Адено-ассоциированный вирус
| Адено-ассоциированный вирус | |
|---|---|
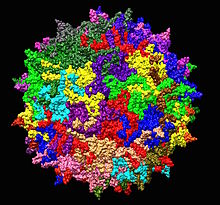
| |
| Адено-ассоциированный вирусный серотип 2 структура от 1LP3. Одна пятикратная ось показана в центре. | |
| Научная классификация | |
| (не вмешательство): | Вирус |
| Область : | Моноднавирия |
| Королевство: | Shotokuvirae |
| Филум: | Cossaviricota |
| Сорт: | Пятый |
| Заказ: | Пиковиралс |
| Семья: | Parvoviridae |
| Подсемейство: | Parvovirinae |
| Род: | Зависимость |
| Вирусы включены: | |
| |
Адено-ассоциированные вирусы ( AAV ) представляют собой небольшие вирусы, которые заражают людей и некоторые другие виды приматов . Они принадлежат к роду Devestoparvovirus , который, в свою очередь, принадлежит семейству Parvoviridae . Они представляют собой небольшие (приблизительно 26 нм в диаметре), дефектные репликации , неразвитые вирусы и имеют линейный одноцепочечный геном ДНК (SSDNA) приблизительно 4,8 килобаз (KB). [ 1 ] [ 2 ]
Несколько особенностей делают AAV привлекательным кандидатом на создание вирусных векторов для генной терапии и для создания изогенных моделей заболеваний человека . [ 3 ] генной терапии Векторы , использующие AAV, могут заразить как делящихся, так и покоящихся клеток и сохраняться в экстрахромосомном состоянии без интеграции в геном клетки -хозяина. Однако в нативном вирусе возникает интеграция генов, переносимых вирусом в геном хозяина. [ 4 ] Интеграция может быть важной для определенных приложений, но также может иметь нежелательные последствия. Недавние клинические испытания человека с использованием AAV для генной терапии в сетчатке показали перспективу. [ 5 ]
В марте 2023 года серия документов природы обнаружила высокие титры адено-ассоциированного вируса 2 (AAV2), наряду с аденовирусом и герпесвирусом, в образцах от волны детства гепатита. [ 6 ] В одной статье предполагается, что коинфекция AAV2 может способствовать более серьезным заболеваниям печени, чем инфекция только с адено- или герпесвирусами, и что причинно-следственная связь еще предстоит установить. [ 7 ]
История
[ редактировать ]Аденоассоцированный вирус (AAV), ранее считался загрязняющим веществом при препаратах аденовируса, был впервые идентифицирован как зависимость в 1960-х годах в лабораториях Боба Атчисона в Питтсбурге и Уоллес-Роу в NIH . Серологические исследования у людей впоследствии показали, что, несмотря на то, что присутствуют у людей, инфицированных вспомогательными вирусами , такими как аденовирус или вирус герпеса, сам AAV не вызывал каких -либо заболеваний. [ 8 ]
Использование в генной терапии
[ редактировать ]Преимущества и недостатки
[ редактировать ]AAV дикого типа вызвал значительный интерес исследователей генной терапии из-за ряда функций. Главным среди них было явное отсутствие патогенности вируса. Он также может заразить невидающиеся клетки и способен стабильно интегрироваться в геном клеток-хозяина на определенном месте (обозначенный AAVS1) в хромосоме человека 19 человека . [ 9 ] [ 10 ] Эта особенность делает его несколько более предсказуемым, чем ретровирусы , которые представляют угрозу случайной вставки и мутагенеза, что иногда сопровождается развитием рака. Геном AAV чаще всего интегрируется в упомянутый сайт, в то время как случайные включения в геном происходят с незначительной частотой. Разработка AAV в качестве векторов генной терапии, однако, устранила эту интегративную способность путем удаления повторения и CAP из ДНК вектора . Желаемый ген вместе с промотором для управления транскрипцией гена вставлен между инвертированными терминальными повторами (ITRS), которые помогают в образовании соглашения в ядре после одноцепочечной векторной ДНК конвертируется ДНК-полимеразы клеток-клетки в двухцепочечные ДНК Векторы генной терапии на основе AAV образуют эпизомальные соглашения в ядре клеток-хозяина. В не добывающихся клетках эти соглаживания остаются нетронутыми для жизни клетки-хозяина. В делящихся клетках ДНК AAV теряется через деление клеток, поскольку эпизомальная ДНК не реплицируется вместе с ДНК клеток -хозяина. [ 11 ] Случайная интеграция ДНК AAV в геном хозяина обнаруживается, но происходит на очень низкой частоте. [ 11 ] AAV также представляют очень низкую иммуногенность , казалось бы, ограниченной генерацией нейтрализующих антител , в то время как они не вызывают четко определенного цитотоксического ответа . [ 12 ] [ 13 ] [ 14 ] Эта особенность, наряду со способностью заражать покоящиеся клетки, представляют свое доминирование над аденовирусами в качестве векторов для генной терапии человека . [ Цитация необходима ]
Использование вируса действительно представляет некоторые недостатки. Клонирующая способность вектора относительно ограничена, и большинство терапевтических генов требуют полной замены генома вируса 4,8 килобазы. Следовательно, большие гены не подходят для использования в стандартном векторе AAV. В настоящее время исследуются варианты, чтобы преодолеть ограниченную мощность кодирования. [ 15 ] AAV ITR из двух геномов могут отжигать, чтобы сформировать конкатемеры с головой к хвостам, почти удваивая способность вектора. Вставка сайтов сплайсинга позволяет удалять ITRS из стенограммы. [ Цитация необходима ]
Из-за специализированных преимуществ генной терапии AAV исследователи создали измененную версию AAV, называемую самодополнительным адено-ассоциированным вирусом (SCAAV) . Принимая во внимание, что пакеты AAV одну цепь ДНК и должны ждать синтезированной его второй цепи, SCAAV упаковывают две более короткие нити, которые дополняют друг друга. Избегая синтеза второго стойки, SCAAAV может выразить быстрее, хотя в качестве предостережения SCAAV может кодировать только половину и без того ограниченной мощности AAV. [ 16 ] Недавние сообщения показывают, что векторы SCAAV являются более иммуногенными, чем одноцепочечные аденовирусные векторы, вызывая более сильную активацию цитотоксических Т -лимфоцитов . [ 17 ]
Гуморальный иммунитет, спровоцированный инфекцией диким типом, считается обычным. Связанная нейтрализующая активность ограничивает полезность наиболее часто используемого серотипа AAV2 в определенных приложениях. Соответственно, большинство клинических испытаний, проведенных, включают в себя доставку AAV2 в мозг, относительно иммунологически привилегированный орган. В мозге AAV2 сильно зависит от нейрон. [ Цитация необходима ]
Клинические испытания
[ редактировать ]По состоянию на 2019 год векторы AAV использовались в более чем 250 клинических испытаниях по всему миру, примерно 8,3% вирусных исследований генной терапии. [ 18 ] В последнее время были получены многообещающие результаты в исследованиях фазы 1 и фазы 2 для ряда заболеваний, включая врожденный амауроз Лебера , [ 5 ] [ 19 ] [ 20 ] гемофилия , [ 21 ] застойная сердечная недостаточность , [ 22 ] позвоночник мышечной атрофии , [ 23 ] дефицит липопротеиновой липазы , [ 24 ] и болезнь Паркинсона . [ 25 ]
| Индикация | Ген | Маршрут администрирования | Фаза | Номер предмета | Статус |
| Муковисцидоз | CFTR | Легкое, через аэрозоль | я | 12 | Полный |
| CFTR | Легкое, через аэрозоль | II | 38 | Полный | |
| CFTR | Легкое, через аэрозоль | II | 100 | Полный | |
| Гемофилия б | ИСПРАВИТЬ | Внутримышечный | я | 9 | Полный |
| ИСПРАВИТЬ | Печеночная артерия | я | 6 | Закончился | |
| Артрит | TNFR: FC | Внутриистористого | я | 1 | Непрерывный |
| Наследственная эмфизема | Аат | Внутримышечный | я | 12 | Непрерывный |
| Врожденный амауроз Лебера | RPE65 | Субретина | Я -ii | Несколько | Несколько продолжающихся и полных ( Voretigene neparvovec ) |
| Возрастная дегенерация макула | SFLT-1 | Субретина | Я -ii | 24 | Непрерывный |
| Мышечная дистрофия Дюшенна | SGCA | Внутримышечный | я | 10 | Непрерывный |
| Болезнь Паркинсона | GAD65 , GAD67 | Внутричерепный | я | 12 | Полный [ 27 ] |
| Болезнь Канавана | Аак | Внутричерепный | я | 21 | Непрерывный |
| Болезнь Баттена | CLN2 | Внутричерепный | я | 10 | Непрерывный |
| Болезнь Альцгеймера | NGF | Внутричерепный | я | 6 | Непрерывный |
| Спинальная мышечная атрофия | Smn1 | Внутривенное, интратекальное | I-ii | 150+ | Несколько продолжающихся и полных ( онсаногеновая абепарвовек ) |
| Застойная сердечная недостаточность | Serca 2a | Внутрикоронарный | IIB | 250 | Непрерывный |
Фундаментальная биология
[ редактировать ]
Геномика, транскриптомика и протеомика
