Экстрахромосомная круговая ДНК
Экстрахромосомная круговая ДНК ( экцДНК ) представляет собой тип двухцепочечной круглой структуры ДНК, который был впервые обнаружен в 1964 году Аликс Бассель и Ясуо Хотта. [ 1 ] В отличие от ранее идентифицированных структур круглых ДНК (например, бактериальные плазмиды , митохондриальную ДНК , круглые бактериальные хромосомы или хлоропластская ДНК ), экцДНК являются круглыми ДНК в эукариотических ядрах растений и животных (включая человека). Экстрахромосомная круговая ДНК получена из хромосомной ДНК, может варьироваться в размере от 50 пар оснований до нескольких мега-пар длиной в длину и может кодировать регуляторные элементы и полноразмерные гены . ЭккДНК наблюдалась у различных эукариотических видов [ 2 ] [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ] и предполагается, что это побочный продукт запрограммированных событий рекомбинации ДНК , таких как рекомбинация V (D) J. [ 8 ] [ 9 ]
Исторический фон
[ редактировать ]В 1964 году Бассель и Хотта опубликовали свое первоначальное открытие экдны, которое они сделали, исследуя Франклина Шталя . хромосомную теорию [ 10 ] В своих экспериментах они визуализировали изолированные ядра пшеницы и кабану с помощью электронной микроскопии . [ 10 ] Их исследование показало, что кабаловые сперматозоиды содержали эккДНК различных размеров. [ 10 ] В 1965 году исследовательская группа Артура Сприггса идентифицировала экцДНК в образцах эмбриональных опухолей пяти педиатрических пациентов и бронхиальной карциномы одного взрослого пациента . [ 11 ] В последующие годы дополнительные исследования привели к открытию экцДНК в различных видах, перечисленных в таблице 1:
| Год | Организм | Ссылка |
|---|---|---|
| 1965 | Кабана сперма | Hotta and Bassel, 1965 [ 10 ] |
| 1965 | Опухоли человека | Cox et al., 1965 [ 11 ] |
| 1969 | Дрожжи | Billheimer и Avers, 1969 [ 12 ] |
| 1984 | Трипаносоматиды | Beverly et al., 1984 [ 13 ] |
| 1972 | Евглена | Насс и Бен-Шаул, 1972 [ 14 ] |
| 1972 | Табак | Wong and Wildman, 1972 [ 15 ] |
| 1972, 1978, 1980 | Грибы | Agsteribbe et al., 1972; [ 16 ] Stahl et al., 1978; [ 17 ] Lazarus et al., 1980 [ 18 ] |
| 1972, 1985 | Культурные человеческие фибробласты | Смит и Виноград, 1972; [ 19 ] Riabowol et al., 1985 [ 4 ] |
| 1976 | Xenopus | Доброе утро-Нарделли и др., 1976 [ 20 ] |
| 1978, 1984 | Цыпленка Бурса | Delap and Rush, 1978; [ 21 ] Тода и Ямагиши, 1984 [ 22 ] |
| 1982 | Человеческие ткани | Calabretta et al., 1982 [ 23 ] |
| 1983 | Эмбрион мыши | Yamagishi et al., 1983 [ 24 ] |
| 1983, 1988, 1990 | Мышиные ткани | Tsuda et al., 1983; [ 25 ] Flores et al., 1988; [ 26 ] Gaubatz and Flores, 1990 [ 2 ] |
| 1983 | Мышиные тимоциты | Yamagishi et al., 1983 [ 24 ] |
| 1983 | Мышиные лимфоциты | Tsuda et al., 1983 [ 26 ] |
Исследование 21 -го века
[ редактировать ]В 21 -м веке исследователи сосредоточились на лучшей характеристике специфических подтипов экцДНК, а также на структуре и функции этих молекул в биологических системах: [ 27 ]
- В 2012 году Shibata et al. обнаружил специфический тип эккДНК, называемую микродной . [ 6 ] Исследователи обнаружили десятки тысяч микродн в тканях мыши и клеточных линиях, а также клеточные линии человека. [ 6 ]
- В 2017 году Turner et al. Идентифицируется с использованием секвенирования всего генома (WGS), цитогенетического анализа и структурного моделирования, что экстрахромосомная круговая ДНК очень амплифицирована и распространена при различных типах рака . [ 28 ] Они обнаружили, что молекулы эккДНК обладают значительной гетерогенностью между различными клетками, даже если они получены из одного и того же человека. [ 28 ] Кроме того, эти молекулы эккДНК содержали гонщики с опухолью и, как сообщалось, редко обнаруживаются в нездоровых тканях. [ 28 ]
- В 2018 году Møller et al. Использовали здоровые образцы мышц человека и клетки крови для выявления более 100 000 типов экцДНК, что предполагало, что экцДНК может быть обнаружена в соматических клетках повсеместно. [ 29 ]
- В 2019 году Wu et al. обнаружил, что экдна (подтип эккДНК) ассоциируется с хроматином , но в отличие от хромосом, он не имеет уплотнения более высокого порядка, что увеличивает его доступность. [ 30 ]
- В 2021 году Wang et al. подробно описывается на формирование ECCDNAS и идентифицировала иммуностимуляторную функцию ECCDNAS. [ 31 ] Они также разработали улучшенный протокол очистки эккДНК, который уменьшает линейную загрязнение ДНК в очищенных образцах. [ 31 ]
