Фотолиаза
| Криптохром/фотолиаза, С-конец, связывание FAD | |||
|---|---|---|---|
 Деазафлавин-фотолиаза из Anacystis nidulans , иллюстрирующая два светособирающих кофактора: FADH. − (желтый) и 8-HDF (голубой). | |||
| Идентификаторы | |||
| Символ | FAD_binding_7 | ||
| Пфам | PF03441 | ||
| ИнтерПро | ИПР005101 | ||
| PROSITE | PDOC00331 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1qnf / SCOPe / СУПФАМ | ||
| |||
| дезоксирибодипиримидин фотолиаза (CPD) | |||
|---|---|---|---|
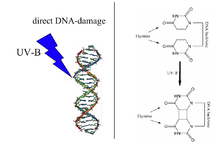 Индуцированный УФ-излучением тимин-тиминциклобутановый димер (справа) представляет собой тип повреждения ДНК , которое восстанавливается ДНК-фотолиазой. Примечание. На приведенной выше диаграмме неправильно обозначен тимин, поскольку в структурах отсутствуют 5-метильные группы. | |||
| Идентификаторы | |||
| Номер ЕС. | 4.1.99.3 | ||
| Номер CAS. | 37290-70-3 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Фотолиазы ( EC 4.1.99.3 ) представляют собой репарации ДНК ферменты , которые восстанавливают повреждения, вызванные воздействием ультрафиолетового света. Этим ферментам требуется видимый свет (из фиолетового/синего конца спектра) как для собственной активации, так и для их собственной активации. [ 1 ] и для фактического восстановления ДНК. [ 2 ] Механизм репарации ДНК с участием фотолиаз называется фотореактивацией. В основном они превращают димеры пиримидина в нормальную пару пиримидиновых оснований. Фотореактивация, первый репарации ДНК , первоначально был описан Альбертом Кельнером в 1949 году. открытый механизм [ 3 ] и независимо Ренато Дульбекко также в 1949 году. [ 4 ] [ 5 ] [ 6 ]
Функция
[ редактировать ]Фотолиазы связывают комплементарные цепи ДНК и разрушают определенные типы пиримидиновых димеров , которые возникают, когда пара тиминовых или цитозиновых оснований на одной и той же цепи ДНК становится ковалентно связанной. Длина связи этой димеризации короче, чем длина связи нормальной структуры B-ДНК, которая создает неправильную матрицу для репликации и транскрипции. [ 7 ] Более распространенная ковалентная связь включает образование циклобутанового мостика. Фотолиазы обладают высоким сродством к этим повреждениям, обратимо связывают их и превращают обратно в исходные основания. Процесс репарации ДНК, катализируемый фотолиазой, при котором расщепляются димеры циклобутан-пиримидина, был изучен с помощью кристаллографии с временным разрешением и компьютерного анализа, позволяющего визуализировать процесс на атомном уровне. [ 8 ]
Эволюция
[ редактировать ]Фотолиаза — это филогенетически старый фермент, который присутствует и функционирует у многих видов, от бактерий до грибов и растений. [ 9 ] и животным . [ 10 ] Фотолиаза особенно важна для восстановления повреждений растений, вызванных УФ-излучением. Фотолиазный механизм больше не работает у людей и других плацентарных млекопитающих, которые вместо этого полагаются на менее эффективный механизм эксцизионной репарации нуклеотидов , хотя у них сохраняется много криптохромов . [ 11 ] Морозовый стресс у однолетней пшеницы Triticum aestivum и ее многолетнего родственника Thinopyrum intermedium сопровождается значительным увеличением экспрессии ДНК-фотолиаз. [ 12 ]
Фотолиазы представляют собой флавопротеины и содержат два светособирающих кофактора . Многие фотолиазы имеют N-концевой домен , который связывает второй кофактор. Все фотолиазы содержат двухэлектронно-восстановленную ФАДГ. − ; они разделены на два основных класса в зависимости от второго кофактора, которым может быть либо метенилтетрагидрофолат птерина (MTHF) в фолат-фотолиазах , либо деазафлавин- 8-гидрокси-7,8-дидеметил-5-деазарибофлавин (8-HDF) в деазафлавин-фотолиазах. . Хотя для каталитической активности необходим только ФАД, второй кофактор значительно ускоряет скорость реакции в условиях низкой освещенности. Фермент действует путем переноса электронов , при котором восстановленный флавин FADH − активируется энергией света и действует как донор электронов, разрушая димер пиримидина. [ 13 ]
На основании сходства последовательностей ДНК-фотолиазы можно разделить на несколько классов: [ 14 ] [ 15 ]
|
- Фотолиазы CPD класса 1 представляют собой ферменты, которые обрабатывают повреждения циклобутан-пиримидинового димера (CPD) грамотрицательных и грамположительных бактерий, а также галофильных архей Halobacterium halobium . [ 16 ]
- Фотолиазы CPD класса 2 также обрабатывают поражения CPD. Они встречаются в таких растениях, как кресс-салат Arabidopsis thaliana и рис .