[ редактировать ]Геном AAV построен из одноцепочечной дезоксирибонуклеиновой кислоты (SS DNA ), либо положительной, или отрицательной, длиной около 4,7 килобазы. Геном состоит из ITRS на обоих концах цепи ДНК и две открытые рамки считывания (ORF): Rep and Cap . Первый состоит из четырех перекрывающихся генов, кодирующих REP -белки, необходимые для жизненного цикла AAV, а последний содержит перекрывающиеся нуклеотидные последовательности капсидных белков: VP1, VP2 и VP3, которые взаимодействуют с образованием капсида с икозаэдральной симметрией. [ 28 ]
ITR -последовательности
[ редактировать ]Последовательности перевернутых терминальных повторений (ITR) составляют 145 основы каждая. Они были названы так из -за их симметрии, которая, как было показано, требовалось для эффективного умножения генома AAV. [ 29 ] Особенностью этих последовательностей, которая дает им это свойство, является их способность образовывать шпильку , которая способствует так называемому самоповреждению, которое позволяет primase -независимый синтез второй прядь ДНК. Также было показано, что ITRS потребуется как для интеграции ДНК AAV в геном клеток -хозяина (19 -я хромосома у людей), так и спасение от нее, [ 30 ] [ 31 ] а также для эффективного инкапстакса ДНК AAV в сочетании с генерацией полностью собранных, дезоксирибонуклеазируемых частиц AAV. [ 32 ]
Что касается генной терапии, ITR, по -видимому, являются единственными последовательностями, необходимыми в CIS рядом с терапевтическим геном: структурные ( CAP ) и упаковочные белки ( Rep ) могут быть доставлены в транс . С этим предположением было установлено много методов для эффективной продукции рекомбинантных векторов AAV (RAAV), содержащих репортер или терапевтический ген. Тем не менее, было также опубликовано, что ITR не являются единственными элементами, необходимыми в CIS для эффективной репликации и инкапсадации. Несколько исследовательских групп идентифицировали последовательность, обозначенную CIS-действие REP-зависимый элемент (CARE), внутри кодирующей последовательности гена Rep . Было показано, что забота увеличивает репликацию и инкапдиацию, когда присутствует в CIS . [ 33 ] [ 34 ] [ 35 ] [ 36 ]
rep gene и rep белки
[ редактировать ]На «левой стороне» генома есть два промотора, называемых p5 и p19, из которых могут быть получены две перекрывающиеся градоруклеиновые кислоты ( мРНК ) различной длины. Каждый из них содержит интрон , который может быть либо сплайсирован , либо нет. Учитывая эти возможности, могут быть синтезированы эти возможности, четыре различных мРНК и, следовательно, четыре различных белка Rep с перекрывающейся последовательности. Их имена изображают их размеры в килодотоне (KDA): Rep78, Rep68, Rep52 и Rep40. [ 37 ] Rep78 и 68 могут специфически связывать шпильку , образованную ITR в Законе о самопознании и расщепляются в определенной области, обозначенном месте разрешения терминала, внутри шпильки. Также было показано, что они необходимы для AAVS-специфической интеграции генома AAV. Было показано, что все четыре белка повторения связывают АТФ и обладают активностью геликазы . Также было показано, что они активируют транскрипцию от промотора P40 (упомянутое ниже), но подавляют промоторов как P5, так и P19. [ 31 ] [ 37 ] [ 38 ] [ 39 ] [ 40 ] [ 41 ]
CAP Gene и VP -белки
[ редактировать ]Правая сторона генома AAV с положительностью кодирует перекрывающиеся последовательности трех капсидных белков, VP1, VP2 и VP3, и двух вспомогательных белков, MAAP & AAP, которые начинаются с одного промотора, обозначенного p40. Молекулярные массы этих белков составляют 87, 72 и 62 килодонда соответственно. [ 42 ] AAV -капсид состоит из смесью VP1, VP2 и VP3, общей 60 мономеров, расположенных в икосаэдрической симметрии в соотношении 1: 1: 10, [ 43 ] с пустой массой приблизительно 3,8 МДа . [ 44 ] Кристаллическую структуру белка VP3 определяли Xie, Bue, et al. [ 45 ]

Ген CAP продуцирует дополнительный неструктурный белок, называемый активирующим сборку белка (AAP). Этот белок продуцируется из ORF2 и имеет важное значение для процесса капсид-сборки. [ 46 ] Точная функция этого белка в процессе сборки и его структуры не была решена до настоящего времени. [ Цитация необходима ]
Все три VP переводятся из одной мРНК. После того, как эта мРНК синтезируется, ее можно сплачивать в двух разных манерах: либо более длинный, либо более короткий интрон может быть вырезан, что приведет к образованию двух пулов мРНК: 2,3 кб и пула мРНК длиной 2,6 кб. Обычно, особенно при наличии аденовируса, более длинный интрон является предпочтительным, поэтому мРНК длиной 2,3 кб представляет собой так называемый «основной сплайс». В этой форме первый кодон AUG , из которого запускается синтез белка VP1, вырезается, что приводит к снижению общего уровня синтеза белка VP1. Первый кодон AUG, который остается в основном сплайке, является кодоном инициации для белка VP3. Однако выше этого кодона в той же открытой рамке считывания лежит последовательность ACG (кодирование треонина), которая окружена оптимальным контекстом Козака . Это способствует низкому уровню синтеза белка VP2, который фактически представляет собой белок VP3 с дополнительными N -конечными остатками, как и VP1. [ 47 ] [ 48 ] [ 49 ] [ 50 ]
Поскольку предпочтительным предпочтительнее, что более крупный интрон был сплайсирован, и, поскольку в основном сплайке кодон ACG является гораздо более слабым сигналом инициации трансляции , отношение, при котором структурные белки AAV синтезируются in vivo, составляет около 1: 1: 20, которое является так же, как в зрелой частице вируса. [ 51 ] Было показано, что уникальный фрагмент на N -конце белка VP1 обладает активностью фосфолипазы A2 (PLA2), что, вероятно, требуется для выпуска частиц AAV из поздних эндосом . [ 52 ] Muralidhar et al. сообщили, что VP2 и VP3 имеют решающее значение для правильной сборки вириона. [ 49 ] Совсем недавно, Warrington et al. показал, что VP2 не нужен для полного образования вирусных частиц и эффективной инфекционности, а также представил, что VP2 может переносить большие вставки в его N -конце, в то время как VP1 не может, вероятно, из -за присутствия домена PLA2. [ 53 ]
Посттрансляционные модификации
[ редактировать ]Недавние открытия, сделанные с использованием высокопроизводительных подходов к омике, включают тот факт, что капсиды AAV посттрансляционно модифицируются (PTM) во время производства, такие как ацетилирование, метилирование, фосфорилирование, деамидирование, O-гликнацилирование [ 54 ] и сумоилирование по всему капсидным белкам VP1, VP2 и VP3. Эти PTM различаются в зависимости от производственной производственной платформы. Другим таким открытием является тот факт, что геномы AAV эпигенетически метилированы во время продукции. Помимо цены, эти результаты могут повлиять на кинетику экспрессии, связывание рецептора RAAV, перенос, векторную иммуногенность и долговечность экспрессии. [ 55 ] [ 56 ]
Классификация, серотипы, рецепторы и нативный тропизм
[ редактировать ]| Серотип | Ткань тропизм [ 57 ] | Пройти барьер кровавого мозга | Клеточный тропизм | Синтетический | Комментарий | Опубликованный Cap / Rep |
|---|---|---|---|---|---|---|
| 2 | Гладкая мышца, ЦНС, печень | Нет | Нет | |||
| 5 | ЦНС, гладкая мышца | Нет | Нет | |||
| 8 | ЦНС, мозг, печень, гладкая мышца | Нет | Нет | |||
| 9 | ЦНС, печень, гладкая мышца | Да | Нет | |||
| PAL2 | CNS | Да | Да | Экспрессия печени составляет 1/4 AAV9 | Нет | |
| 9p1 / aavmyo | CNS | Да | астроциты | Да | Высокая мускулатура, включая скелетные мышцы, сердце и диафрагсклетальная трансдукция | Нет |
Два вида AAV были признаны Международным комитетом по таксономии вирусов в 2013 году: адено-ассоциированного зависимости зависимости зависимости A (ранее AAV-1, -2, -3 и -4) и адено, ассоциированного с зависимым от зависимых от AAV-5). [ 58 ]
До 1990 -х годов практически вся биология AAV была изучена с использованием серотипа AAV 2. Серотипы 2, 3, 5 и 6 были обнаружены в клетках человека, серотипах AAV 1, 4 и 7–11 в нечеловеческих образцах приматов. [ 59 ] По состоянию на 2006 год было описано 11 серотипов AAV , 11 -е место в 2004 году. [ 60 ] Капсидные белки AAV содержат 12 областей поверхностных областей с гипервариацией, причем большая часть изменчивости происходит в трехкратных проксимальных пиках, но геном парвовируса в целом представляет высоко консервативную репликацию и структурные гены в серотипах. [ 59 ] Все известные серотипы могут заражать клетки из нескольких разнообразных типов тканей. Специфичность ткани определяется капсидным серотипом, и псевдотипирование векторов AAV для изменения их диапазона тропизма, вероятно, будет важно для их использования в терапии.
