Экстрахромосомная круговая ДНК
Экстрахромосомная круговая ДНК ( экцДНК ) представляет собой тип двухцепочечной круглой структуры ДНК, который был впервые обнаружен в 1964 году Аликс Бассель и Ясуо Хотта. [ 1 ] В отличие от ранее идентифицированных структур круглых ДНК (например, бактериальные плазмиды , митохондриальную ДНК , круглые бактериальные хромосомы или хлоропластская ДНК ), экцДНК являются круглыми ДНК в эукариотических ядрах растений и животных (включая человека). Экстрахромосомная круговая ДНК получена из хромосомной ДНК, может варьироваться в размере от 50 пар оснований до нескольких мега-пар длиной в длину и может кодировать регуляторные элементы и полноразмерные гены . ЭккДНК наблюдалась у различных эукариотических видов [ 2 ] [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ] и предполагается, что это побочный продукт запрограммированных событий рекомбинации ДНК , таких как рекомбинация V (D) J. [ 8 ] [ 9 ]
Исторический фон
[ редактировать ]В 1964 году Бассель и Хотта опубликовали свое первоначальное открытие экдны, которое они сделали, исследуя Франклина Шталя . хромосомную теорию [ 10 ] В своих экспериментах они визуализировали изолированные ядра пшеницы и кабану с помощью электронной микроскопии . [ 10 ] Их исследование показало, что кабаловые сперматозоиды содержали эккДНК различных размеров. [ 10 ] В 1965 году исследовательская группа Артура Сприггса идентифицировала экцДНК в образцах эмбриональных опухолей пяти педиатрических пациентов и бронхиальной карциномы одного взрослого пациента . [ 11 ] В последующие годы дополнительные исследования привели к открытию экцДНК в различных видах, перечисленных в таблице 1:
| Год | Организм | Ссылка |
|---|---|---|
| 1965 | Кабана сперма | Hotta and Bassel, 1965 [ 10 ] |
| 1965 | Опухоли человека | Cox et al., 1965 [ 11 ] |
| 1969 | Дрожжи | Billheimer и Avers, 1969 [ 12 ] |
| 1984 | Трипаносоматиды | Beverly et al., 1984 [ 13 ] |
| 1972 | Евглена | Насс и Бен-Шаул, 1972 [ 14 ] |
| 1972 | Табак | Wong and Wildman, 1972 [ 15 ] |
| 1972, 1978, 1980 | Грибы | Agsteribbe et al., 1972; [ 16 ] Stahl et al., 1978; [ 17 ] Lazarus et al., 1980 [ 18 ] |
| 1972, 1985 | Культурные человеческие фибробласты | Смит и Виноград, 1972; [ 19 ] Riabowol et al., 1985 [ 4 ] |
| 1976 | Xenopus | Доброе утро-Нарделли и др., 1976 [ 20 ] |
| 1978, 1984 | Цыпленка Бурса | Delap and Rush, 1978; [ 21 ] Тода и Ямагиши, 1984 [ 22 ] |
| 1982 | Человеческие ткани | Calabretta et al., 1982 [ 23 ] |
| 1983 | Эмбрион мыши | Yamagishi et al., 1983 [ 24 ] |
| 1983, 1988, 1990 | Мышиные ткани | Tsuda et al., 1983; [ 25 ] Flores et al., 1988; [ 26 ] Gaubatz and Flores, 1990 [ 2 ] |
| 1983 | Мышиные тимоциты | Yamagishi et al., 1983 [ 24 ] |
| 1983 | Мышиные лимфоциты | Tsuda et al., 1983 [ 26 ] |
Исследование 21 -го века
[ редактировать ]В 21 -м веке исследователи сосредоточились на лучшей характеристике специфических подтипов экцДНК, а также на структуре и функции этих молекул в биологических системах: [ 27 ]
- В 2012 году Shibata et al. обнаружил специфический тип эккДНК, называемую микродной . [ 6 ] Исследователи обнаружили десятки тысяч микродн в тканях мыши и клеточных линиях, а также клеточные линии человека. [ 6 ]
- В 2017 году Turner et al. Идентифицируется с использованием секвенирования всего генома (WGS), цитогенетического анализа и структурного моделирования, что экстрахромосомная круговая ДНК очень амплифицирована и распространена при различных типах рака . [ 28 ] Они обнаружили, что молекулы эккДНК обладают значительной гетерогенностью между различными клетками, даже если они получены из одного и того же человека. [ 28 ] Кроме того, эти молекулы эккДНК содержали гонщики с опухолью и, как сообщалось, редко обнаруживаются в нездоровых тканях. [ 28 ]
- В 2018 году Møller et al. Использовали здоровые образцы мышц человека и клетки крови для выявления более 100 000 типов экцДНК, что предполагало, что эккДНК может быть обнаружена в соматических клетках повсеместно. [ 29 ]
- В 2019 году Wu et al. обнаружил, что экдо (подтип эккДНК) ассоциируется с хроматином , но в отличие от хромосом, он не имеет уплотнения более высокого порядка, что увеличивает его доступность. [ 30 ]