- Криптохромы растений и грибов аналогичны CPD класса 1. Это фоторецепторы синего света, которые опосредуют экспрессию генов, индуцированную синим светом, и модуляцию циркадных ритмов .
- Лиазы CPD класса 3 составляют сестринскую группу криптохромов растений, которые, в свою очередь, являются сестринской группой CPD класса 1.
- Группа Cry-DASH представляет собой CPD-лиазы, высокоспецифичные к одноцепочечной ДНК. Членами являются Vibrio cholerae , X1Cry от Xenopus laevis и AtCry3 от Arabidopsis thaliana . [ 10 ] Первоначально DASH был назван в честь Drosophila , Arabidopsis , Synechocystis и Human — четырех таксонов, которые первоначально считались носителями этого семейства лиаз. С тех пор классификация изменилась. Часть их имени «Крик» возникла из-за первоначальных предположений, что они были криптохромами. [ 14 ]
- Эукариотические (6-4)ДНК-фотолиазы образуют группу с криптохромами животных, которые контролируют циркадные ритмы. Они встречаются у различных видов, включая дрозофилу и человека. Криптохромы имеют свою детальную группировку. [ 15 ]
- Бактериальные лиазы 6-4 ( InterPro : IPR007357 ), также известные как группа FeS-BCP, образуют собственную внешнюю группу по отношению ко всем фотолиазам.
В некоторых системах, таких как PRINTS (PR00147), ветвь CPD, не относящаяся к классу 2, имеет тенденцию группироваться в класс 1. Хотя члены более мелких групп согласованы, филогения может сильно различаться среди авторов из-за различий в методологии, что приводит к некоторой путанице с авторами, которые пытаются уместить все (за исключением FeS-BCP) в классификацию двух классов. [ 15 ] Криптохромы образуют полифилетическую группу, включающую фотолиазы, которые утратили активность по восстановлению ДНК и вместо этого контролируют циркадные ритмы. [ 14 ] [ 15 ]
Приложение
[ редактировать ]Добавление фотолиазы из сине-зеленых водорослей Anacystis nidulans к клеткам HeLa частично уменьшило повреждение ДНК от воздействия UVB. [ 17 ]
Белки человека, содержащие этот домен
[ редактировать ]Номенклатура
[ редактировать ]Систематическое название этого класса ферментов — дезоксирибоциклобутадипиримидинпиримидинлиаза . Другие широко используемые названия включают фотореактивирующий фермент , ДНК-фотолиазу , ДНК-фотореактивирующий фермент , ДНК-циклобутандипиримидинфотолиазу , ДНК-фотолиазу , дезоксирибонуклеиновую фотолиазу , дезоксирибодипиримидинфотолиазу , фотолиазу , PRE , фотолиазу PhrB , дезоксирибонуклеиновую циклобутандипиримидинфотолиазу , Phr A фотолиаза , дипиримидинфотолиаза (фоточувствительная) и дезоксирибонуклеатпиримидиндимерлиаза (фоточувствительная) . Этот фермент принадлежит к семейству лиаз , а именно к универсальному классу углерод-углеродных лиаз.
Ссылки
[ редактировать ]- ^ Ямамото Дж., Симидзу К., Канда Т., Хосокава Ю., Иваи С., Плаза П., Мюллер П. (октябрь 2017 г.). «Потеря четвертого переносящего электрон триптофана у животных (6-4) фотолиазы ухудшает активность восстановления ДНК в бактериальных клетках». Биохимия . 56 (40): 5356–64. doi : 10.1021/acs.biochem.7b00366 . ПМИД 28880077 .