Серотип 2
[ редактировать ]Серотип 2 (AAV2) был наиболее тщательно изучен до сих пор. [ 61 ] [ 62 ] [ 63 ] [ 64 ] [ 65 ] [ 66 ] AAV2 представляет естественный тропизм в направлении скелетных мышц , [ 67 ] нейроны , [ 61 ] сосудистые клетки гладких мышц [ 68 ] и гепатоциты . [ 69 ]
Три клеточных рецептора были описаны для AAV2: гепаран-сульфатный протеогликан (HSPG), V β 5 Интегрин и фактора роста фибробластов рецептор (FGFR-1). Первые функционируют как первичный рецептор, в то время как последние два обладают активностью ко-рецептора и позволяют AAV входить в клетку путем рецепторного эндоцитоза . [ 70 ] [ 71 ] [ 72 ] Эти результаты исследования были оспорены QIU, Handa, et al. [ 73 ] HSPG функционирует как первичный рецептор, хотя его изобилие во внеклеточном матриксе может привлекать частицы AAV и ухудшить эффективность инфекции. [ 74 ]
Исследования показали, что серотип 2 вируса (AAV-2), по-видимому, убивает раковые клетки, не причиняя вреда здоровым. «Наши результаты показывают, что адено-ассоциированный вирус типа 2, который заражает большую часть популяции, но не имеет известных вредных воздействий, убивает несколько видов раковых клеток, но не влияет на здоровые клетки»,-сказал Крейг Мейерс, [ 75 ] Профессор иммунологии и микробиологии в штата Пенсильвания Медицинском колледже в Пенсильвании в 2005 году. [ 76 ] Это может привести к новому противораковому агенту.
В марте 2023 года серия документов природы связала инфекцию адено-ассоциированного вируса 2 (AAV2) с волной гепатита детства. [ 6 ]
Другие серотипы
[ редактировать ]Этот раздел должен быть обновлен . ( Декабрь 2023 г. ) |
Хотя AAV2 является наиболее популярным серотипом в различных исследованиях на основе AAV, было показано, что другие серотипы могут быть более эффективными в качестве векторов доставки генов. AAV9 проходит кроваво-мозговой барьер у людей, AAV6 выглядит гораздо лучше при заражении эпителиальных клеток дыхательных путей, [ 77 ] [ 78 ] AAV7 представляет очень высокую скорость трансдукции клеток мышиных скелетных мышц (аналогично AAV1 и AAV5), AAV8 трансдуцируют гепатоциты. [ 79 ] [ 80 ] [ 81 ] и было показано, что AAV1 и 5 очень эффективны при доставке генов в эндотелиальные клетки сосудов. [ 82 ] В мозге большинство серотипов AAV показывают нейрональный тропизм, в то время как AAV5 также трансдуцирует астроциты. [ 83 ] AAV6, гибрид AAV1 и AAV2, [ 81 ] Также показывает более низкую иммуногенность, чем AAV2. [ 80 ]
Серотипы могут отличаться с уважением к рецепторам, с которыми они связаны. Например, трансдукция AAV4 и AAV5 может быть ингибирована растворимыми сиаловыми кислотами (различной формы для каждого из этих серотипов), [ 84 ] и было показано, что AAV5 входит в клетки через рецептор фактора роста, полученный из тромбоцитов . [ 85 ]
Синтетические серотипы
[ редактировать ]Было предпринято много усилий по разработке и улучшению новых вариантов AAV как для клинических, так и для исследовательских целей. Такие модификации включают новые тропизмы для нацеливания на определенные ткани и модифицированные поверхностные остатки, чтобы избежать обнаружения иммунной системой. Помимо выбора конкретных штаммов рекомбинантного AAV (RAAV) для нацеливания на конкретные клетки, исследователи также изучили псевдотипирование AAV, практику создания гибридов определенных штаммов AAV, чтобы приблизиться к еще более утонченной мишени. Гибрид создается путем взятия капсида из одного штамма и генома из другого штамма. Например, исследования с участием AAV2/5, гибридом с геномом AAV2 и капсидом AAV5, смогли достичь большей точности и диапазона в клетках мозга, чем AAV2, смогут достичь негибридированного. Исследователи продолжали экспериментировать с псевдотипированием, создавая штаммы с гибридными капсидами. AAV-DJ имеет гибридный капсид из восьми различных штаммов AAV; Таким образом, он может заразить различные клетки во многих областях тела, свойство, которого не будет иметь единственного штамма AAV с ограниченным тропизмом. [ 86 ] Другие усилия по разработке и улучшению новых вариантов AAV включали в себя наследственную реконструкцию вариантов вируса для создания новых векторов с улучшенными свойствами для клинических применений и изучением биологии AAV. [ 87 ]
Иммунология
[ редактировать ]AAV представляет особый интерес для генных терапевтов из-за ее очевидной ограниченной способности индуцировать иммунные реакции у людей, что должно положительно влиять на эффективность трансдукции векторной трансдукции при одновременном снижении риска любой иммунной патологии .
AAV не считается какой -либо известной ролью в болезнях. [ 88 ] Тем не менее, реакция иммунной системы хозяина и иммунная толерантность снижают эффективность AAV-опосредованной генной терапии. Было показано, что иммунный ответ хозяина реагирует на векторы AAV, трансдуцированные клетки и трансдуцированные белки. [ 89 ] Иммунный ответ может быть подразделен на две категории: врожденные и адаптивные, последний из которых разделен на гуморальный и клеточный опосредованный. [ 90 ] [ 91 ]
Врожденный
[ редактировать ]Врожденный . иммунный ответ на векторы AAV был охарактеризован на животных моделях Внутривенное введение у мышей вызывает переходную продукцию провоспалительных цитокинов и некоторую инфильтрацию нейтрофилов и других лейкоцитов в печень, что, по-видимому, изобилует большой процент инъецированных вирусных частиц. Как растворимые уровни, так и инфильтрация клеток, по -видимому, возвращаются к исходному уровню в течение шести часов. Напротив, более агрессивные вирусы производят врожденные реакции, длится 24 часа или дольше. [ 92 ]
Исследования in vivo показывают, что векторы AAV взаимодействуют с путями Toll-подобного рецептора (TLR) 9- и TLR2-MYD88, чтобы вызвать врожденный иммунный ответ, стимулируя продукцию интерферонов. [ 93 ] Показано, что мыши с дефицитом TLR9 более восприимчивы к лечению AAV и демонстрируют более высокие уровни экспрессии трансгена [ 94 ]
Гуморальный
[ редактировать ]Из -за предыдущей естественной инфекции многие люди имеют ранее существовавшие нейтрализующие антитела (NAB) против AAV, что может значительно препятствовать его применению в генной терапии. [ 95 ] Несмотря на то, что AAV очень варьируются среди вариантов дикого типа и синтетических вариантов, сайты распознавания антител могут быть сохранены эволюционно. [ 96 ]
Известно, что вирус способствует надежному гуморальному иммунитету на животных моделях и в человеческой популяции, где, как считается, до 80% людей являются серопозитивными для AAV2. антитела Известно, что нейтрализуют, а для применений генной терапии они влияют на эффективность трансдукции векторной вектора через некоторые маршруты введения. Помимо постоянных уровней антител, специфичных для AAV, он появляется как из первичных исследований у животных, так и из клинических испытаний, что B-клеточная память также является сильной. [ 97 ] У серопозитивных людей циркулирующие антитела IgG для AAV2, по -видимому, в основном состоят из подклассов IgG1 и IgG2, с незначительным или отсутствующим IgG3 или IgG4. [ 98 ]
Клеточная опосредованная
[ редактировать ]Клеточный ответ на вирус и векторы плохо охарактеризован и в значительной степени игнорируется в литературе еще в 2005 году. [ 97 ] Клинические испытания с использованием вектора на основе AAV2 для лечения гемофилии B, по-видимому, указывают на то, что может происходить целенаправленное разрушение трансдуцированных клеток. [ 99 ] В сочетании с данными, которые показывают, что CD8+ Т-клетки могут распознавать элементы капсида AAV in vitro , [ 100 ] Похоже, что может быть реакция цитотоксических Т -лимфоцитов на векторы AAV. Цитотоксические реакции будут подразумевать участие CD4+ T Helper -клеток в ответ на данные AAV, а in vitro из исследований на людях предполагают, что вирус действительно может вызывать такие ответы, включая ответы как TH1, так и TH2 памяти. [ 98 ] Ряд кандидатов, стимулирующих Т -клеточные эпитопы, был идентифицирован в капсидном белке AAV VP1, который может быть привлекательным мишенями для модификации капсида, если вирус должен использоваться в качестве вектора для генной терапии. [ 98 ] [ 99 ]
Инфекционный цикл
[ редактировать ]В цикле инфекции AAV есть несколько этапов, от заражения клетки до получения новых инфекционных частиц: [ Цитация необходима ]
- Прикрепление к клеточной мембране
- Рецептор-опосредованный эндоцитоз
- Эндосомальная торговля
- сбежать из поздней эндосомы или лизосомы
- транслокация в ядро
- не зарегистрируясь
- образование двухцепочечной репликативной формы генома AAV
- Экспрессия повторения генов
- генома Репликация
- Экспрессия генов CAP , синтез частиц потомства ssDNA
- сборка полных вирионов и
- высвобождение из зараженной клетки.