Очистка эккДНК
[ редактировать ]Исторически, ECCDNA очищали с использованием двухэтапной процедуры, которая включала первую изолирующую неочищенную экстрахромосомную ДНК и впоследствии переваривает линейную ДНК посредством экзонуклеазы расщепления . [ 31 ] Тем не менее, этот метод часто приводит к линейному загрязнению ДНК, поскольку расщепление экзонуклеазы недостаточно для удаления всей линейной ДНК. [ 31 ] В 2021 году Wang et al. разработал трехступенчатый метод обогащения эккДНК, который улучшил очистку экцДНК: [ 31 ]
- Клетки сначала дегидратировали в> 90% метаноле. Для извлечения неочищенной экстрахромосомной ДНК клетки лизировали с pH 11,8 щелочным лизисным буфером, нейтрализовали с нейтрализационным буфером и осаждали с использованием буфера осадков. Коммерческий набор для очистки плазмиды была использована для выделения ДНК из других клеточных компонентов.
- Элюированную ДНК расщепляли с помощью рестрикционного фермента PACI для линеаризации митохондриальной ДНК ( мтДНК ) и экзонуклеазы, которая может переваривать линейную ДНК.
- Наконец, круглая ДНК избирательно извлекали коммерческим раствором и шариками кремнезема для удаления линейной ДНК, которая не была удалена экзонуклеазной расщеплением.
Двойные минуты (дм) против экстрахромосомной круглой ДНК (экцДНК)
[ редактировать ]Первоначально термин «двойные минуты » (DM) обычно использовался для обозначения экстрахромосомной круглой ДНК, потому что он часто появлялся как пара в ранних исследованиях. [ 27 ] Поскольку исследования продолжались, были идентифицированы различные подтипы экстрахромосомной круглой ДНК, которые не являются двойными минутами (например, микродна ). В 2014 году Barreto et al. определил, что двойные минуты составляют только примерно 30% экстрахромосомной ДНК. [ 32 ] Таким образом, термин экстрахромосомной круглой ДНК (экцДНК) становится все более широко используемым, в то время как термин двойной минуты теперь зарезервирован для определенного подтипа эккДНК. [ 32 ]
Структура
[ редактировать ]ЭккДНК представляют собой круглую ДНК, которые были обнаружены в клетках человека, растений и животных и присутствуют в клеточном ядре в дополнение к хромосомной ДНК . ЭккДНК отличается от другой круглой ДНК в клетках, таких как митохондриальная ДНК (мтДНК), потому что она колеблется в размерах от нескольких сотен оснований до мегабаз и получена из геномной ДНК. [ 1 ] Например, эккДНК может быть образована из экзонов кодирующих белков генов, таких как муцин и титин . Исследователи предположили, что эккДНК может способствовать экспрессии различных изоформ гена, мешая или способствуя транскрипции специфических экзонов . [ 1 ]
ЭккДНК была классифицирована как одна из четырех различных категорий круговой ДНК на основе размера и последовательности, включая небольшую полидисперсенную круглую ДНК (SPCDNA), теломерные круги (T-цирки), микродНК (100-400 п.н.) и экстрахромосомную ДНК (ECDNA), ECDNA), микродНК (100-400 п.н. и экстрахромосомная ДНК (ECDNA), ECDNA), микродНК (100-400 п.н. и экстрахромосомная ДНК (ECDNA), микродНК (100-400 п.н. Полем [ 27 ] Каждый из этих типов имеет свои уникальные биологические характеристики (см. Таблицу 2): [ 27 ]
| Название экцдны | Размер | Характеристики | Функция |
|---|---|---|---|
| spcdna | 100–10 КБ | Высоко разнообразный тип эккДНК, существует большой диапазон количества SpcDNA, обнаруженных клетки | Участвует в генетической нестабильности человека |
| Теломерные круги | кратные 738 п.н. | Образуются с помощью теломерных массивов, которые представляют собой серию повторяющихся последовательностей в конце линейной ДНК. | Участвует в альтернативном удлинении теломер (ALT) |
| Микродна | 100-400 п.н. | Получен из геномных мест, которые имеют высокое содержание GC и экзонов плотность | Экспрессируют небольшие функциональные регуляторные РНК (например, микроРНК и новые РНК, подобные IS). |
| экдна | 1-3 МБ | Включите полные гены, без теломер , пентрик | Усиление генов, участвующих в развитии рака и лекарственной устойчивости |
Биогенез эккДНК
[ редактировать ]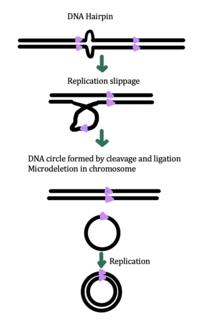


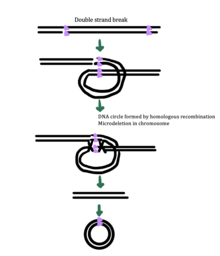
