМИД 5006208 .
- ^ «Синдром Ваарденбурга». Домашний справочник по генетике. Октябрь 2012.
- ^ Роджерс, Дж. М. (2016). «Поиск недостающих lnc: генные регуляторные сети в развитии нервного гребня и длинные некодирующие РНК-биомаркеры болезни Гиршпрунга». Нейрогастроэнтерол Мотил . 28 (2): 161–166. дои : 10.1111/nmo.12776 . ПМИД 26806097 . S2CID 12394126 .
- ^ Сэмпсон, PD; Стрейссгут, AP; Букштейн, Флорида; Литтл, Р.Э.; Кларен, СК; Деэн, П.; Грэм, Дж. М. младший (1997). «Заболеваемость фетального алкогольного синдрома и распространенность связанных с алкоголем нарушений нервного развития» . Тератология . 56 (5): 317–326. doi : 10.1002/(SICI)1096-9926(199711)56:5<317::AID-TERA5>3.0.CO;2-U . ПМИД 9451756 .
- ^ Смит, С.М.; Гарик, А.; Флентке, Г. Р.; Беррес, Мэн (2014). «Развитие нервного гребня при алкогольном синдроме плода» . Исследование врожденных дефектов, часть C: Эмбрион сегодня: обзоры . 102 (3): 210–220. дои : 10.1002/bdrc.21078 . ПМК 4827602 . ПМИД 25219761 .
- ^ Мошенник, Питер Дж. (2000). «Синдром делеции 22q11» . Молекулярная генетика человека . 9 (16): 2421–2426. дои : 10.1093/hmg/9.16.2421 . ПМИД 11005797 .
- ^ Ахмед, М.; Йе, Х.; Тауб, П. (2016). «Обзор генетической основы пороков развития челюстей» . Журнал детской генетики . 05 (4): 209–219. дои : 10.1055/s-0036-1593505 . ПМК 5123890 . ПМИД 27895973 .
- ^ Jump up to: а б с д Гилберт, Скотт Ф. (2000). Нервный гребень . Синауэр Ассошиэйтс.
- ^ Вега-Лопес, Гильермо А.; Серрисуэла, Сантьяго; Айбар, Мануэль Дж. (2017). «Туловищные клетки нервного гребня: формирование, миграция и не только» . Международный журнал биологии развития . 61 (1–2): 5–15. дои : 10.1387/ijdb.160408gv . hdl : 11336/53692 . ISSN 0214-6282 . ПМИД 28287247 .
- ^ Такамура, Казуши; Окишима, Такахиро; Одо, Сёдзо; Хаякава, Кунио (1990). «Связь клеток цефалического нервного гребня с развитием сердечно-сосудистой системы, особенно полулунных клапанов». Анатомия и эмбриология . 182 (3): 263–72. дои : 10.1007/BF00185519 . ПМИД 2268069 . S2CID 32986727 .
- ^ Ганс, К.; Норткатт, Р.Г. (1983). «Нервный гребень и происхождение позвоночных: новая голова». Наука . 220 (4594): 268–274. Бибкод : 1983Sci...220..268G . дои : 10.1126/science.220.4594.268 . ПМИД 17732898 . S2CID 39290007 .
- ^ Норткатт, Гленн (2005). «Пересмотр гипотезы новой головы». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 304Б (4): 274–297. дои : 10.1002/jez.b.21063 . ПМИД 16003768 .
- ^ Саука-Шпенглер, Т.; Броннер-Фрейзер, М. (2006). «Развитие и эволюция мигрирующего нервного гребня: взгляд на регуляцию генов». Curr Opin Genet Dev . 13 (4): 360–6. дои : 10.1016/j.где.2006.06.006 . ПМИД 16793256 .
- ^ Донохью, ПК; Грэм, А.; Келш, Р.Н. (2008). «Происхождение и эволюция нервного гребня» . Биоэссе . 30 (6): 530–41. doi : 10.1002/bies.20767 . ПМК 2692079 . ПМИД 18478530 .
- ^ Абитуа, ПБ; Вагнер, Э.; Наваррете, Айова; Левин, М. (2012). «Идентификация рудиментарного нервного гребня у хордовых беспозвоночных» . Природа . 492 (7427): 104–107. Бибкод : 2012Natur.492..104A . дои : 10.1038/nature11589 . ПМЦ 4257486 . ПМИД 23135395 .
- ^ Кальхайм, К. и Ле Дуарен, Нью-Мексико (1998). Нервный гребень (2-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета.
- ^ Ван Кеймюлен, А; Маскр, Дж; Юсефф, К.К.; и др. (октябрь 2009 г.). «Эпидермальные предшественники дают начало клеткам Меркеля во время эмбрионального развития и гомеостаза взрослых» . Дж. Клеточная Биол . 187 (1): 91–100. дои : 10.1083/jcb.200907080 . ПМК 2762088 . ПМИД 19786578 .
- ^ Седер, В; Грим, М; Халата, З; Зибер-Блюм, М. (январь 2003 г.). «Происхождение нервного гребня клеток Меркеля млекопитающих» . Дев. Биол . 253 (2): 258–63. дои : 10.1016/s0012-1606(02)00015-5 . ПМИД 12645929 .
- ^ Лейк, Дж.И.; Хейкерот, РОД (1 июля 2013 г.). «Развитие кишечной нервной системы: миграция, дифференциация и болезни» . Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 305 (1): G1–24. дои : 10.1152/ajpgi.00452.2012 . ПМЦ 3725693 . ПМИД 23639815 .