Некоторые из этих шагов могут выглядеть по -разному в различных типах клеток, которые частично способствуют определенному и довольно ограниченному нативному тропизму AAV. Репликация вируса также может варьироваться в одном типе ячейки, в зависимости от фазы текущего клеточного цикла клетки . [ 101 ]
Характерной особенностью адено-ассоциированного вируса является дефицит репликации и, следовательно, его неспособность умножать в незатронутых клетках. Адено-ассоциированный вирус распространяется за счет совместного инфицирования клетки с помощью помощника. Первым вспомогательным вирусом, который был описан как обеспечение успешного генерации новых частиц AAV, был аденовирусом, из которого возникло имя AAV. Тогда было показано, что репликации AAV может быть облегчена выбранными белками, полученными из аденовирусного генома,. [ 102 ] [ 103 ] другими вирусами, такими как HSV [ 104 ] или вакциния, или генотоксические агенты, такие как ультрафиолетовое излучение или гидроксимочевина . [ 105 ] [ 106 ] [ 107 ] В зависимости от наличия или отсутствия вируса помощника жизненный цикл AAV следует либо литическим, либо лизогенным путем, соответственно. [ 108 ] Если есть вирус помощника, экспрессия гена AAV активируется, позволяя вирусу воспроизвести с помощью полимеразы клетки хозяина. Когда вирус помощника убивает клетку -хозяина, новые вирионы AAV выпускаются. Если нет вируса помощника, AAV демонстрирует лизогенное поведение. Когда AAV заражает только клетку, его экспрессия гена репрессируется (AAV не повторяется), а ее геном включается в геном хозяина (в хромосому человека 19). В редких случаях лизис может происходить без вируса помощника, но обычно AAV не может воспроизводить и убивать клетку самостоятельно. [ 109 ]
Минимальный набор аденовирусных генов, необходимых для эффективной генерации, частиц потомства AAV, был обнаружен Matsushita, Ellinger et al. [ 102 ] Это открытие позволило создать новые методы производства рекомбинантного AAV, которые не требуют аденовирусного коинфекции клеток, продуцирующих AAV. В отсутствие вспомогательных вирусов или генотоксических факторов ДНК AAV может либо интегрироваться в геном -хозяина, либо сохраняться в эпизомальной форме. В первом случае интеграция опосредована белками Rep78 и Rep68 и требует наличия ITRS, фланкирующих регион, интегрированную. У мышей геном AAV наблюдался, сохраняющийся в течение длительных периодов времени в покоящихся тканях, таких как скелетные мышцы, в эпизомальной форме (круговая конформация головы к хвосту). [ 110 ]
Смотрите также
[ редактировать ]- Изогенные модели заболеваний человека
- Онколитический аав
- Рекомбинантная AAV -опосредованная инженерия генома
Ссылки
[ редактировать ]- ^ Naso MF, Tomkowicz B, Perry WL, Strohl WR (август 2017 г.). «Адено-ассоциированный вирус (AAV) как вектор для генной терапии» . Биодруги . 31 (4): 317–334. doi : 10.1007/s40259-017-0234-5 . PMC 5548848 . PMID 28669112 .
- ^ Wu Z, Yang H, Colosi P (январь 2010 г.). «Влияние размера генома на векторную упаковку AAV» . Молекулярная терапия . 18 (1): 80–86. doi : 10.1038/mt.2009.255 . PMC 2839202 . PMID 19904234 .
- ^ Grieger JC, Samulski RJ (2005). «Аденоассоцированный вирус как вектор генной терапии: развитие вектора, производство и клиническое применение». Адено-ассоциированный вирус как вектор генной терапии: развитие вектора, производство и клиническое применение . Достижения в области биохимической инженерии/биотехнологии. Тол. 99. С. 119–45. doi : 10.1007/10_005 . ISBN 978-3-540-28404-8 Полем PMID 16568890 .
- ^ Deyle Dr, Russell DW (август 2009 г.). «Аденоассоциированная вирусная векторная интеграция» . Современное мнение о молекулярной терапии . 11 (4): 442–7. PMC 2929125 . PMID 19649989 .
- ^ Jump up to: а беременный Магуайр А.М., Симонелли Ф., Пирс Э.А., Пью Эн, Мингоцци Ф., Бенничелли Дж. И др. (Май 2008 г.). «Безопасность и эффективность переноса генов для врожденного амаредоза Либер » » Журнал медицины Новой Англии 358 (21): 2240–8 Doi : 10.1056/ nejmo0802315 2829748PMC 18441370PMID
- ^ Jump up to: а беременный Tacke F (май 2023). «Тяжелая вспышка гепатита у детей, связанная с вирусом AAV2». Природа . 617 (7961): 471–472. Bibcode : 2023natur.617..471t . doi : 10.1038/d41586-023-00570-8 . PMID 36997704 .
- ^ Servellita V, Sotomayor Gonzalez A, Lamson DM, Foresythe A, Huh HJ, Bazinet AL, et al. (Май 2023). «Аденоассоциированный вирус типа 2 у детей с острым тяжелым гепатитом» . Природа . 617 (7961): 574–580. Bibcode : 2023natur.617..574S . doi : 10.1038/s41586-023-05949-1 . PMC 10170441 . PMID 36996871 .
- ^ Картер Б.Дж. (декабрь 2004 г.). «Аденоассоциированный вирус и развитие аденоассоциированных вирусных векторов: историческая перспектива» . Молекулярная терапия . 10 (6): 981–9. doi : 10.1016/j.ymthe.2004.09.011 . PMID 15564130 .
- ^ Kotin RM, Siniscalco M, Samulski RJ, Zhu XD, Hunter L, Laughlin CA, et al. (Март 1990 г.). «Специфичная для сайта интеграция с помощью аденоассоциируемого вируса» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (6): 2211–5. Bibcode : 1990pnas ... 87.2211k . doi : 10.1073/pnas.87.6.2211 . PMC 53656 . PMID 2156265 .
- ^ Супоски Р.Т., Урабе М., Годвин С.Г., МакККИСТОН С.А., Курцман Г.Дж., Озава К. и др. (Октябрь 1997). «Аденоассоциированные вирусные репутации протесты, нацеленные на последовательности ДНК на уникальный локус в геноме человека » Журнал вирусологии 71 (10): 7951–9 Doi : 10.1128/ jvi.71.10.7951-7959.1 192153PMC PMID 9311886
- ^ Jump up to: а беременный Daya S, Berns Ki (октябрь 2008 г.). «Генная терапия с использованием адено-ассоциированных вирусных векторов» . Клинические обзоры микробиологии . 21 (4): 583–93. doi : 10.1128/cmr.00008-08 . PMC 2570152 . PMID 188544481 .
- ^ Chirmule N, Propert K, Magosin S, Qian Y, Qian R, Wilson J (сентябрь 1999). «Иммунные реакции на аденовирус и адено, ассоциированный вирус, у людей» . Генная терапия . 6 (9): 1574–83. doi : 10.1038/sj.gt.3300994 . PMID 10490767 .
- ^ Эрнандес Й.Дж., Ван Дж, Кернс В.Г., Лоулер С., Пуарье А., Флотт Тр (октябрь 1999). «Латентная адено-ассоциированная вирусная инфекция вызывает гуморальную, но не клеточную иммунную реакцию в нечеловеческой модели приматов» . Журнал вирусологии . 73 (10): 8549–58. doi : 10.1128/JVI.73.10.8549-8558.1999 . PMC 112875 . PMID 10482608 .
- ^ Поннажаган С., Мукерджи П., Йодер М.К., Ван Хс, Чжоу С.З., Каплан Дж. И др. (Апрель 1997). «Аденоассоциированный вирус 2-опосредованный перенос генов in vivo: орган-триропизм и экспрессия трансдуцированных последовательностей у мышей». Ген . 190 (1): 203–10. doi : 10.1016/s0378-1119 (96) 00576-8 . PMID 9185868 .