- В 2021 году Wang et al. подробно описывается на формирование ECCDNAS и идентифицировала иммуностимуляторную функцию ECCDNAS. [ 31 ] Они также разработали улучшенный протокол очистки эккДНК, который уменьшает линейную загрязнение ДНК в очищенных образцах. [ 31 ]
Очистка эккДНК
[ редактировать ]Исторически, ECCDNA очищали с использованием двухэтапной процедуры, которая включала первую изолирующую неочищенную экстрахромосомную ДНК и впоследствии переваривает линейную ДНК посредством экзонуклеазы расщепления . [ 31 ] Тем не менее, этот метод часто приводит к линейному загрязнению ДНК, поскольку расщепление экзонуклеазы недостаточно для удаления всей линейной ДНК. [ 31 ] В 2021 году Wang et al. разработал трехступенчатый метод обогащения эккДНК, который улучшил очистку экцДНК: [ 31 ]
- Клетки сначала дегидратировали в> 90% метаноле. Для извлечения неочищенной экстрахромосомной ДНК клетки лизировали с pH 11,8 щелочным лизисным буфером, нейтрализовали с нейтрализационным буфером и осаждали с использованием буфера осадков. Коммерческий набор для очистки плазмиды была использована для выделения ДНК из других клеточных компонентов.
- Элюированную ДНК расщепляли с помощью рестрикционного фермента PACI для линеаризации митохондриальной ДНК ( мтДНК ) и экзонуклеазы, которая может переваривать линейную ДНК.
- Наконец, круглая ДНК избирательно извлекали коммерческим раствором и шариками кремнезема для удаления линейной ДНК, которая не была удалена экзонуклеазной расщеплением.
Двойные минуты (дм) против экстрахромосомной круглой ДНК (экцДНК)
[ редактировать ]Первоначально термин «двойные минуты » (DM) обычно использовался для обозначения экстрахромосомной круглой ДНК, потому что он часто появлялся как пара в ранних исследованиях. [ 27 ] Поскольку исследования продолжались, были идентифицированы различные подтипы экстрахромосомной круглой ДНК, которые не являются двойными минутами (например, микродна ). В 2014 году Barreto et al. определил, что двойные минуты составляют только примерно 30% экстрахромосомной ДНК. [ 32 ] Таким образом, термин экстрахромосомной круглой ДНК (экцДНК) становится все более широко используемым, в то время как термин двойной минуты теперь зарезервирован для определенного подтипа эккДНК. [ 32 ]
Структура
[ редактировать ]ЭккДНК представляют собой круглую ДНК, которые были обнаружены в клетках человека, растений и животных и присутствуют в клеточном ядре в дополнение к хромосомной ДНК . ЭккДНК отличается от другой круглой ДНК в клетках, таких как митохондриальная ДНК (мтДНК), потому что она колеблется в размерах от нескольких сотен оснований до мегабаз и получена из геномной ДНК. [ 1 ] Например, эккДНК может быть образована из экзонов кодирующих белков генов, таких как муцин и титин . Исследователи предположили, что эккДНК может способствовать экспрессии различных изоформ гена, мешая или способствуя транскрипции специфических экзонов . [ 1 ]
ЭккДНК была классифицирована как одна из четырех различных категорий круговой ДНК на основе размера и последовательности, включая небольшую полидисперсенную круглую ДНК (SPCDNA), теломерные круги (T-цирки), микродНК (100-400 п.н.) и экстрахромосомную ДНК (ECDNA), ECDNA), микродНК (100-400 п.н. и экстрахромосомная ДНК (ECDNA), ECDNA), микродНК (100-400 п.н. и экстрахромосомная ДНК (ECDNA), микродНК (100-400 п.н. Полем [ 27 ] Каждый из этих типов имеет свои уникальные биологические характеристики (см. Таблицу 2): [ 27 ]
| Название экцдны | Размер | Характеристики | Функция |
|---|---|---|---|
| spcdna | 100–10 КБ | Высоко разнообразный тип эккДНК, существует большой диапазон количества SpcDNA, обнаруженных клетки | Участвует в генетической нестабильности человека |
| Теломерные круги | кратные 738 п.н. | Образуются с помощью теломерных массивов, которые представляют собой серию повторяющихся последовательностей в конце линейной ДНК. | Участвует в альтернативном удлинении теломер (ALT) |
| Микродна | 100-400 п.н. | Получен из геномных мест, которые имеют высокое содержание GC и экзонов плотность | Экспрессируют небольшие функциональные регуляторные РНК (например, микроРНК и новые РНК, подобные IS). |
| экдна | 1-3 МБ | Включите полные гены, без теломер , пентрик | Усиление генов, участвующих в развитии рака и лекарственной устойчивости |
