Смилодон
| Смилодон Временной диапазон: от раннего плейстоцена до раннего голоцена ,
| |
|---|---|

| |
| Установленный популяции S. скелет в Музее науки Теллуса | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Плотоядный |
| Подотряд: | Фелиформия |
| Семья: | кошачьих |
| Подсемейство: | † Махародонтины |
| Племя: | † Смилодонтини |
| Род: | † Смилодон Лунд , 1842 г. |
| Типовой вид | |
| † Популяция смилодона Лунд , 1842 г.
| |
| Другие виды | |
| Синонимы | |
|
Синонимия рода Синонимия видов | |
Смилодон вымерший род кошачьих — . Это один из самых известных саблезубых хищников и доисторических млекопитающих. Хотя он широко известен как саблезубый тигр , он не был тесно связан с тигром или другими современными кошками, принадлежащими к вымершему подсемейству Machairodontinae , с предполагаемой датой расхождения с предком ныне живущих кошек около 20 миллионов лет назад. Смилодон был одним из последних выживших махайродонтов наряду с дальним родственником Homotherium . Смилодон жил в Америке в эпоху плейстоцена (2,5 млн лет назад – 10 000 лет назад). Род был назван в 1842 году на основе окаменелостей из Бразилии; родовое название означает «скальпель» или «обоюдоострый нож» в сочетании с «зубом». три вида Сегодня известны : S. gracilis , S. Fatalis и S. populator . Два последних вида, вероятно, произошли от S. gracilis , который сам, вероятно, произошел от Megantereon . Сотни экземпляров, добытых из смоляных ям Ла Бреа в Лос-Анджелесе, составляют самую большую коллекцию Окаменелости смилодона .
В целом смилодон имел более крепкое телосложение, чем любая из ныне живущих кошек, с особенно хорошо развитыми передними конечностями и исключительно длинными верхними клыками . Его челюсть была шире, чем у современных кошек, а верхние клыки были тонкими и хрупкими и приспособлены для точного убийства. S. gracilis был самым маленьким видом, его вес составлял от 55 до 100 кг (от 121 до 220 фунтов). S. Fatalis имел вес от 160 до 280 кг (от 350 до 620 фунтов) и рост 100 см (39 дюймов). Оба эти вида известны в основном из Северной Америки , но останки из Южной Америки к ним относят и . S. populator из Южной Америки был самым крупным видом, весом от 220 до 436 кг (от 485 до 961 фунта) и ростом 120 см (47 дюймов), и был одним из крупнейших известных кошачьих. Рисунок шерсти смилодона художественно неизвестен, но он был восстановлен с использованием однотонных или пятнистых узоров.
В Северной Америке смилодон охотился на крупных травоядных животных, таких как бизоны и верблюды , и оставался успешным даже при встрече с новыми видами добычи в Южной Америке. Считается, что смилодон убил свою жертву, удерживая ее передними конечностями и кусая, но неясно, каким образом был нанесен сам укус. Ученые спорят, вел ли смилодон социальный или одиночный образ жизни; Анализ поведения современных хищников, а также смилодона ископаемых останков может быть истолкован как подтверждающий любую точку зрения. Смилодон , вероятно, жил в закрытых местах обитания, таких как леса и кустарники , которые служили укрытием для засады на добычу. Смилодон вымер в результате вымирания в конце плейстоцена около 13-10 000 лет назад вместе с большинством других крупных животных по всей Америке. Его зависимость от крупных животных была предложена как причина его исчезновения. S. Fatalis мог подвергнуться воздействию изменения среды обитания и потери добычи, на которой он специализировался, из-за возможных климатических воздействий, последствий недавно прибывшие люди в популяциях-жертвах и другие факторы, в то время как вымирание популяции S. остается плохо изученным.