Хотя точный механизм генерации эккДНК до сих пор неизвестен, некоторые исследования показали, что генерация эккДНК может быть связана с восстановлением повреждения ДНК, [ 33 ] гипер-транскрипция, [ 33 ] [ 34 ] гомологичная рекомбинация , [ 35 ] и репликация стресса . [ 33 ] Существует несколько предложенных механизмов для образования экцДНК: (1) проскальзывание репликации создает петлю на цепи шаблона , которая затем иссекается и лигируется в круг, оставляя микроделеуцию на хромосоме , (2) проскальзывание репликации создает петлю в цепи продукта, что это целесообразно. Исправляется и лигируется в круг, который не генерирует , ( 3 механизм образования эккДНК oder ) микроделеуцию в хромосоме Круг и хромосома страдают микроделеоном [ 1 ]
Исследования, проведенные в 2021 году, показали, что апоптотические клетки являются источником ECCDNAS; Это было завершено из -за исследования, показывающего, что апоптотическая фрагментация ДНК (ADF) является предпосылкой для образования экцДНК с помощью методов очистки. [ 31 ]
ЭккДНК может быть получена в результате образования микро-нуклеев, что указывает на хромосомную нестабильность . Было предложено, что преждевременный апоптоз и/или ошибки в хромосомной сегрегации во время митоза могут привести к образованию микро-нуклеев. [ 36 ]
эккДНК в нездоровых клетках
[ редактировать ]Чтобы проверить, используются ли ECCDNA в нелоковых клетках, эмбриональные стволовые клетки мыши и анализ южно-блоттинга использовались ; Результаты подтвердили, что экцДНК обнаруживается как в раковых, так и в незвуковых клетках. [ 31 ] Также известно, что эккДНК вряд ли будет получена из специфических областей генома; Данные секвенирования от 2021 года сообщают, что данные предполагают, что ECCDNAS широко распространены на протяжении всего генома . [ 31 ] Картирование генома полноразмерных ECCDNAs продемонстрировало их различные паттерны выравнивания геном, которые включают в соседние, перекрывающиеся или вложенные положения на одной и той же хромосоме или в разных хромосомах . [ 31 ] ECCDNAs происходят в основном из одиночных, непрерывных геномных локусов, что означает, что один отдельный геномный фрагмент самостоятельно используется для формирования эккДНК, а не образуется из лигирования различных геномных фрагментов. [ 31 ] Эти два варианта могут быть классифицированы как непрерывные и непрерывные ECCDNA, соответственно. [ 31 ] причину циркулялизации фрагментированной ДНК, лигазы три различных фермента были протестированы млекопитающих: LIG1 , LIG3 и Чтобы еще больше понять LIG4 [ 31 ] Полем Используя нокаутные модели в клеточной линии мыши B12F3 B-лимфоцитов , исследования, проведенные в 2021 году, идентифицировали LIG3 в качестве основной лигазы для генерации экцдо в этих клетках. [ 31 ]
Функция
[ редактировать ]Точная функция ECCDNA была обсуждена, но некоторые исследования показали, что ECCDNA могут способствовать усилению генов при раке , [ 1 ] иммунная функция , [ 31 ] и старение . [ 34 ] [ 35 ] [ 37 ]
Функция экцДНК в иммунной системе
[ редактировать ]Согласно исследованию, проведенному в 2021 году, другой функцией ECCDNAS является их роль в качестве возможных иммуностимуляторов . [ 31 ] ЭккДНК значительно индуцирует интерфероны типа I (IFNα, IFNβ), интерлейкин-6 (IL-6) и фактор некроза опухоли (TNF), даже в большей степени, чем линейная ДНК и другие, как правило, мощные индукторы цитокинов при их высоких уровнях концентрации. [ 31 ] Подобные паттерны наблюдаются с макрофагами, поскольку данные показали, что ECCDNAs являются очень мощными иммуностимулянтами в активации как костном мозге, полученных в дендритных клеток, полученных из костного мозга так и макрофагов, . [ 31 ] Кроме того, эксперименты изменили структуру экцДНК с одним ник -ником на сегмент эккДНК и впоследствии обрабатывались ферментами для генерации линейных версий экцДНК. [ 31 ] В этих экспериментах было показано, цитокинов что транскрипция , важный маркер для активности иммунной системы , намного выше в необработанной экцДНК по сравнению с линеаризованной обработкой, придавая, что круговая структура эккДНК, а не сама генетическая последовательность, дает его эккДНК иммунная функция. [ 31 ]
Функция экцДНК при раке
[ редактировать ]Некоторые известные функции эккДНК включают вклад в межклеточную генетическую гетерогенность в опухолях , а также, в частности, онкогенов и усиление лекарственных генов . Это также подтверждает, что гены на экцДНК экспрессируются. В целом, эккДНК была связана с раком и лекарственной устойчивостью , старением , компенсацией генов, [ 1 ] И по этой причине это продолжает оставаться важной темой для обсуждения.