- ^ Тиагараджан В., Бирдин М., Экер А.П., Мюллер П., Бреттель К. (июнь 2011 г.). «Кинетика расщепления димера циклобутан-тимина ДНК-фотолиазой, контролируемая непосредственно в УФ-излучении» . Proc Natl Acad Sci США . 108 (23): 9402–7. Бибкод : 2011PNAS..108.9402T . дои : 10.1073/pnas.1101026108 . ПМК 3111307 . ПМИД 21606324 .
- ^ Кельнер А. (февраль 1949 г.). «Влияние видимого света на восстановление Streptomyces Griseus Conidia после повреждений, вызванных ультрафиолетовым облучением» . Proc Natl Acad Sci США . 35 (2): 73–9. дои : 10.1073/pnas.35.2.73 . ПМЦ 1062964 . ПМИД 16588862 .
- ^ Дульбекко Р. (июнь 1949 г.). «Реактивация бактериофага, инактивированного ультрафиолетом, видимым светом». Природа . 163 (4155): 949. дои : 10.1038/163949b0 . ПМИД 18229246 .
- ^ Дульбекко Р. (март 1950 г.). «Опыты по фотореактивации бактериофагов, инактивированных ультрафиолетовым излучением» . J Бактериол . 59 (3): 329–47. дои : 10.1128/jb.59.3.329-347.1950 . ПМЦ 385765 . ПМИД 15436402 .
- ^ Фридберг ЕС (сентябрь 2015 г.). «История репарации ДНК и мутагенеза: I. Открытие ферментативной фотореактивации». Репарация ДНК (Амст) . 33 : 35–42. дои : 10.1016/j.dnarep.2015.06.007 . ПМИД 26151545 .
- ^ Гарретт Р.Х., Гришэм К.М. (2010). Биохимия . Брукс/Коул, Cengage Learning. ISBN 978-0-495-10935-8 . OCLC 984382855 .
- ^ Маэстре-Рейна М, Ван ПХ, Нанго Э, Хосокава Ю, Сафт М, Фуррер А, Ян CH, Густи Нгура Путу ЭП, Ву ВДж, Эммерих ХДж, Карамелло Н, Франц-Бадур С, Ян С, Энгильберге С, Враник М и др., Гловер Х.Л., Вайнерт Т., Ву HY, Ли CC, Хуан В.К., Хуан К.Ф., Чанг Ю.К., Ляо Дж.Х., Вэн Дж.Х., Гад В., Чанг К.В., Пан А.Х., Ян К.К., Лин В.Т., Чанг Ю.К., Гаши. Д , Бил Озеров Д, Насс К, Кнопп Г, Джонсон П.Дж., Сирелли С, Милн С, Баселлар С, Сугахара М, Овада С, Джоти Ю, Ямашита А, Танака Р, Танака Т, Луо Ф, Тоно К, Зажицка В , Мюллер П., Алахмад М.А., Безольд Ф., Фукс В., Гнау П., Кионтке С., Корф Л., Райтофер В., Рознер К.Дж., Зайлер Э.М., Ватад М., Верель Л., Спадаччини Р., Ямамото Дж., Ивата С., Чжун Д., Стендфусс Дж. 2010–2015 гг., Ройант А., Бесшо Ю., Эссен Л.О., Цай, доктор медицинских наук (декабрь 2023 г.). «Визуализация процесса восстановления ДНК с помощью фотолиазы с атомным разрешением». Наука 382 (6674): eadd7 дои : 10.1126/science.add7795 . ПМИД 38033054 .
- ^ Эраниши М., Накамура К., Мориока Х., Ямамото К., Хидема Дж. (2008). «Нативная циклобутанпиримидиндимерная фотолиаза риса фосфорилируется» . Физиология растений . 146 (4): 1941–51. дои : 10.1104/стр.107.110189 . ПМК 2287361 . ПМИД 18235036 .
- ^ Jump up to: а б Селби КП, Санкар А (ноябрь 2006 г.). «Класс ферментов криптохрома/фотолиазы с фотолиазной активностью, специфичной для одноцепочечной ДНК» . Proc Natl Acad Sci США . 103 (47): 17696–700. Бибкод : 2006PNAS..10317696S . дои : 10.1073/pnas.0607993103 . ПМК 1621107 . ПМИД 17062752 .