Биогенез эккДНК
[ редактировать ]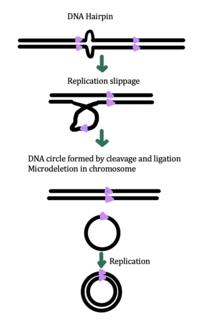


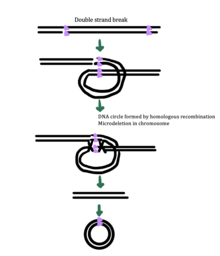
Хотя точный механизм генерации эккДНК до сих пор неизвестен, некоторые исследования показали, что генерация эккДНК может быть связана с восстановлением повреждения ДНК, [ 33 ] гипер-транскрипция, [ 33 ] [ 34 ] гомологичная рекомбинация , [ 35 ] и репликация стресса . [ 33 ] Существует несколько предложенных механизмов для образования экцДНК: (1) проскальзывание репликации создает петлю на цепи шаблона , которая затем иссекается и лигируется в круг, оставляя микроделеуцию на хромосоме , (2) проскальзывание репликации создает петлю в цепи продукта, что это целесообразно. Исправляется и лигируется в круг, который не генерирует , ( 3 механизм образования эккДНК oder ) микроделеуцию в хромосоме Круг и хромосома страдают микроделеоном [ 1 ]
Исследования, проведенные в 2021 году, показали, что апоптотические клетки являются источником ECCDNAS; Это было завершено из -за исследования, показывающего, что апоптотическая фрагментация ДНК (ADF) является предпосылкой для образования экцДНК с помощью методов очистки. [ 31 ]
ЭккДНК может быть получена в результате образования микро-нуклеев, что указывает на хромосомную нестабильность . Было предложено, что преждевременный апоптоз и/или ошибки в хромосомной сегрегации во время митоза могут привести к образованию микро-нуклеев. [ 36 ]
эккДНК в нездоровых клетках
[ редактировать ]Чтобы проверить, используются ли ECCDNA в нелоковых клетках, эмбриональные стволовые клетки мыши и анализ южно-блоттинга использовались ; Результаты подтвердили, что экцДНК обнаруживается как в раковых, так и в незвуковых клетках. [ 31 ] Также известно, что эккДНК вряд ли будет получена из специфических областей генома; Данные секвенирования от 2021 года сообщают, что данные предполагают, что ECCDNAS широко распространены на протяжении всего генома . [ 31 ] Картирование генома полноразмерных ECCDNAs продемонстрировало их различные паттерны выравнивания геном, которые включают в соседние, перекрывающиеся или вложенные положения на одной и той же хромосоме или в разных хромосомах . [ 31 ] ECCDNAs происходят в основном из одиночных, непрерывных геномных локусов, что означает, что один отдельный геномный фрагмент самостоятельно используется для формирования эккДНК, а не образуется из лигирования различных геномных фрагментов. [ 31 ] Эти два варианта могут быть классифицированы как непрерывные и непрерывные ECCDNA, соответственно. [ 31 ] причину циркулялизации фрагментированной ДНК, лигазы три различных фермента были протестированы млекопитающих: LIG1 , LIG3 и Чтобы еще больше понять LIG4 [ 31 ] Полем Используя нокаутные модели в клеточной линии мыши B12F3 B-лимфоцитов , исследования, проведенные в 2021 году, идентифицировали LIG3 в качестве основной лигазы для генерации экцдо в этих клетках. [ 31 ]
Функция
[ редактировать ]Точная функция ECCDNA была обсуждена, но некоторые исследования показали, что ECCDNA могут способствовать усилению генов при раке , [ 1 ] иммунная функция , [ 31 ] и старение . [ 34 ] [ 35 ] [ 37 ]
Функция экцДНК в иммунной системе
[ редактировать ]Согласно исследованию, проведенному в 2021 году, другой функцией ECCDNAS является их роль в качестве возможных иммуностимуляторов . [ 31 ] ЭккДНК значительно индуцирует интерфероны типа I (IFNα, IFNβ), интерлейкин-6 (IL-6) и фактор некроза опухоли (TNF), даже в большей степени, чем линейная ДНК и другие, как правило, мощные индукторы цитокинов при их высоких уровнях концентрации. [ 31 ] Подобные паттерны наблюдаются с макрофагами, поскольку данные показали, что ECCDNAs являются очень мощными иммуностимулянтами в активации как костном мозге, полученных в дендритных клеток, полученных из костного мозга так и макрофагов, . [ 31 ] Кроме того, эксперименты изменили структуру экцДНК с одним ник -ником на сегмент эккДНК и впоследствии обрабатывались ферментами для генерации линейных версий экцДНК. [ 31 ] В этих экспериментах было показано, цитокинов что транскрипция , важный маркер для активности иммунной системы , намного выше в необработанной экцДНК по сравнению с линеаризованной обработкой, придавая, что круговая структура эккДНК, а не сама генетическая последовательность, дает его эккДНК иммунная функция. [ 31 ]