Приложения
[ редактировать ]Роль в раке
[ редактировать ]Подтип эккДНК, такой как экдо, рибосомной ДНК локус ( экстрахромосомный круг рДНК ), и двойные минуты были связаны с геномной нестабильностью . Двойные минуты ECDNAs представляют собой фрагменты экстрахромосомной ДНК , которые первоначально наблюдались в большом количестве опухолей человека , включая молочную грудь, легкие, яичники, толстую кишку и, в частности, нейробластому . Они являются проявлением амплификации генов во время развития опухолей, которые дают клеткам селективные преимущества роста и выживания. Двойные минуты, как и фактические , состоят из хроматина и повторяются в ядре клетки хромосомы во время деления клеток . В отличие от типичных хромосом, они состоят из круглых фрагментов ДНК , размером только несколько миллионов пар оснований и не содержат центромеры или теломер .
Было показано, что двойные хромосомы (DMS), которые присутствуют в виде парных хроматиновых тел при световой микроскопии , являются подмножеством экдо. [ 28 ] [ 38 ] Двойные минутные хромосомы составляют около 30% рак-содержащего спектра экдо, включая отдельные тела, [ 28 ] и было обнаружено, что он содержит идентичное содержание генов как отдельные тела. Обозначение экДНК охватывает все формы большой гены-содержащей экстрахромосомной ДНК, обнаруженной в раковых клетках . Этот тип экДНК обычно наблюдается в раковых клетках различных гистологий , но практически никогда не в нормальной ткани. [ 39 ] [ 28 ] Считается, что ECDNA продуцируется с помощью двухцепочечных разрывов в хромосомах или чрезмерной репликации ДНК в организме. [ 40 ]
Круглая форма экдо эксдо отличается от линейной структуры хромосомной ДНК значимыми способами, которые влияют на патогенез рака . [ 41 ] [ 30 ] Онкогены, кодируемые на экду, имеют огромный транскрипционный выход, ранжируя в 1% генов во всем транскриптоме . В отличие от бактериальных плазмид или митохондриальной ДНК , эСДНК хроматинизируются, содержащие высокие уровни активных марок гистонов , но недостаток репрессивных марок гистонов. В архитектуре экдна хроматина отсутствует уплотнение более высокого порядка, которое присутствует на хромосомной ДНК и является одной из наиболее доступной ДНК во всем геноме рака.
Из ECCDNA было обнаружено, что области прикрепления матрицы (MARS) активируют амплификацию онкогенов . [ 1 ] Трансфекция этих MARS в эмбриональные клетки почек 293T человека привела к увеличению экспрессии генов , что позволяет предположить, что эти MARS, полученные из ECCDNA, участвуют в активации онкогена. [ 42 ] ЭккДНК также, по-видимому, играет роль в других видах рака, таких как рак молочной железы , где онкогены в рецепторе эпидермального фактора роста человека (HER2), позитивные гены рака молочной железы в экцдо, амплифицируются. [ 1 ] Эта эккДНК также показала способность приобретать устойчивость к терапии рецепторных тирозинкиназ (RTK), как HER26. [ 43 ]
Роль в старении
[ редактировать ]Дрожжи являются модельными организмами для изучения старения , а ECCDNAs накапливаются в старых клетках и играют роль в выборе старения у дрожжей. [ 37 ] Продолжается спекуляция на общности этой концепции у более высоких видов, таких как млекопитающие . [ 37 ]
Смотрите также
[ редактировать ]- Экстрахромосомная ДНК
- Экстрахромосомный круг рДНК
- Двойная минута
- Микродна
- Эгоистичные генетические элементы
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час Полсен Т., Кумар П., Косеоглу М.М., Датта А (апрель 2018 г.). «Открытия экстрахромосомных кругов ДНК в нормальных и опухолевых клетках» . Тенденции в генетике . 34 (4): 270–278. doi : 10.1016/j.tig.2017.12.010 . PMC 5881399 . PMID 29329720 .