- ^ Bak Ro, Porteus MH (июль 2017 г.). «CRISPR-опосредованная интеграция больших генов с использованием донорских векторов AAV» . Сотовые отчеты . 20 (3): 750–756. doi : 10.1016/j.celrep.2017.06.064 . PMC 5568673 . PMID 28723575 .
- ^ McCarty DM, Monahan PE, Samulski RJ (август 2001 г.). «Самокомпьютационные рекомбинантные адено-ассоциированные вирусные векторы (SCAAV) способствуют эффективной трансдукции независимо от синтеза ДНК» . Генная терапия . 8 (16): 1248–54. doi : 10.1038/sj.gt.3301514 . PMID 11509958 .
- ^ Роджерс Г.Л., Мартино А.Т., Золотухин I, Эртл Х.К., Герцог Р.В. (январь 2014 г.). «Роль векторного генома и лежащий в основе мутации фактора IX в иммунных реакциях на генную терапию AAV для гемофилии В» . Журнал трансляционной медицины . 12 : 25. DOI : 10.1186/1479-5876-12-25 . PMC 3904690 . PMID 24460861 .
- ^ «Векторы, используемые в клинических испытаниях генной терапии» . Журнал генной медицины . Уайли. Декабрь 2018 года. Архивировано с оригинала 21 октября 2019 года . Получено 4 января 2012 года .
- ^ Bainbridge JW, Smith AJ, Barker SS, Robbie S, Henderson R, Balaggan K, et al. (Май 2008 г.). «Влияние генной терапии на визуальную функцию при врожденном амаурозе Лебера». Новая Англия Журнал медицины . 358 (21): 2231–9. Citeseerx 10.1.1.574.4003 . doi : 10.1056/nejmoa0802268 . PMID 18441371 .
- ^ Hauswirth WW, немецкий TS. (Октябрь 2008 г.). Горные результаты испытания фазы I После генной терапии 19 (10) (10): 979–90. doi : 10.1089/hum . PMC 2940541 . PMID 18744912 .
- ^ Наттвани А.С., Тудденхэм Э.Г., Рангараджан С., Росалес С., Макинтош Дж., Линч Д.К. и др. (Декабрь 2011 г.). «Аденовирус, ассоциированная с вирусным вектором-опосредованным переносом гена при гемофилии В» . Новая Англия Журнал медицины . 365 (25): 2357–65. doi : 10.1056/nejmoa1108046 . PMC 3265081 . PMID 22149959 .
- ^ Джессап М., Гринберг Б., Манчини Д., Каппола Т., Поли Д.Ф., Яски Б. и др. (Июль 2011). «Повышенная регуляция кальция за счет чрескожного введения генной терапии при сердечных заболеваниях (Cupid): фаза 2 исследования внутрикоронной генной терапии саркоплазматической ретикулума Ca2+-атпазы у пациентов с активной сердечной недостаточностью» . Циркуляция . 124 (3): 304–13. doi : 10.1161/circulationaha.111.022889 . PMC 5843948 . PMID 21709064 .
- ^ «Avexis сообщает данные о продолжающемся исследовании фазы 1 AVXS-101 в позвоночнике мышечной атрофии типа 1» . Авексис . 6 мая 2016 года. Архивировано с оригинала 11 июня 2017 года . Получено 8 мая 2016 года .
- ^ Gaudet D, Méthot J, Déry S, Brisson D, Essiembre C, Tremblay G, et al. (Апрель 2013). «Эффективность и долгосрочная безопасность генной терапии алипогеном Typarvovec (AAV1-LPLS447X) для дефицита липопротеиновой липазы: открытое исследование» . Генный тур . 20 (4): 361–9. doi : 10.1038/gt.2012.43 . PMC 4956470 . PMID 22717743 .
- ^ Lewitt PA, Rezai AR, Leehey MA, Ojemann SG, Flaherty AW, Eskandar EN, et al. (Апрель 2011). «Генная терапия AAV2-GAD для развитой болезни Паркинсона: двойное слепое, контролируемое фиктивной хирургией, рандомизированное исследование». Lancet. Неврология . 10 (4): 309–19. doi : 10.1016/s1474-4422 (11) 70039-4 . PMID 21419704 . S2CID 37154043 .
- ^ Картер Б.Дж. (май 2005 г.). «Аденоассоциированные вирусные векторы в клинических испытаниях». Генная терапия человека . 16 (5): 541–50. doi : 10.1089/hum.2005.16.541 . PMID 15916479 .
- ^ Kaplitt MG, Feigin A, Tang C, Fitzsimons HL, Mattis P, Lawlor PA, et al. (Июнь 2007 г.). «Безопасность и переносимость генной терапии с помощью аденоассоциированного вируса (AAV) GAD GENE для болезни Паркинсона: открытый этикетка, исследование I фазы». Лансет . 369 (9579): 2097–105. doi : 10.1016/s0140-6736 (07) 60982-9 . PMID 17586305 . S2CID 24491886 .
- ^ Картер Б.Дж. (2000). «Аденоассоциированные вирус и аденоассоциированные вирусные векторы для доставки генов». В Lassic DD, Templeton NS (Eds.). Генная терапия: терапевтические механизмы и стратегии . Нью -Йорк: Marcel Dekker, Inc. с. 41–59. ISBN 978-0-585-39515-9 .
- ^ Bohenzky RA, Lefebvre RB, Berns Ki (октябрь 1988 г.). «Требования к последовательности и симметрии во внутренних палиндромических последовательностях аденоассоциированного вирусного терминального повторения». Вирусология . 166 (2): 316–27. doi : 10.1016/0042-6822 (88) 90502-8 . PMID 2845646 .
- ^ Wang XS, Ponnazhagan S, Srivastava A (июль 1995 г.). «Скажи и репликации генома адено-ассоциированного вируса 2». Журнал молекулярной биологии . 250 (5): 573–80. doi : 10.1006/jmbi.1995.0398 . PMID 7623375 .
- ^ Jump up to: а беременный Weitzman MD, Kyöstiö SR, Kotin RM, Owens RA (июнь 1994 г.). «Белки Rep Adeno-ассоциированного вируса (AAV) опосредуют комплексное образование между ДНК AAV и его сайтом интеграции в ДНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (13): 5808–12. Bibcode : 1994pnas ... 91.5808w . doi : 10.1073/pnas.91.13.5808 . PMC 44086 . PMID 8016070 .
- ^ Zhou X, Muzyczka N (апрель 1998 г.). «Упаковка in vitro Адено-акцентированной Рейрус ДНК » Журнал вирусологии 72 (4): 3241–7 Doi : 10.1128/ jvi.72.4.3241-3247.1 109794PMC 9525651PMID
- ^ Nony P, Tessier J, Chadeuf G, Ward P, Giraud A, Dugast M, et al. (Октябрь 2001 г.). «Новый цис-действительный элемент репликации в адено-ассоциированном геноме типа 2 вируса участвует в амплификации интегрированных последовательностей Rep-CAP» . Журнал вирусологии . 75 (20): 9991–4. doi : 10.1128/JVI.75.20.9991-9994.2001 . PMC 114572 . PMID 11559833 .
- ^ Nony P, Chadeuf G, Tessier J, Moullier P, Salvetti A (январь 2003 г.). «Свидетельство о упаковке последовательностей REP-CAP в аденоассоциированные вирусные капсиды типа 2 в отсутствие перевернутых терминальных повторений: модель для генерации реп-позитивных частиц AAV» . Журнал вирусологии . 77 (1): 776–81. doi : 10.1128/jvi.77.1.776-781.2003 . PMC 140600 . PMID 12477885 .
- ^ Philpott NJ, Giraud-Wali C, Dupuis C, Gomos J, Hamilton H, Berns Ki, et al. (Июнь 2002 г.). «Эффективная интеграция рекомбинантных адено-ассоциированных векторов ДНК-вирусной ДНК требует последовательности P5-REP в цис» . Журнал вирусологии . 76 (11): 5411–21. doi : 10.1128/jvi.76.11.5411-5421.2002 . PMC 137060 . PMID 11991970 .
- ^ Tullis GE, Shenk T (декабрь 2000 г.). «Эффективная репликация векторов адено-ассоциированного вируса типа 2: цис-действие за пределами терминальных повторений и минимальный размер» . Журнал вирусологии . 74 (24): 11511–21. doi : 10.1128/JVI.74.24.11511-11521.2000 . PMC 112431 . PMID 11090148 .
- ^ Jump up to: а беременный Kyöstiö Sr, Owens RA, Weitzman MD, Antoni BA, Chejanovsky N, Carter BJ (май 1994). «Анализ адено-ассоциированных вирусных (AAV) белков дикого типа и мутанта для их способностей негативно регулировать уровни мРНК AAV P5 и P19» . Журнал вирусологии . 68 (5): 2947–57. doi : 10.1128/JVI.68.5.2947-2957.1994 . PMC 236783 . PMID 8151765 .