Функция экцДНК при раке
[ редактировать ]Некоторые известные функции эккДНК включают вклад в межклеточную генетическую гетерогенность в опухолях , а также, в частности, онкогенов и усиление лекарственных генов . Это также подтверждает, что гены на экцДНК экспрессируются. В целом, эккДНК была связана с раком и лекарственной устойчивостью , старением , компенсацией генов, [ 1 ] И по этой причине это продолжает оставаться важной темой для обсуждения.
Приложения
[ редактировать ]Роль в раке
[ редактировать ]Подтип эккДНК, такой как экдо, рибосомной ДНК локус ( экстрахромосомный круг рДНК ), и двойные минуты были связаны с геномной нестабильностью . Двойные минуты ECDNAs представляют собой фрагменты экстрахромосомной ДНК , которые первоначально наблюдались в большом количестве опухолей человека , включая молочную грудь, легкие, яичники, толстую кишку и, в частности, нейробластому . Они являются проявлением амплификации генов во время развития опухолей, которые дают клеткам селективные преимущества роста и выживания. Двойные минуты, как и фактические , состоят из хроматина и повторяются в ядре клетки хромосомы во время деления клеток . В отличие от типичных хромосом, они состоят из круглых фрагментов ДНК , размером только несколько миллионов пар оснований и не содержат центромеры или теломер .
Было показано, что двойные хромосомы (DMS), которые присутствуют в виде парных хроматиновых тел при световой микроскопии , являются подмножеством экдо. [ 28 ] [ 38 ] Двойные минутные хромосомы составляют около 30% рак-содержащего спектра экдо, включая отдельные тела, [ 28 ] и было обнаружено, что он содержит идентичное содержание генов как отдельные тела. Обозначение экДНК охватывает все формы большой гены-содержащей экстрахромосомной ДНК, обнаруженной в раковых клетках . Этот тип экДНК обычно наблюдается в раковых клетках различных гистологий , но практически никогда не в нормальной ткани. [ 39 ] [ 28 ] Считается, что ECDNA продуцируется с помощью двухцепочечных разрывов в хромосомах или чрезмерной репликации ДНК в организме. [ 40 ]
Круглая форма экдо эксдо отличается от линейной структуры хромосомной ДНК значимыми способами, которые влияют на патогенез рака . [ 41 ] [ 30 ] Онкогены, кодируемые на экду, имеют огромный транскрипционный выход, ранжируя в 1% генов во всем транскриптоме . В отличие от бактериальных плазмид или митохондриальной ДНК , эСДНК хроматинизируются, содержащие высокие уровни активных марок гистонов , но недостаток репрессивных марок гистонов. В архитектуре экдна хроматина отсутствует уплотнение более высокого порядка, которое присутствует на хромосомной ДНК и является одной из наиболее доступной ДНК во всем геноме рака.
Из ECCDNA было обнаружено, что области прикрепления матрицы (MARS) активируют амплификацию онкогенов . [ 1 ] Трансфекция этих MARS в эмбриональные клетки почек 293T человека привела к увеличению экспрессии генов , что позволяет предположить, что эти MARS, полученные из ECCDNA, участвуют в активации онкогена. [ 42 ] ЭккДНК также, по-видимому, играет роль в других видах рака, таких как рак молочной железы , где онкогены в рецепторе эпидермального фактора роста человека (HER2), позитивные гены рака молочной железы в экцдо, амплифицируются. [ 1 ] Эта эккДНК также показала способность приобретать устойчивость к терапии рецепторных тирозинкиназ (RTK), как HER26. [ 43 ]
Роль в старении
[ редактировать ]Дрожжи являются модельными организмами для изучения старения , а ECCDNAs накапливаются в старых клетках и играют роль в выборе старения у дрожжей. [ 37 ] Продолжается спекуляция на общности этой концепции у более высоких видов, таких как млекопитающие . [ 37 ]
Смотрите также
[ редактировать ]- Экстрахромосомная ДНК
- Экстрахромосомный круг рДНК
- Двойная минута
- Микродна
- Эгоистичные генетические элементы
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час Полсен Т., Кумар П., Косеоглу М.М., Датта А (апрель 2018 г.). «Открытия экстрахромосомных кругов ДНК в нормальных и опухолевых клетках» . Тенденции в генетике . 34 (4): 270–278. doi : 10.1016/j.tig.2017.12.010 . PMC 5881399 . PMID 29329720 .