- ^ Jump up to: а беременный в Gaubatz JW (1990). «Экстрахромосомные круглые DNA и пластичность геномной последовательности в эукариотических клетках». Мутационные исследования . 237 (5–6): 271–292. doi : 10.1016/0921-8734 (90) 90009-G . PMID 2079966 .
- ^ Cohen S, Yacobi K, Segal D (июнь 2003 г.). «Экстрахромосомная круговая ДНК тандемно повторяющихся геномных последовательностей у дрозофилы» . Исследование генома . 13 (6A): 1133–1145. doi : 10.1101/gr.907603 . PMC 403641 . PMID 12799349 .
- ^ Jump up to: а беременный Cohen S, Agmon N, Sobol O, Segal D (март 2010 г.). «Экстрахромосомные круги спутниковых повторений и рибосомной ДНК 5S в клетках человека» . Мобильная ДНК . 1 (1): 11. doi : 10.1186/1759-8753-1-11 . PMC 3225859 . PMID 20226008 .
- ^ Стэнфилд С., Хелински Д.Р. (октябрь 1976 г.). «Маленькая круговая ДНК в Drosophila melanogaster». Клетка . 9 (2): 333–345. doi : 10.1016/0092-8674 (76) 90123-9 . PMID 824055 . S2CID 39382051 .
- ^ Jump up to: а беременный в Shibata Y, Kumar P, Layer R, Willcox S, Gagan Jr, Griffith JD, Dutta A (апрель 2012 г.). «Экстрахромосомные микродны и хромосомные микроделеи в нормальных тканях» . Наука . 336 (6077): 82–86. Bibcode : 2012sci ... 336 ... 82 с . doi : 10.1126/science.1213307 . PMC 3703515 . PMID 22403181 .
- ^ Møller HD, Parsons L, Jørgensen TS, Botstein D, Regenberg B (июнь 2015 г.). «Экстрахромосомная круговая ДНК распространена у дрожжей» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (24): E3114 - E3122. BIBCODE : 2015PNAS..112E3114M . doi : 10.1073/pnas.1508825112 . PMC 4475933 . PMID 26038577 .
- ^ Jump up to: а беременный Shoura MJ, Gabdank I, Hansen L, Merker J, Gotlib J, Levene SD, Fire AZ (октябрь 2017 г.). «Сложные и специфичные для клеток популяции эндогенной круглой ДНК (экцДНК) у Caenorhabditis elegans и Homo Sapiens » . G3 . 7 (10): 3295–3303. doi : 10.1534/g3.117.300141 . PMC 5633380 . PMID 28801508 .
- ^ Hayday AC, Saito H, Gillies SD, Kranz DM, Tanigawa G, Eisen HN, Tonegawa S (февраль 1985 г.). «Структура, организация и соматическая перестройка гамма -генов Т -клеток». Клетка . 40 (2): 259–269. doi : 10.1016/0092-8674 (85) 90140-0 . PMID 3917858 . S2CID 34582929 .
- ^ Jump up to: а беременный в дюймовый Hotta Y, Bassel A (февраль 1965 г.). «Молекулярный размер и кругость ДНК в клетках млекопитающих и более высоких растений» . Труды Национальной академии наук Соединенных Штатов Америки . 53 (2): 356–362. Bibcode : 1965pnas ... 53..356H . doi : 10.1073/pnas.53.2.356 . PMC 219520 . PMID 14294069 .
- ^ Jump up to: а беременный Cox D, Yuncken C, Spriggs AI (июль 1965 г.). «Миловые хроматинские тела в злокачественных опухолях детства». Лансет . 1 (7402): 55–58. doi : 10.1016/s0140-6736 (65) 90131-5 . PMID 14304929 .
- ^ Billheimer FE, Avers CJ (октябрь 1969 г.). «Ядерная и митохондриальная ДНК из дикого типа и миниатюрных дрожжей: круглости, длина и плавучивая плотность» . Труды Национальной академии наук Соединенных Штатов Америки . 64 (2): 739–746. Bibcode : 1969pnas ... 64..739b . doi : 10.1073/pnas.64.2.739 . PMC 223406 . PMID 5261045 .
- ^ Beverley SM, Coderre JA, Santi DV, Schimke RT (сентябрь 1984 г.). «Нестабильные амплификацию ДНК у резистентных к метотрексату лейшмании состоят из экстрахромосомных кругов, которые ревоказуются во время стабилизации». Клетка . 38 (2): 431–439. doi : 10.1016/0092-8674 (84) 90498-7 . PMID 6467372 . S2CID 2030494 .