- ^ Im ds, muzyczka n (май 1990). «Белок, связывающий происхождение AAV Rep68, является АТФ-зависимой сайт-специфической эндонуклеазой с активностью ДНК геликазы». Клетка . 61 (3): 447–57. doi : 10.1016/0092-8674 (90) 90526-K . PMID 2159383 . S2CID 27997617 .
- ^ Im DS, Muzyczka N (февраль 1992 г.). «Частичная очистка аденоассоциированного вируса Rep78, Rep52 и Rep40 и их биохимической характеристики» . Журнал вирусологии . 66 (2): 1119–28. doi : 10.1128/jvi.66.2.1119-1128.1992 . PMC 240816 . PMID 1309894 .
- ^ Самулски Р.Дж. (2003). «Векторы AAV, будущая рабочая лошадка человеческой генной терапии». Эрнст Шеринговый Исследовательский Фонд . Человеческая генная терапия: современные возможности и будущие тенденции. С. 25–40. doi : 10.1007/978-3-662-05352-2_3 . ISBN 978-3-662-05354-6 Полем PMID 12894449 .
- ^ Trempe JP, Carter BJ (январь 1988 г.). «Регуляция экспрессии гена адено-ассоциированного вируса в 293 клетках: контроль численности мРНК и трансляции» . Журнал вирусологии . 62 (1): 68–74. doi : 10.1128/jvi.62.1.68-74.1988 . PMC 250502 . PMID 2824856 .
- ^ Jay Ft, Laughlin CA, Carter BJ (май 1981). «Эукариотический трансляционный контроль: аденоассоцированный вирусный белок влияет мутация в аденовирусном ДНК-связывающем белке» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (5): 2927–31. Bibcode : 1981pnas ... 78.2927j . doi : 10.1073/pnas.78.5.2927 . PMC 319472 . PMID 6265925 .
- ^ Sonntag F, Schmidt K, Kleinschmidt JA (июнь 2010 г.). «Коэффициент вирусной сборки способствует образованию капсида AAV2 в ядреве» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (22): 10220–5. Bibcode : 2010pnas..10710220S . doi : 10.1073/pnas.1001673107 . PMC 2890453 . PMID 20479244 .
- ^ Pierson EE, Keifer DZ, Asokan A, Jarrold MF (июль 2016 г.). «Решение аденоассоциированного разнообразия вирусных частиц с масс-спектрометрией обнаружения заряда» . Аналитическая химия . 88 (13). Американское химическое общество (ACS): 6718–6725. doi : 10.1021/acs.analchem.6b00883 . PMC 6537880 . PMID 27310298 .
- ^ Xie Q, Bu W, Bhatia S, Hare J, Somasundaram T, Azzi A, et al. (Август 2002). «Атомная структура адено-ассоциированного вируса (AAV-2), вектор для генной терапии человека» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (16): 10405–10. Bibcode : 2002pnas ... 9910405x . doi : 10.1073/pnas.162250899 . PMC 124927 . PMID 12136130 .
- ^ Sonntag F, Köther K, Schmidt K, Weghofer M, Raupp C, Nieto K, et al. (Декабрь 2011 г.). «Активирующий сборник белок способствует капсидной сборке различных аденоассоциированных вирусных серотипов» . Журнал вирусологии . 85 (23): 12686–97. doi : 10.1128/jvi.05359-11 . PMC 3209379 . PMID 21917944 .
- ^ Becerra SP, Rose JA, Hardy M, Baroudy BM, Anderson CW (декабрь 1985 г.). «Прямое картирование адено-ассоциированных вирусных капсидных белков B и C: возможный кодон инициации ACG» . Труды Национальной академии наук Соединенных Штатов Америки . 82 (23): 7919–23. Bibcode : 1985pnas ... 82.7919b . doi : 10.1073/pnas.82.23.7919 . PMC 390881 . PMID 2999784 .
- ^ Cassinotti P, Weitz M, Tratschin JD (ноябрь 1988). «Организация капсидного гена адено-ассоциированного вируса (AAV): картирование незначительной сплайсированной мРНК, кодирующей вирусной капсидный белок 1». Вирусология . 167 (1): 176–84. doi : 10.1016/0042-6822 (88) 90067-0 . PMID 2847413 .
- ^ Jump up to: а беременный Muralidhar S, Becerra SP, Rose Ja (январь 1994 г.). «Сайт-направленный мутагенез адено-ассоциированных вируса типа 2 структурного белка кодонов: влияние на регуляцию синтеза и биологической активности» . Журнал вирусологии . 68 (1): 170–6. doi : 10.1128/jvi.68.1.170-176.1994 . PMC 236275 . PMID 8254726 .
- ^ Trempe JP, Carter BJ (сентябрь 1988 г.). «Альтернативный сплайсинг мРНК необходим для синтеза аденоассоциированного капсидного белка VP1-вируса» . Журнал вирусологии . 62 (9): 3356–63. doi : 10.1128/jvi.62.9.3356-3363.1988 . PMC 253458 . PMID 2841488 .
- ^ Rabinowitz JE, Samulski RJ (декабрь 2000 г.). «Построение лучшего вектора: манипуляции с вирионами AAV» . Вирусология . 278 (2): 301–8. doi : 10.1006/viro.2000.0707 . PMID 11118354 .
- ^ Джирод А., Вобус К.Е., Зайдори З., Рид М., Лейк К., Тижссен П. и др. (Май 2002 г.). «Капсидный белок VP1 с аденоассоцированным вирусом типа 2 несет домен фосфолипазы A2, необходимый для инфекции вируса» . Журнал общей вирусологии . 83 (Pt 5): 973–8. doi : 10.1099/0022-1317-83-5-973 . PMID 11961250 .
- ^ Уоррингтон К.Х., Горбатюк О.С., Харрисон Дж.К., Опи С.Р., Золотухин С., Музицка Н. (июнь 2004 г.). «Аденоассоцированный капсидный белок VP2 типа 2 типа 2 типа является несущественным и может переносить большие пептидные вставки на его n-конце» . Журнал вирусологии . 78 (12): 6595–609. doi : 10.1128/JVI.78.12.6595-6609.2004 . PMC 416546 . PMID 15163751 .
- ^ Rumachik Ng, Malaker SA, Poweleit N, Maynard LH, Adams CM, Leib RD, et al. (Май 2020). «Методы имеют значение: стандартные производственные платформы для рекомбинантного AAV производят химически и функционально различные векторы» . Молекулярная терапия - методы и клиническая разработка . 18 : 98–118. doi : 10.1016/j.omtm.2020.05.018 . PMC 7488757 . PMID 32995354 .
- ^ Chen Q, Njenga R, Leuchs B, Chiocca S, Kleinschmidt J, Muller M (сентябрь 2020 г.). «Сумоилирование нацелена на адено, ассоциированные с вирусными капсидами, но в основном ограничивает трансдукцию клеточными механизмами» . Журнал вирусологии . 94 (19): E00871-20. doi : 10.1128/jvi.00871-20 . PMC 7495375 . PMID 32669341 .
- ^ Джайлс А.Р., Симс Дж.Дж., Тернер К.Б., Говиндасами Л., Альвира М.Р., Лок М. и др. (Декабрь 2018). «Деамидирование аминокислот на поверхности адено, ассоциированных с вирусными капсидами, приводит к гетерогенности зарядки и измененной векторной функции» . Молекулярная терапия . 26 (12): 2848–2862. doi : 10.1016/j.ymthe.2018.09.013 . PMC 6277538 . PMID 30343890 .
- ^ "AAV Тропизмы" . Векторбейдер .
- ^ «Подсемейство: Parvovirinae, род: Devestoparvovirus» . Международный комитет по таксономии вирусов (ICTV) . Получено 31 января 2020 года .
- ^ Jump up to: а беременный Вейцман М.Д., Линден Р.М. (2011). «Аденоассоциированная вирусная биология». В Snyder Ro, Moullier P (Eds.). Адено-ассоциированные вирусные методы и протоколы . Тотова, Нью -Джерси: Humana Press. ISBN 978-1-61779-370-7 .
- ^ Мори С., Ван Л., Такеучи Т., Канда Т (декабрь 2004 г.). «Два новых аденоассоциированных вируса от Cynomolgus monkey: псевдотипировая характеристика капсидного белка» . Вирусология . 330 (2): 375–83. doi : 10.1016/j.virol.2004.10.012 . PMID 15567432 .
- ^ Jump up to: а беременный Bartlett JS, Samulski RJ, McCown TJ (май 1998). «Селективное и быстрое поглощение аденоассоциированного вируса типа 2 в мозге». Генная терапия человека . 9 (8): 1181–6. doi : 10.1089/hum.1998.9.8-1181 . PMID 9625257 .
- ^ Fischer AC, Beck SE, Smith CI, Laube BL, Askin FB, Guggino SE, et al. (Декабрь 2003 г.). «Успешная экспрессия трансгена с серийными дозами аэрозольных векторов RAAV2 в макаках -резусах» . Молекулярная терапия . 8 (6): 918–26. doi : 10.1016/j.ymthe.2003.08.015 . PMID 14664794 .