Таксономия
[ редактировать ]
В 1830-х годах датский натуралист Питер Вильгельм Лунд и его помощники собирали окаменелости в известковых пещерах недалеко от небольшого городка Лагоа-Санта, штат Минас-Жерайс , Бразилия. Среди тысяч найденных окаменелостей он признал несколько изолированных щечных зубов принадлежащими гиене , которую он назвал Hyaena neogaea в 1839 году. После того, как было найдено больше материала (включая клыки и кости стопы), Лунд пришел к выводу, что окаменелости принадлежали гиене. отдельный род кошачьих, хотя и переходный к гиенам. Он заявил, что по размеру он соответствовал бы самым крупным современным хищникам и был более крепким, чем любая современная кошка. Лунд первоначально хотел назвать новый род Hyaenodon , но, понимая, что это имя недавно было применено к другому доисторическому хищнику, он вместо этого назвал его популяцией Smilodon в 1842 году. Он объяснил древнегреческое значение Smilodon как σμίλη ( smilē ), «скальпель» или «скальпель». «нож обоюдоострый» и οδόντος ( odóntos ), «зуб». Это также переводится как «зуб в форме обоюдоострого ножа». Он объяснил название вида populator как «разрушитель», что также переводится как «тот, кто несет опустошение». К 1846 году Лунд приобрел почти все части скелета (от разных людей), а в последующие годы другие коллекционеры нашли еще больше экземпляров в соседних странах. [ 1 ] [ 2 ] Хотя некоторые более поздние авторы использовали оригинальное видовое название Лунда neogaea вместо populator , теперь оно считается недействительным nomen nudum , поскольку не сопровождалось надлежащим описанием и не было обозначено никаких типовых экземпляров. [ 3 ] Некоторые южноамериканские экземпляры были отнесены к другим родам, подродам, видам и подвидам , таким как Smilodontidion riggii , Smilodon ( Prosmilodon ) ensenadensis и S. bonaeriensis , но теперь они считаются младшими синонимами S. populator . [ 4 ]

Окаменелости смилодона были обнаружены в Северной Америке со второй половины XIX века. [ 1 ] В 1869 году американский палеонтолог Джозеф Лейди описал фрагмент верхней челюсти с коренным зубом , который был обнаружен в нефтяном пласте в округе Хардин, штат Техас . Он отнес этот экземпляр к роду Felis (который затем использовался для обозначения большинства кошек, как ныне живущих, так и вымерших), но нашел его достаточно отличным, чтобы быть частью собственного подрода , как F. ( Trcifelis ) Fatalis . [ 5 ] Название вида означает «смертельный». [ 6 ] В статье 1880 года о вымерших американских кошках американский палеонтолог Эдвард Дринкер Коуп отметил, что коренной зуб F. Fatalis идентичен таковому у Smilodon , и предложил новую комбинацию S. Fatalis . [ 7 ] Большинство североамериканских находок были скудными, пока не начались раскопки в смоляных ямах Ла-Бреа в Лос-Анджелесе сотни особей S. Fatalis . , где с 1875 года были обнаружены [ 1 ] У S. Fatalis есть младшие синонимы, такие как S. mercerii , S. floridanus и S.californicus . [ 4 ] Американский палеонтолог Анналиса Берта считала голотип S. Fatalis слишком неполным, чтобы быть адекватным типовым экземпляром, и иногда этот вид предлагался как младший синоним S. populator . [ 3 ] Скандинавские палеонтологи Бьорн Куртен и Ларс Верделин поддержали различие этих двух видов в статье, опубликованной в 1990 году. [ 8 ] В статье американского палеонтолога Джона П. Бабиарца и его коллег, опубликованной в 2018 году, был сделан вывод, что S.californicus , представленный экземплярами из смоляных ям Ла-Бреа, в конце концов был отдельным видом от S. Fatalis , и что необходимы дополнительные исследования для уточнения таксономии. рода. [ 9 ]
В своей статье 1880 года о вымерших кошках Коуп также назвал третий вид смилодонов — S. gracilis . Этот вид был основан на частичной собачьей особи, которая была получена в пещере Порт-Кеннеди недалеко от реки Шуйлкилл в Пенсильвании. Коуп обнаружил, что этот клык отличается от клыка других видов Smilodon из-за его меньшего размера и более сжатого основания. [ 7 ] Его видовое название относится к более легкому телосложению этого вида. [ 10 ] Этот вид известен по меньшему количеству и менее полным останкам, чем другие представители рода. [ 11 ] S. gracilis иногда считался частью таких родов, как Megantereon и Ischyrosmilus . [ 12 ] S. populator , S. Fatalis и S. gracilis в настоящее время считаются единственными действительными видами Smilodon , а признаки, используемые для определения большинства их младших синонимов, были отклонены как вариации между особями одного и того же вида (внутривидовые вариации). [ 4 ] [ 3 ] , одно из самых известных доисторических млекопитающих, Смилодон часто упоминается в популярных средствах массовой информации и является окаменелостью штата Калифорния. [ 1 ]