- ^ Jump up to: а беременный в Gaubatz JW (1990). «Экстрахромосомные круглые DNA и пластичность геномной последовательности в эукариотических клетках». Мутационные исследования . 237 (5–6): 271–292. doi : 10.1016/0921-8734 (90) 90009-G . PMID 2079966 .
- ^ Cohen S, Yacobi K, Segal D (июнь 2003 г.). «Экстрахромосомная круговая ДНК тандемно повторяющихся геномных последовательностей у дрозофилы» . Исследование генома . 13 (6A): 1133–1145. doi : 10.1101/gr.907603 . PMC 403641 . PMID 12799349 .
- ^ Jump up to: а беременный Cohen S, Agmon N, Sobol O, Segal D (март 2010 г.). «Экстрахромосомные круги спутниковых повторений и рибосомной ДНК 5S в клетках человека» . Мобильная ДНК . 1 (1): 11. doi : 10.1186/1759-8753-1-11 . PMC 3225859 . PMID 20226008 .
- ^ Стэнфилд С., Хелински Д.Р. (октябрь 1976 г.). «Маленькая круговая ДНК в Drosophila melanogaster». Клетка . 9 (2): 333–345. doi : 10.1016/0092-8674 (76) 90123-9 . PMID 824055 . S2CID 39382051 .
- ^ Jump up to: а беременный в Shibata Y, Kumar P, Layer R, Willcox S, Gagan Jr, Griffith JD, Dutta A (апрель 2012 г.). «Экстрахромосомные микродны и хромосомные микроделеи в нормальных тканях» . Наука . 336 (6077): 82–86. Bibcode : 2012sci ... 336 ... 82 с . doi : 10.1126/science.1213307 . PMC 3703515 . PMID 22403181 .
- ^ Møller HD, Parsons L, Jørgensen TS, Botstein D, Regenberg B (июнь 2015 г.). «Экстрахромосомная круговая ДНК распространена у дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (24): E3114 - E3122. BIBCODE : 2015PNAS..112E3114M . doi : 10.1073/pnas.1508825112 . PMC 4475933 . PMID 26038577 .
- ^ Jump up to: а беременный Shoura MJ, Gabdank I, Hansen L, Merker J, Gotlib J, Levene SD, Fire AZ (октябрь 2017 г.). «Сложные и специфичные для клеток популяции эндогенной круглой ДНК (экцДНК) у Caenorhabditis elegans и Homo Sapiens » . G3 . 7 (10): 3295–3303. doi : 10.1534/g3.117.300141 . PMC 5633380 . PMID 28801508 .
- ^ Hayday AC, Saito H, Gillies SD, Kranz DM, Tanigawa G, Eisen HN, Tonegawa S (февраль 1985 г.). «Структура, организация и соматическая перестройка гамма -генов Т -клеток». Клетка . 40 (2): 259–269. doi : 10.1016/0092-8674 (85) 90140-0 . PMID 3917858 . S2CID 34582929 .
- ^ Jump up to: а беременный в дюймовый Hotta Y, Bassel A (февраль 1965 г.). «Молекулярный размер и кругость ДНК в клетках млекопитающих и более высоких растений» . Труды Национальной академии наук Соединенных Штатов Америки . 53 (2): 356–362. Bibcode : 1965pnas ... 53..356H . doi : 10.1073/pnas.53.2.356 . PMC 219520 . PMID 14294069 .
- ^ Jump up to: а беременный Cox D, Yuncken C, Spriggs AI (июль 1965 г.). «Миловые хроматинские тела в злокачественных опухолях детства». Лансет . 1 (7402): 55–58. doi : 10.1016/s0140-6736 (65) 90131-5 . PMID 14304929 .
- ^ Billheimer FE, Avers CJ (октябрь 1969 г.). «Ядерная и митохондриальная ДНК из дикого типа и миниатюрных дрожжей: круглости, длина и плавучивая плотность» . Труды Национальной академии наук Соединенных Штатов Америки . 64 (2): 739–746. Bibcode : 1969pnas ... 64..739b . doi : 10.1073/pnas.64.2.739 . PMC 223406 . PMID 5261045 .