- ^ Nass MM, Ben-Shaul Y (июнь 1972 г.). «Новая закрытая круговая дуплексная ДНК в обесцвеченных мутантах и зеленых штаммах Euglena gracilis». Biochimica et Biophysica Acta (BBA) - нуклеиновые кислоты и синтез белка . 272 (1): 130–136. doi : 10.1016/0005-2787 (72) 90041-x . PMID 4625469 .
- ^ Вонг Фа, Wildman SG (январь 1972 г.). «Простая процедура выделения спутниковой ДНК из листьев табака с высоким уровнем урожая и демонстрации мини -округ». Biochimica et Biophysica Acta (BBA) - нуклеиновые кислоты и синтез белка . 259 (1): 5–12. doi : 10.1016/0005-2787 (72) 90468-6 . PMID 5011974 .
- ^ Агтериббе Э., Крун А.М., Ван Бругген Эф (май 1972). «Круглая ДНК из митохондрий нейроспоры Crassa» Biochimica et Biophysica Acta (BB) - нуклеиновые кислоты и протесты 269 (2): 299–3 Doi : 10.1016/0005-2787 (72) 90439- x 4260513PMID
- ^ Шталь У, Лемке П.А., Тудзинский П., Кюк У, Эссек К (июль 1978 г.). «Свидетельство о плазмиде, подобной ДНК, в нити, а ascomycete podospora anserina». Молекулярная и общая генетика . 162 (3): 341–343. doi : 10.1007/bf00268860 . PMID 683172 . S2CID 126554440 .
- ^ Lazarus CM, Earl AJ, Turner G, Küntzel H (май 1980). «Амплификация митохондриальной последовательности ДНК в цитоплазмически наследственном« рваном »мутанте aspergillus amstelodami» . Европейский журнал биохимии . 106 (2): 633–641. doi : 10.1111/j.1432-1033.1980.tb04611.x . PMID 6249580 .
- ^ Смит С.А., Виноград Дж (август 1972 г.). Полем Jogoral 69 (2): 163–178. doi : 10.1016/0022-2836 (72) 022-7-7 PMID 5070865 .
- ^ Buongiorno-Nardelli M, Amaldi F, Lava-Sanchez PA (март 1976 г.). «Анализ электронного микроскопа амплифицирующей рибосомной ДНК от Xenopus laevis». Экспериментальные исследования клеток . 98 (1): 95–103. doi : 10.1016/0014-4827 (76) 90467-5 . PMID 1253845 .
- ^ Delap RJ, Rush Mg (декабрь 1978 г.). «Изменение количества и распределения по размерам небольших круговых ДНК во время развития куриной бурсы» . Труды Национальной академии наук Соединенных Штатов Америки . 75 (12): 5855–5859. Bibcode : 1978pnas ... 75.5855d . doi : 10.1073/pnas.75.12.5855 . PMC 393074 . PMID 282606 .
- ^ Тода М., Хирама Т., Такешита С., Ямагиши Х (июнь 1989 г.). «Продукты эксцизионных перестроек гена иммуноглобулина». Иммунологические письма . 21 (4): 311–316. doi : 10.1016/0165-2478 (89) 90025-4 . PMID 2767726 .
- ^ Calabretta B, Robberson DL, Barrera-Saldaña HA, Lambrou TP, Saunders GF (март 1982 г.). «Нестабильность генома в области человеческой ДНК, обогащенной повторными последовательностями ALU». Природа . 296 (5854): 219–225. Bibcode : 1982natur.296..219c . doi : 10.1038/296219A0 . PMID 6278320 . S2CID 4265874 .
- ^ Jump up to: а беременный Yamagishi H, Kunisada T, Iwakura Y, Nishimune Y, Ogiso Y, Matsushiro A (декабрь 1983 г.). «Появление экстрахромосомных круговых ДНК-комплексов в качестве одного из самых ранних сигналов клеточной дифференцировки в раннем развитии эмбриона мыши (эмбрион мыши/тератокарцинома/слюна-пресс-адсорбция/круглое комплекс ДНК/перестройка ДНК)» . Развитие, рост и дифференциация . 25 (6): 563–569. doi : 10.1111/j.1440-169x.1983.00563.x . PMID 37282129 . S2CID 83666742 .
- ^ Цуда Т., Ямагиши Х, Охниши Н., Ямада Й, Изуми Х, Мори К.Дж. (ноябрь 1983 г.). «Экстрахромосомные круглые ДНК из мышиных гемопоэтических тканевых клеток» Плазмида 10 (3): 235–2 Doi : 10.1016/0147-619x (83) 90037-9 6657775PMID
- ^ Jump up to: а беременный Flores SC, Sunnerhagen P, Moore TK, Gaubatz JW (май 1988). «Характеристика повторяющихся семейств последовательностей у мышиного сердца малые полидисперсные циркулярные DNA: возрастные исследования» . Исследование нуклеиновых кислот . 16 (9): 3889–3906. doi : 10.1093/nar/16.9.3889 . PMC 336563 . PMID 3375074 .