- ^ Никлин С.А., Буининг Х, Диарт К.Л., Де Алвис М., Джирод А., Хакер У. и др. (Сентябрь 2001 г.). «Эффективный и селективный AAV2-опосредованный перенос генов, направленный на эндотелиальные клетки сосудов человека» . Молекулярная терапия . 4 (3): 174–81. doi : 10.1006/mthe.2001.0424 . PMID 11545607 .
- ^ Rabinowitz JE, Xiao W, Samulski RJ (декабрь 1999 г.). «Инсервосный мутагенез капсида AV2 и производство рекомбинантного вируса » Вирусология 265 (2): 274–8 Doi : 10.1006/ viro.1999.0 10600599PMID
- ^ Shi W, Bartlett JS (апрель 2003 г.). «Включение RGD в VP3 обеспечивает аденоассоцированные векторы вируса типа 2 (AAV2) с гепаран-сульфатом механизмом ввода клеток» . Молекулярная терапия . 7 (4): 515–25. doi : 10.1016/s1525-0016 (03) 00042-х . PMID 12727115 .
- ^ Wu P, Xiao W, Conlon T, Hughes J, Agbandje-Mckenna M, Ferkol T, et al. (Сентябрь 2000). «Мутационный анализ адено-ассоциированного вируса типа 2 (AAV2) капсидного гена и построение векторов AAV2 с измененным тропизмом» . Журнал вирусологии . 74 (18): 8635–47. doi : 10.1128/JVI.74.18.8635-8647.2000 . PMC 102071 . PMID 10954565 .
- ^ Манно С.С., Чу А.Дж., Хатчисон С., Ларсон П.Дж., Херцог Р.В., Арруда В.Р. и др. (Апрель 2003 г.). «AAV-опосредованный фактор IX перенос гена IX в скелетные мышцы у пациентов с тяжелой гемофилией В» . Кровь . 101 (8): 2963–72. doi : 10.1182/blood-2002-10-3296 . PMID 12515715 .
- ^ Рихтер М., Ивата А., Нихуис Дж., Нитта Й., Миллер А.Д., Хэлберт К.Л. и др. (Апрель 2000). «Аденоассоциированная вирусная векторная трансдукция клеток гладких мышц сосудов in vivo». Физиологическая геномика . 2 (3): 117–27. doi : 10.1152/physiolgenomics.2000.2.3.117 . PMID 11015590 . S2CID 14150468 .
- ^ Koeberl DD, Alexander IE, Halbert CL, Russell DW, Miller AD (февраль 1997 г.). «Постоянная экспрессия угасного фактора свертывания человека из печени мыши после внутривенной инъекции аденоассоциированных векторов вируса» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (4): 1426–31. Bibcode : 1997pnas ... 94.1426K . doi : 10.1073/pnas.94.4.1426 . PMC 19807 . PMID 9037069 .
- ^ Цин К., Мах С., Хансен Дж., Чжоу С., Дварки В., Шривастава А (январь 1999 г.). «Рецептор 1-рецептора фактора роста фибробластов человека является со-рецептором инфекции аденоассоциированным вирусом 2». Природная медицина . 5 (1): 71–7. doi : 10.1038/4758 . PMID 9883842 . S2CID 31602526 .
- ^ Summerford C, Samulski RJ (февраль 1998 г.). «Мембрановый гепаран-сульфатный протео-гликан является рецептором для аденоассоциированных вирусов типа 2 типа» . Журнал вирусологии . 72 (2): 1438–45. doi : 10.1128/jvi.72.2.1438-1445.1998 . PMC 124624 . PMID 9445046 .
- ^ Summerford C, Bartlett JS, Samulski RJ (январь 1999 г.). «Alphavbeta5 Integrin: со-рецептор для адено-ассоциированного вируса инфекции типа 2». Природная медицина . 5 (1): 78–82. doi : 10.1038/4768 . PMID 9883843 . S2CID 23326070 .
- ^ Qiu J, Handa A, Kirby M, Brown Ke (март 2000 г.). «Взаимодействие гепаринового сульфата и адено-ассоциированного вируса 2» . Вирусология . 269 (1): 137–47. doi : 10.1006/viro.2000.0205 . PMID 10725206 .
- ^ Pajusola K, Gruchala M, Joch H, Lüscher TF, Ylä-Herttuala S, Büeler H (ноябрь 2002 г.). «Характеристики клеток-типа модулируют эффективность трансдукции адено-ассоциированного вируса типа 2 и ограничивают инфекцию эндотелиальных клеток» . Журнал вирусологии . 76 (22): 11530–40. doi : 10.1128/JVI.76.22.11530-11540.2002 . PMC 136795 . PMID 12388714 .
- ^ «Общий вирус« убивает рак » . CNN . 22 июня 2005 г. Получено 5 августа 2009 года .
- ^ Penn State (23 июня 2005 г.). «Безвредный вирус убивает некоторые виды рака» . Наука ежедневно.
- ^ MP Limberis, Vandenberghe LH, Zhang L, Pickles RJ, Wilson JM (февраль 2009 г.). «Эффективность трансдукции новых векторов AAV в эпителии дыхательных путей мыши in vivo и эпителия дыхательных путей человека in vitro» . Молекулярная терапия . 17 (2): 294–301. doi : 10.1038/mt.2008.261 . PMC 2835069 . PMID 19066597 .
- ^ Стробель Б., Духс М.Дж., Шмид Р., Стисторфер Б.Е., Бухер Х., Кваст К. и др. (Сентябрь 2015). «Моделирование путей легочной болезни с использованием рекомбинантного аденоассоциированного вируса 6,2». Американский журнал респираторных клеток и молекулярной биологии . 53 (3): 291–302. doi : 10.1165/rcmb.2014-0338ma . PMID 25845025 .
- ^ Gao GP, Alvira MR, Wang L, Calcedo R, Johnston J, Wilson JM (сентябрь 2002 г.). «Новые адено-ассоциированные вирусы от резус-обезьян в качестве векторов для генной терапии человека» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (18): 11854–9. Bibcode : 2002pnas ... 9911854G . doi : 10.1073/pnas.182412299 . PMC 129358 . PMID 12192090 .
- ^ Jump up to: а беременный Halbert CL, Allen JM, Miller AD (июль 2001 г.). «Аденоассоцированные векторы вируса типа 6 (AAV6) опосредуют эффективную трансдукцию эпителиальных клеток дыхательных путей в легких мышиных векторов» . Журнал вирусологии . 75 (14): 6615–24. doi : 10.1128/JVI.75.14.6615-6624.2001 . PMC 114385 . PMID 11413329 .
- ^ Jump up to: а беременный Rabinowitz JE, Bowles DE, Faust SM, Ledford JG, Cunningham SE, Samulski RJ (май 2004 г.). «Поперечная одежда вириона: транскапсидирование аденоассоциированных вирусных серотипов функционально определяет подгруппы» . Журнал вирусологии . 78 (9): 4421–32. doi : 10.1128/JVI.78.9.4421-4432.2004 . PMC 387689 . PMID 15078923 .
- ^ Chen S, Kapturczak M, Loiler SA, Zolotukhin S, Glushakova Oy, Madsen KM, et al. (Февраль 2005 г.). «Эффективная трансдукция эндотелиальных клеток сосудов с рекомбинантным аденоассоцированным вирусным серотипом 1 и 5 векторов» . Генная терапия человека . 16 (2): 235–47. doi : 10.1089/hum.2005.16.235 . PMC 1364465 . PMID 15761263 .
- ^ Ортинский Пи, Донг Дж., Мунгенаст А., Юэ С., Такано Х., Уотсон Д.Дж. и др. (Май 2010). «Селективная индукция астроцитарного глиоза генерирует дефицит при ингибировании нейронов» . Nature Neuroscience . 13 (5): 584–91. doi : 10.1038/nn.2535 . PMC 3225960 . PMID 20418874 .
- ^ Калудов Н., Браун К.Е., Уолтерс Р.В., Забнер Дж., Чиорини Дж.А. (август 2001 г.). «Адено-ассоциированный вирусный серотип 4 (AAV4) и AAV5 требуют связывания сиаловой кислоты для гемагглютинации и эффективной трансдукции, но отличаются по специфичности связи сиаловой кислоты» . Журнал вирусологии . 75 (15): 6884–93. doi : 10.1128/JVI.75.15.6884-6893.2001 . PMC 114416 . PMID 11435568 .
- ^ Di Pasquale G, Davidson BL, Stein CS, Martins I, Scudiero D, Monks A, et al. (Октябрь 2003 г.). «Идентификация PDGFR в качестве рецептора для трансдукции AAV-5» . Природная медицина . 9 (10): 1306–12. doi : 10.1038/nm929 . PMID 14502277 . S2CID 32547188 .
- ^ «Адено-ассоциированный гид вируса (AAV)» . Addgene . Уотертаун, Массачусетс.