- ^ Beverley SM, Coderre JA, Santi DV, Schimke RT (сентябрь 1984 г.). «Нестабильные амплификацию ДНК у резистентных к метотрексату лейшмании состоят из экстрахромосомных кругов, которые ревоказуются во время стабилизации». Клетка . 38 (2): 431–439. doi : 10.1016/0092-8674 (84) 90498-7 . PMID 6467372 . S2CID 2030494 .
- ^ Nass MM, Ben-Shaul Y (июнь 1972 г.). «Новая закрытая круговая дуплексная ДНК в обесцвеченных мутантах и зеленых штаммах Euglena gracilis». Biochimica et Biophysica Acta (BBA) - нуклеиновые кислоты и синтез белка . 272 (1): 130–136. doi : 10.1016/0005-2787 (72) 90041-x . PMID 4625469 .
- ^ Вонг Фа, Wildman SG (январь 1972 г.). «Простая процедура выделения спутниковой ДНК из листьев табака с высоким уровнем урожая и демонстрации мини -округ». Biochimica et Biophysica Acta (BBA) - нуклеиновые кислоты и синтез белка . 259 (1): 5–12. doi : 10.1016/0005-2787 (72) 90468-6 . PMID 5011974 .
- ^ Агтериббе Э., Крун А.М., Ван Бругген Эф (май 1972). «Круглая ДНК из митохондрий нейроспоры Crassa» Biochimica et Biophysica Acta (BB) - нуклеиновые кислоты и протесты 269 (2): 299–3 Doi : 10.1016/0005-2787 (72) 90439- x 4260513PMID
- ^ Шталь У, Лемке П.А., Тудзинский П., Кюк У, Эссек К (июль 1978 г.). «Свидетельство о плазмиде, подобной ДНК, в нити, а ascomycete podospora anserina». Молекулярная и общая генетика . 162 (3): 341–343. doi : 10.1007/bf00268860 . PMID 683172 . S2CID 126554440 .
- ^ Lazarus CM, Earl AJ, Turner G, Küntzel H (май 1980). «Амплификация митохондриальной последовательности ДНК в цитоплазмически наследственном« рваном »мутанте aspergillus amstelodami» . Европейский журнал биохимии . 106 (2): 633–641. doi : 10.1111/j.1432-1033.1980.tb04611.x . PMID 6249580 .
- ^ Смит С.А., Виноград Дж (август 1972 г.). Полем Jogoral 69 (2): 163–178. doi : 10.1016/0022-2836 (72) 022-7-7 PMID 5070865 .
- ^ Buongiorno-Nardelli M, Amaldi F, Lava-Sanchez PA (март 1976 г.). «Анализ электронного микроскопа амплифицирующей рибосомной ДНК от Xenopus laevis». Экспериментальные исследования клеток . 98 (1): 95–103. doi : 10.1016/0014-4827 (76) 90467-5 . PMID 1253845 .
- ^ Delap RJ, Rush Mg (декабрь 1978 г.). «Изменение количества и распределения по размерам небольших круговых ДНК во время развития куриной бурсы» . Труды Национальной академии наук Соединенных Штатов Америки . 75 (12): 5855–5859. Bibcode : 1978pnas ... 75.5855d . doi : 10.1073/pnas.75.12.5855 . PMC 393074 . PMID 282606 .
- ^ Тода М., Хирама Т., Такешита С., Ямагиши Х (июнь 1989 г.). «Продукты эксцизионных перестроек гена иммуноглобулина». Иммунологические письма . 21 (4): 311–316. doi : 10.1016/0165-2478 (89) 90025-4 . PMID 2767726 .
- ^ Calabretta B, Robberson DL, Barrera-Saldaña HA, Lambrou TP, Saunders GF (март 1982 г.). «Нестабильность генома в области человеческой ДНК, обогащенной повторными последовательностями ALU». Природа . 296 (5854): 219–225. Bibcode : 1982natur.296..219c . doi : 10.1038/296219A0 . PMID 6278320 . S2CID 4265874 .
- ^ Jump up to: а беременный Yamagishi H, Kunisada T, Iwakura Y, Nishimune Y, Ogiso Y, Matsushiro A (декабрь 1983 г.). «Появление экстрахромосомных круговых ДНК-комплексов в качестве одного из самых ранних сигналов клеточной дифференцировки в раннем развитии эмбриона мыши (эмбрион мыши/тератокарцинома/слюна-пресс-адсорбция/круглое комплекс ДНК/перестройка ДНК)» . Развитие, рост и дифференциация . 25 (6): 563–569. doi : 10.1111/j.1440-169x.1983.00563.x . PMID 37282129 . S2CID 83666742 .