- ^ Jump up to: а беременный в дюймовый и Ван Т., Чжан Х, Чжоу Й, Ши Дж (июнь 2021 г.). «Экстрахромосомная круговая ДНК: новая потенциальная роль в прогрессировании рака» . Журнал трансляционной медицины . 19 (1): 257. doi : 10.1186/s12967-021-02927-x . PMC 8194206 . PMID 34112178 .
- ^ Jump up to: а беременный в дюймовый и фон Тернер К.М., Дешпанде В., Бейтер Д., Кога Т., Русерт Дж., Ли С. и др. (Март 2017). «Экстрахромосомная амплификация онкогена стимулирует эволюцию опухоли и генетическую гетерогенность» . Природа . 543 (7643): 122–125. Bibcode : 2017natur.543..122t . doi : 10.1038/nature21356 . PMC 5334176 . PMID 28178237 .
- ^ Møller HD, Mohiyuddin M, Prada-Luengo I, Sailani MR, Halling JF, Plomgaard P, et al. (Март 2018 г.). «Круглые ДНК -элементы хромосомного происхождения распространены в здоровой соматической ткани человека» . Природная связь . 9 (1): 1069. Bibcode : 2018natco ... 9.1069M . doi : 10.1038/s41467-018-03369-8 . PMC 5852086 . PMID 29540679 .
- ^ Jump up to: а беременный Wu S, Turner KM, Nguyen N, Raviram R, Erb M, Santini J, et al. (Ноябрь 2019). «Круглая экдо способствует доступному хроматину и высокой экспрессии онкогена» . Природа . 575 (7784): 699–703. Bibcode : 2019natur.575..699W . doi : 10.1038/s41586-019-1763-5 . PMC 7094777 . PMID 31748743 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Ван Ю., Ван М., Декидель М.Н., Чен Х, Лю Д., Альт Ф.В., Чжан Ю (ноябрь 2021 г.). «ECCDNAs - это апоптотические продукты с высокой врожденной иммуностимулирующей активностью» . Природа . 599 (7884): 308–314. Bibcode : 2021natur.599..308W . doi : 10.1038/s41586-021-04009-w . PMC 9295135 . PMID 34671165 . S2CID 239051756 .
- ^ Jump up to: а беременный Barreto SC, Uppalapati M, Ray A (май 2014). «Маленькие круглые ДНК в человеческой патологии» . Малайзийский журнал медицинских наук . 21 (3): 4–18. PMC 4163554 . PMID 25246831 .
- ^ Jump up to: а беременный в Dillon LW, Kumar P, Shibata Y, Wang YH, Willcox S, Griffith JD, et al. (Июнь 2015 г.). «Производство экстрахромосомных микродней связана с путями восстановления несоответствия и транскрипционной активностью» . Сотовые отчеты . 11 (11): 1749–1759. doi : 10.1016/j.celrep.2015.05.020 . PMC 4481157 . PMID 26051933 .
- ^ Jump up to: а беременный Халл Р.М., Кинг М., Пицца Г., Крюгер Ф., Вергара Х, Хаули Дж (декабрь 2019 г.). «Транскрипционное индуцированное образование экстрахромосомной ДНК при старении дрожжей» . PLOS Биология . 17 (12): E3000471. doi : 10.1371/journal.pbio.3000471 . PMC 6890164 . PMID 31794573 .
- ^ Jump up to: а беременный Грешам Д., Усайт Р., Германн С.М., Лисби М., Ботштейн Д., Регенберг Б. (октябрь 2010 г.). «Адаптация к разнообразным азотным средам путем делеции или экстрахромосомного образования локуса GAP1» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (43): 18551–18556. BIBCODE : 2010PNAS..10718551G . doi : 10.1073/pnas.1014023107 . PMC 2972935 . PMID 20937885 .
- ^ Altungöz; Юксель (сентябрь 2023 г.). «Амплификации генов и экстрахромосомные круглые DNA: функция и биогенез» . Молекулярная биология отчетов . 50 (9): 7693–7703. doi : 10.1007/s11033-023-08649-1 . PMID 37433908 - Via Springer.
- ^ Jump up to: а беременный в Синклер Д.А., Гуаренте Л (декабрь 1997 г.). «Экстрахромосомные круги рДНК-причина старения в дрожжах» . Клетка . 91 (7): 1033–1042. doi : 10.1016/s0092-8674 (00) 80493-6 . PMID 9428525 . S2CID 12735979 .