- ^ Лукас-Льедо Дж.И., Линч М. (май 2009 г.). «Эволюция частоты мутаций: филогеномный анализ семейства фотолиаз/криптохромов» . Молекулярная биология и эволюция . 26 (5): 1143–53. дои : 10.1093/molbev/msp029 . ПМЦ 2668831 . ПМИД 19228922 .
- ^ Джайкумар Н.С., Дорн К.М., Баас Д., Уилке Б., Капп С., Снапп СС (декабрь 2020 г.). «На повреждение нуклеиновой кислоты и восстановление ДНК влияет стресс замерзания у однолетней пшеницы ( Triticum aestivum ), а также возраст растения и замерзание у ее многолетнего родственника ( Thinopyrum intermedium )». Я Джей Бот . 107 (12): 1693–1709. дои : 10.1002/ajb2.1584 . ПМИД 33340368 .
- ^ Санджар А (июнь 2003 г.). «Структура и функция ДНК-фотолиазы и криптохромных фоторецепторов синего света». Химические обзоры . 103 (6): 2203–37. дои : 10.1021/cr0204348 . ПМИД 12797829 .
- ^ Jump up to: а б с д Ширер П., Чжан Ф., Калмс Дж., фон Штеттен Д., Краус Н., Оберпихлер И., Лампартер Т. (май 2015 г.). «Структура фотолиазы циклобутанпиримидиндимера III класса обнаруживает новый сайт связывания антенного хромофора и альтернативные пути фотовосстановления» . Журнал биологической химии . 290 (18): 11504–14. дои : 10.1074/jbc.M115.637868 . ПМЦ 4416854 . ПМИД 25784552 .
- ^ Jump up to: а б с д Ривера А.С., Озтюрк Н., Фэйи Б., Плашецки Д.К., Дегнан Б.М., Санкар А., Окли Т.Д. (апрель 2012 г.). «Восприимчивый к синему свету криптохром проявляется в губчатом глазу, лишенном нейронов и опсина» . Журнал экспериментальной биологии . 215 (Часть 8): 1278–86. дои : 10.1242/jeb.067140 . ПМК 3309880 . ПМИД 22442365 .
- ^ Маккриди С., Марчелло Л. (июнь 2003 г.). «Восстановление УФ-повреждений у Halobacterium salinarum ». Биохим Соц Транс . 31 (Часть 3): 694–8. дои : 10.1042/bst0310694 . ПМИД 12773185 .
- ^ Кульмс Д., Поппельманн Б., Ярош Д., Люгер Т.А., Крутманн Дж., Шварц Т. (июль 1999 г.). «Эффекты ядер и клеточных мембран независимо способствуют индукции апоптоза в клетках человека, подвергшихся воздействию УФВ-излучения» . Proc Natl Acad Sci США . 96 (14): 7974–9. Бибкод : 1999PNAS...96.7974K . дои : 10.1073/pnas.96.14.7974 . ПМК 22172 . ПМИД 10393932 .
Дальнейшее чтение
[ редактировать ]- Экер А.П., Фихтингер-Шепман А.М. (1975). «Исследование фотореактивирующего фермента ДНК из Streptomyces griseus II. Очистка фермента». Биохим. Биофиз. Акта . 378 (1): 54–63. дои : 10.1016/0005-2787(75)90136-7 . ПМИД 804322 .
- Санкар ГБ, Смит Ф.В., Рид Р., Пейн Дж., Леви М., Санкар А. (1987). «Механизм действия ДНК-фотолиазы Escherichia coli. I. Образование фермент-субстратного комплекса» . Ж. Биол. Хим . 262 (1): 478–85. дои : 10.1016/S0021-9258(19)75952-3 . ПМИД 3539939 .
- Сетлоу Дж. К., Боллум Ф. Дж. (1968). «Минимальный размер субстрата для фотореактивирующего фермента дрожжей». Биохим. Биофиз. Акта . 157 (2): 233–7. дои : 10.1016/0005-2787(68)90077-4 . ПМИД 5649902 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Фотолиазой, на Викискладе?
СМИ, связанные с Фотолиазой, на Викискладе?