- ^ Tsuda T, Yamagishi H, Ohnishi N, Yamada Y, Izumi H, Mori KJ (November 1983). "Extrachromosomal circular DNAs from murine hemopoietic tissue cells". Plasmid. 10 (3): 235–241. doi:10.1016/0147-619X(83)90037-9. PMID 6657775.
- ^ Jump up to: a b Flores SC, Sunnerhagen P, Moore TK, Gaubatz JW (May 1988). "Characterization of repetitive sequence families in mouse heart small polydisperse circular DNAs: age-related studies". Nucleic Acids Research. 16 (9): 3889–3906. doi:10.1093/nar/16.9.3889. PMC 336563. PMID 3375074.
- ^ Jump up to: a b c d e Wang T, Zhang H, Zhou Y, Shi J (June 2021). "Extrachromosomal circular DNA: a new potential role in cancer progression". Journal of Translational Medicine. 19 (1): 257. doi:10.1186/s12967-021-02927-x. PMC 8194206. PMID 34112178.
- ^ Jump up to: a b c d e f Turner KM, Deshpande V, Beyter D, Koga T, Rusert J, Lee C, et al. (March 2017). "Extrachromosomal oncogene amplification drives tumour evolution and genetic heterogeneity". Nature. 543 (7643): 122–125. Bibcode:2017Natur.543..122T. doi:10.1038/nature21356. PMC 5334176. PMID 28178237.
- ^ Møller HD, Mohiyuddin M, Prada-Luengo I, Sailani MR, Halling JF, Plomgaard P, et al. (March 2018). "Circular DNA elements of chromosomal origin are common in healthy human somatic tissue". Nature Communications. 9 (1): 1069. Bibcode:2018NatCo...9.1069M. doi:10.1038/s41467-018-03369-8. PMC 5852086. PMID 29540679.
- ^ Jump up to: a b Wu S, Turner KM, Nguyen N, Raviram R, Erb M, Santini J, et al. (November 2019). "Circular ecDNA promotes accessible chromatin and high oncogene expression". Nature. 575 (7784): 699–703. Bibcode:2019Natur.575..699W. doi:10.1038/s41586-019-1763-5. PMC 7094777. PMID 31748743.
- ^ Jump up to: a b c d e f g h i j k l m n o p q r s Wang Y, Wang M, Djekidel MN, Chen H, Liu D, Alt FW, Zhang Y (November 2021). "eccDNAs are apoptotic products with high innate immunostimulatory activity". Nature. 599 (7884): 308–314. Bibcode:2021Natur.599..308W. doi:10.1038/s41586-021-04009-w. PMC 9295135. PMID 34671165. S2CID 239051756.
- ^ Jump up to: a b Barreto SC, Uppalapati M, Ray A (May 2014). "Small Circular DNAs in Human Pathology". The Malaysian Journal of Medical Sciences. 21 (3): 4–18. PMC 4163554. PMID 25246831.
- ^ Jump up to: a b c Dillon LW, Kumar P, Shibata Y, Wang YH, Willcox S, Griffith JD, et al. (June 2015). "Production of Extrachromosomal MicroDNAs Is Linked to Mismatch Repair Pathways and Transcriptional Activity". Cell Reports. 11 (11): 1749–1759. doi:10.1016/j.celrep.2015.05.020. PMC 4481157. PMID 26051933.
- ^ Jump up to: a b Hull RM, King M, Pizza G, Krueger F, Vergara X, Houseley J (December 2019). "Transcription-induced formation of extrachromosomal DNA during yeast ageing". PLOS Biology. 17 (12): e3000471. doi:10.1371/journal.pbio.3000471. PMC 6890164. PMID 31794573.
- ^ Jump up to: a b Gresham D, Usaite R, Germann SM, Lisby M, Botstein D, Regenberg B (October 2010). "Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus". Proceedings of the National Academy of Sciences of the United States of America. 107 (43): 18551–18556. Bibcode:2010PNAS..10718551G. doi:10.1073/pnas.1014023107. PMC 2972935. PMID 20937885.
- ^ Altungöz; Yüksel (September 2023). "Gene amplifications and extrachromosomal circular DNAs: function and biogenesis". Molecular Biology Reports. 50 (9): 7693–7703. doi:10.1007/s11033-023-08649-1. PMID 37433908 – via Springer.