- ^ Zinn E, Pacouret S, Khaychuk V, Turunen HT, Carvalho LS, Andres-Mateos E, et al. (Август 2015). «При реконструкции силико эволюционной линии вирусной эволюции дает мощный вектор генной терапии» . Сотовые отчеты . 12 (6): 1056–68. doi : 10.1016/j.celrep.2015.07.019 . PMC 4536165 . PMID 26235624 .
- ^ Flotte TR, Berns Ki (апрель 2005 г.). «Адено-ассоциированный вирус: вездесущий комменсал млекопитающих». Генная терапия человека . 16 (4): 401–407. doi : 10.1089/hum.2005.16.401 . PMID 15871671 .
- ^ Ронцитти Г., Гросс Д.А., Мингоцци Ф (2020). «Иммунные реакции человека на адено, ассоциированные с вирусами (AAV) векторы» . Границы в иммунологии . 11 : 670. DOI : 10.3389/fimmu.2020.00670 . PMC 7181373 . PMID 32362898 .
- ^ Вандамм С., Аджали О, Мингоцци Ф (ноябрь 2017). «Раскрытие сложной истории иммунных реакций на испытание векторов AAV после испытания» . Генная терапия человека . 28 (11): 1061–1074. doi : 10.1089/hum.2017.150 . PMC 5649404 . PMID 28835127 .
- ^ Barnes C, Scheideler O, Schaffer D (декабрь 2019 г.). «Инженерная капсид AAV уклониться от иммунных ответов» . Текущее мнение о биотехнологии . 60 : 99–103. doi : 10.1016/j.copbio.2019.01.002 . PMC 7117822 . PMID 30807882 .
- ^ Zaiss AK, Liu Q, Bowen GP, Wong NC, Bartlett JS, Muruve DA (май 2002). «Дифференциальная активация врожденных иммунных реакций аденовирусом и аденоассоциированными вирусными векторами» . Журнал вирусологии . 76 (9): 4580–90. doi : 10.1128/JVI.76.9.4580-4590.2002 . PMC 155101 . PMID 11932423 .
- ^ Чжу Дж, Хуан Х, Ян Y (август 2009 г.). «Путь TLR9-MYD88 является критическим для адаптивных иммунных реакций на аденоассоциированные векторы генной терапии вируса у мышей» . Журнал клинических исследований . 119 (8): 2388–2398. doi : 10.1172/jci37607 . PMC 2719948 . PMID 19587448 .
- ^ Martino AT, Suzuki M, Markusic DM, Zolotukhin I, Ryals RC, Moghimi B, et al. (Июнь 2011 г.). «Геном самодополнительных адено-ассоциированных вирусных векторов увеличивает Toll-подобные рецептор 9-зависимых врожденных иммунных реакций в печени» . Кровь . 117 (24): 6459–6468. doi : 10.1182/blood-2010-10-314518 . PMC 3123017 . PMID 21474674 .
- ^ Calcedo R, Morizono H, Wang L, McCarter R, He J, Jones D, et al. (Сентябрь 2011). «Аденоассоциированные профили вирусных антител у новорожденных, детей и подростков» . Клиническая и вакцинная иммунология . 18 (9): 1586–1588. doi : 10.1128/cvi.05107-11 . PMC 3165215 . PMID 21775517 .
- ^ TSE LV, Klinc KA, Madigan VJ, Castellanos Rivera RM, Wells LF, Havlik LP, et al. (Июнь 2017 г.). «Структурная эволюция антигенных различных аденоассоцированных вариантов вируса для иммунной уклонения» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (24): E4812 - E4821. BIBCODE : 2017PNAS..114E4812T . doi : 10.1073/pnas.17047661114 . PMC 5474820 . PMID 28559317 .
- ^ Jump up to: а беременный Zaiss AK, Muruve DA (июнь 2005 г.). «Иммунные ответы на аденоассоциированные вирусные векторы». Современная генная терапия . 5 (3): 323–31. doi : 10.2174/1566523054065039 . PMID 15975009 .
- ^ Jump up to: а беременный в Мэдсен Д., Кантвелл Э.Р., О'Брайен Т., Джонсон П.А., Махон Б.П. (ноябрь 2009 г.). «Адено-ассоциированный вирусный серотип 2 индуцирует клеточные иммунные ответы, направленные на множественные эпитопы капсидного белка VP1» . Журнал общей вирусологии . 90 (Pt 11): 2622–33. doi : 10.1099/vir.0.014175-0 . PMC 2885037 . PMID 19641045 .
- ^ Jump up to: а беременный Manno CS, Pierce GF, Arruda VR, Glader B, Ragni M, Rasko JJ, et al. (Март 2006 г.). «Успешная трансдукция печени при гемофилии с помощью AAV-Factor IX и ограничения, налагаемые иммунным ответом хозяина». Природная медицина . 12 (3): 342–7. doi : 10.1038/nm1358 . PMID 16474400 . S2CID 14664182 .
- ^ Sabatino DE, Mingozzi F, Hui DJ, Chen H, Colosi P, Ertl HC, et al. (Декабрь 2005 г.). «Идентификация мышиных капсид-капсид-специфических эпитопов CD8+ T-клеток» . Молекулярная терапия . 12 (6): 1023–33. doi : 10.1016/j.ymthe.2005.09.009 . PMID 16263332 .
- ^ Rohr Up, Kronenwett R, Grimm D, Kleinschmidt J, Haas R (сентябрь 2002 г.). «Первичные клетки человека отличаются по их восприимчивости к RAAV-2-опосредованному переносу генов и продолжительности экспрессии репортерного гена». Журнал вирусологических методов . 105 (2): 265–75. doi : 10.1016/s0166-0934 (02) 00117-9 . PMID 12270659 .
- ^ Jump up to: а беременный Matsushita T, Elliger S, Elliger C, Podsakoff G, Villarreal L, Kurtzman GJ, et al. (Июль 1998 г.). «Аденоассоциированные вирусные векторы могут эффективно продуцироваться без вируса помощника» . Генная терапия . 5 (7): 938–45. doi : 10.1038/sj.gt.3300680 . PMID 9813665 .
- ^ Myers MW, Laughlin CA, Jay Ft, Carter BJ (июль 1980 г.). «Аденовирусная вспомогательная функция для роста аденоассоциированного вируса: эффект чувствительных к температуре мутаций в аденовирусной области ранней генной области 2» . Журнал вирусологии . 35 (1): 65–75. doi : 10.1128/jvi.35.1.65-75.1980 . PMC 288783 . PMID 6251278 .
- ^ Ханда Х, Картер Б.Дж. (июль 1979 г.). «Аденоассоцированные комплексы репликации ДНК вируса у вируса простого герпеса или клеток, инфицированных аденовирусом» . Журнал биологической химии . 254 (14): 6603–10. doi : 10.1016/s0021-9258 (18) 50411-7 . PMID 221504 .
- ^ Yalkinoglu Ao, Heilbronn R, Bürkle A, Schlehofer Jr, Zur Hausen H (июнь 1988 г.). «Амплификация ДНК адено-ассоциированного вируса как ответ на клеточный генотоксический стресс» . РАНКА . 48 (11): 3123–9. PMID 2835153 .
- ^ Якобсон Б., Кох Т, Винокур Е (апрель 1987 г.). «Репликация адено-ассоциированного вируса в синхронизированных клетках без добавления вспомогательного вируса» . Журнал вирусологии . 61 (4): 972–81. doi : 10.1128/jvi.61.4.972-981.1987 . PMC 254052 . PMID 3029431 .
- ^ Якобсон Б., Хринко Т.А., Пик М.Дж., Винокур Е (март 1989 г.). «Репликация адено-ассоциированного вируса в клетках, облученных ультрафиолетовым светом при 254 нм» . Журнал вирусологии . 63 (3): 1023–30. doi : 10.1128/JVI.63.3.1023-1030.1989 . PMC 247794 . PMID 2536816 .
- ^ «Введение в адено-ассоциированный вирус (AAV)» . Векторные биол . Малверн, Пенсильвания.
- ^ «Аденоассоциированные вирусы и аденоассоциированные вирусные векторы» . Здоровье окружающей среды и безопасность . Университет Айовы. Архивировано с оригинала 20 сентября 2018 года . Получено 19 сентября 2018 года .
- ^ Duan D, Sharma P, Yang J, Yue Y, Dudus L, Zhang Y, et al. (Ноябрь 1998). «Круглые промежуточные соединения рекомбинантного адено-ассоциированного вируса определяют структурные характеристики, ответственные за долгосрочную эпизомальную стойкость в мышечной ткани» . Журнал вирусологии . 72 (11): 8568–77. doi : 10.1128/jvi.72.11.8568-8577.1998 . PMC 110267 . PMID 9765395 .
Внешние ссылки
[ редактировать ]- Kimball JW (17 мая 2015 г.). «Генная терапия II» . Страницы биологии Кимбалла . Архивировано из оригинала 18 марта 2005 года . Получено 13 мая 2005 года .
- « Адено-ассоциированный вирус » . NCBI таксономия браузер . 272636.