- ^ Decarvalho AC, Kim H, Poisson LM, Winn ME, Mueller C, Cherba D, et al. (Май 2018). «Разобточное наследство хромосомных и экстрахромосомных ДНК -элементов способствует динамическому эволюции заболеваний при глиобластоме» . Природа генетика . 50 (5): 708–717. doi : 10.1038/s41588-018-0105-0 . PMC 5934307 . PMID 29686388 .
- ^ Ким Х., Нгуен Н.П., Тернер К., Ву С., Гуджар А.Д., Лубек Дж. И др. (Сентябрь 2020 г.). «Экстрахромосомная ДНК связана с амплификацией онкогена и плохим результатом по нескольким видам рака» . Природа генетика . 52 (9): 891–897. doi : 10.1038/s41588-020-0678-2 . PMC 7484012 . PMID 32807987 .
- ^ Kuttler F, Mai S (февраль 2007 г.). «Образование нелупиточных экстрахромосомных элементов во время развития, дифференцировки и онкогенеза». Семинары по биологии рака . 17 (1): 56–64. doi : 10.1016/j.semcancer.2006.10.007 . PMID 17116402 .
- ^ Циммер С (20 ноября 2019 г.). «Ученые только начинают понимать таинственные круги ДНК, распространенные в раковых клетках» . Нью -Йорк Таймс .
- ^ Jin Y, Liu Z, Cao W, Ma X, Fan Y, Yu Y, et al. (2012). «Новые функциональные элементы MAR двойных хромосом в клетках яичников человека, способные повысить экспрессию генов» . Plos один . 7 (2): E30419. BIBCODE : 2012PLOSO ... 730419J . doi : 10.1371/journal.pone.0030419 . PMC 3272018 . PMID 22319568 .
- ^ Викарий Р., Пег В., Моранчо Б., Закарияас-Флюк М., Чжан Дж., Мартинес-Барриоканал А. и др. (2015-06-15). «Паттерны усиления гена HER2 и реакция на терапию против HER2» . Plos один . 10 (6): E0129876. Bibcode : 2015proso..1029876V . Doi : 10.1371/journal.pone.0129876 . PMC 4467984 . PMID 26075403 .
Дальнейшее чтение
[ редактировать ]- Синклер Д.А., Гуаренте Л (декабрь 1997 г.). «Экстрахромосомные круги рДНК-причина старения в дрожжах» . Клетка . 91 (7): 1033–1042. doi : 10.1016/s0092-8674 (00) 80493-6 . PMID 9428525 .
- Cohen S, Houben A, Segal D (март 2008 г.). «Экстрахромосомная круговая ДНК, полученная из тандемно повторяющихся геномных последовательностей в растениях» . Заводский журнал . 53 (6): 1027–1034. doi : 10.1111/j.1365-313x.2007.03394.x . PMID 18088310 .
- Cohen S, Regev A, Lavi S (февраль 1997 г.). «Маленькая полидисперсная круговая ДНК (SPCDNA) в клетках человека: связь с геномной нестабильностью» . Онкоген . 14 (8): 977–985. doi : 10.1038/sj.onc.1200917 . PMID 9050997 .
- Кумар П., Диллон Л.В., Шибата Ю., Джазаери А.А., Джонс Д.Р., Датта А (сентябрь 2017 г.). «Нормальные и раковые ткани высвобождают экстрахромосомную круглую ДНК (эккДНК) в кровообращение» . Молекулярное исследование рака . 15 (9): 1197–1205. doi : 10.1158/1541-7786.mcr-17-0095 . PMC 5581709 . PMID 28550083 .
- Baskin F, Rosenberg RN, Dev V (июнь 1981 г.). «Корреляция двойной минуты хромосом с нестабильной междолевой перекрестной резистентностью у поглощающих мутантов клеток нейробластомы» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (6): 3654–3658. Bibcode : 1981pnas ... 78.3654b . doi : 10.1073/pnas.78.6.3654 . PMC 319629 . PMID 6943568 . Бесплатный полный текст.
- Barker PE (февраль 1982 г.). «Двойные минуты в опухолевых клетках человека». Генетика рака и цитогенетика . 5 (1): 81–94. doi : 10.1016/0165-4608 (82) 90043-7 . PMID 6175392 .
- Masters J, Keeley B, Gay H, Attardi G (май 1982). «Изменное содержание двойных хромосом не коррелирует со степенью нестабильности фенотипа в метоксат-резистентных клеточных линиях человеческих клеток» . Молекулярная и клеточная биология . 2 (5): 498–507. doi : 10.1128/mcb.2.5.498 . PMC 369819 . PMID 7110138 . Бесплатный полный текст.
- Yüksel A, Altungöz O (сентябрь 2023 г.). «Амплификации генов и экстрахромосомные круглые DNA: функция и биогенез». MOL BIOLOGY Reports . 50 (9): 7693–7703. doi : 10.1007/s11033-023-08649-1 . PMID 37433908 .