- ^ Jump up to: a b c Sinclair DA, Guarente L (December 1997). "Extrachromosomal rDNA circles--a cause of aging in yeast". Cell. 91 (7): 1033–1042. doi:10.1016/s0092-8674(00)80493-6. PMID 9428525. S2CID 12735979.
- ^ deCarvalho AC, Kim H, Poisson LM, Winn ME, Mueller C, Cherba D, et al. (May 2018). "Discordant inheritance of chromosomal and extrachromosomal DNA elements contributes to dynamic disease evolution in glioblastoma". Nature Genetics. 50 (5): 708–717. doi:10.1038/s41588-018-0105-0. PMC 5934307. PMID 29686388.
- ^ Kim H, Nguyen NP, Turner K, Wu S, Gujar AD, Luebeck J, et al. (September 2020). "Extrachromosomal DNA is associated with oncogene amplification and poor outcome across multiple cancers". Nature Genetics. 52 (9): 891–897. doi:10.1038/s41588-020-0678-2. PMC 7484012. PMID 32807987.
- ^ Kuttler F, Mai S (February 2007). "Formation of non-random extrachromosomal elements during development, differentiation and oncogenesis". Seminars in Cancer Biology. 17 (1): 56–64. doi:10.1016/j.semcancer.2006.10.007. PMID 17116402.
- ^ Zimmer C (November 20, 2019). "Scientists Are Just Beginning to Understand Mysterious DNA Circles Common in Cancer Cells". New York Times.
- ^ Jin Y, Liu Z, Cao W, Ma X, Fan Y, Yu Y, et al. (2012). "Novel functional MAR elements of double minute chromosomes in human ovarian cells capable of enhancing gene expression". PLOS ONE. 7 (2): e30419. Bibcode:2012PLoSO...730419J. doi:10.1371/journal.pone.0030419. PMC 3272018. PMID 22319568.
- ^ Vicario R, Peg V, Morancho B, Zacarias-Fluck M, Zhang J, Martínez-Barriocanal Á, et al. (2015-06-15). "Patterns of HER2 Gene Amplification and Response to Anti-HER2 Therapies". PLOS ONE. 10 (6): e0129876. Bibcode:2015PLoSO..1029876V. doi:10.1371/journal.pone.0129876. PMC 4467984. PMID 26075403.
Further reading
[edit]- Sinclair DA, Guarente L (December 1997). "Extrachromosomal rDNA circles--a cause of aging in yeast". Cell. 91 (7): 1033–1042. doi:10.1016/s0092-8674(00)80493-6. PMID 9428525.
- Cohen S, Houben A, Segal D (March 2008). "Extrachromosomal circular DNA derived from tandemly repeated genomic sequences in plants". The Plant Journal. 53 (6): 1027–1034. doi:10.1111/j.1365-313x.2007.03394.x. PMID 18088310.
- Cohen S, Regev A, Lavi S (February 1997). "Small polydispersed circular DNA (spcDNA) in human cells: association with genomic instability". Oncogene. 14 (8): 977–985. doi:10.1038/sj.onc.1200917. PMID 9050997.
- Kumar P, Dillon LW, Shibata Y, Jazaeri AA, Jones DR, Dutta A (September 2017). "Normal and Cancerous Tissues Release Extrachromosomal Circular DNA (eccDNA) into the Circulation". Molecular Cancer Research. 15 (9): 1197–1205. doi:10.1158/1541-7786.MCR-17-0095. PMC 5581709. PMID 28550083.
- Baskin F, Rosenberg RN, Dev V (June 1981). "Correlation of double-minute chromosomes with unstable multidrug cross-resistance in uptake mutants of neuroblastoma cells". Proceedings of the National Academy of Sciences of the United States of America. 78 (6): 3654–3658. Bibcode:1981PNAS...78.3654B. doi:10.1073/pnas.78.6.3654. PMC 319629. PMID 6943568. Free full-text.
- Barker PE (February 1982). "Double minutes in human tumor cells". Cancer Genetics and Cytogenetics. 5 (1): 81–94. doi:10.1016/0165-4608(82)90043-7. PMID 6175392.
- Masters J, Keeley B, Gay H, Attardi G (May 1982). "Variable content of double minute chromosomes is not correlated with degree of phenotype instability in methotrexate-resistant human cell lines". Molecular and Cellular Biology. 2 (5): 498–507. doi:10.1128/MCB.2.5.498. PMC 369819. PMID 7110138. Free full-text.
- Yüksel A, Altungöz O (September 2023). "Gene amplifications and extrachromosomal circular DNAs: function and biogenesis". Mol Biology Reports. 50 (9): 7693–7703. doi:10.1007/s11033-023-08649-1. PMID 37433908.