Эволюция
[ редактировать ]


долгое время являвшийся наиболее известной саблезубой кошкой , Смилодон, по-прежнему остается одним из самых известных представителей группы, до такой степени, что эти два понятия путают. Сам термин « саблезубый » относится к экоморфу, состоящему из различных групп вымерших хищных синапсид (млекопитающих и близких родственников), у которых в ходе конвергентной эволюции развились чрезвычайно длинные верхнечелюстные клыки , а также адаптации черепа и скелета, связанные с их использованием. Сюда входят представители Gorgonopsia , Thylacosmilidae , Machaeroidinae , Nimravidae , Barbourofelidae и Machairodontinae . [ 1 ] [ 13 ] В семействе Felidae (настоящие кошки) члены подсемейства Machairodontinae называются саблезубыми кошками, и сама эта группа делится на три трибы : Metailurini (ложные саблезубые); Homotherini ( ятаганзубые кошки); и Smilodontini ( кортикозубые кошки), к которым относится смилодон . [ 4 ]
Члены Smilodontini отличаются длинными тонкими клыками с мелкими зубцами или без них , тогда как Homotherini характеризуются более короткими, широкими и более уплощенными клыками с более грубыми зубцами. [ 14 ] Представители Metailurini были менее специализированы и имели более короткие и менее уплощенные клыки, и некоторые исследователи не признавали их представителями Machairodontinae. [ 4 ]
Несмотря на разговорное название «саблезубый тигр», смилодон не имеет близкого родства ни с современным тигром (который принадлежит к подсемейству Pantherinae ), ни с каким-либо другим существующим кошачьим. [ 15 ] Анализ древней ДНК 1992 года показал, что смилодона следует отнести к современным кошкам (подсемейства Felinae и Pantherinae). [ 16 ] Исследование 2005 года показало, что смилодон принадлежал к отдельной линии. [ 17 ] Исследование, опубликованное в 2006 году, подтвердило это, показав, что Machairodontinae рано отделились от предков ныне живущих кошек и не были тесно связаны ни с одним из ныне живущих видов. [ 18 ] Предки современных кошек и Machairodontinae, по оценкам, разошлись около 20 миллионов лет назад. [ 19 ] Следующая кладограмма, основанная на окаменелостях и анализе ДНК, показывает место Smilodon среди вымерших и современных кошачьих, по данным Ринкона и его коллег, 2011: [ 20 ]
| кошачьих |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Самые ранние кошачьи известны из олигоцена Европы, такие как Proailurus , а самый ранний с саблезубыми чертами — миоценовый род Pseudaelurus . [ 4 ] Морфология черепа и нижней челюсти самых ранних саблезубых кошек была аналогична таковой у современных дымчатых леопардов ( Neofelis ). Эта линия в дальнейшем адаптировалась к точному убийству крупных животных за счет развития удлиненных клыков и более широких зияний, при этом жертвуя высокой силой укуса . [ 21 ] Поскольку их клыки стали длиннее, тела кошек стали более устойчивыми к обездвиживанию добычи. [ 14 ] У производных смилодонтинов и гомотеринов поясничный отдел позвоночника и хвост укоротились, как и задние конечности. [ 4 ] Махародонты когда-то представляли собой доминирующую группу кошачьих, распространенную в Африке, Евразии и Северной Америке в эпохи миоцена и плиоцена. [ 22 ] но постепенно сокращался в течение плейстоцена, [ 23 ] к позднему плейстоцену осталось только два рода махайродонтов: Smilodon и отдаленно родственный Homotherium , оба в основном обитали в Америке. Судя по митохондриальной ДНК, последовательностям извлеченным из древних костей , линии Homotherium и Smilodon , по оценкам, разошлись около 18 миллионов лет назад. [ 19 ]
Самым ранним видом смилодона является S. gracilis , который существовал от 2,5 миллионов до 500 000 лет назад (от раннего Бланкана до ирвингтонского периода) и был преемником в Северной Америке Мегантереона , от которого он, вероятно, произошел. Сам Мегантереон проник в Северную Америку из Евразии в плиоцене вместе с гомотерием . S. gracilis достиг северных районов Южной Америки в раннем плейстоцене в рамках Великого американского обмена . [ 20 ] [ 14 ] S. Fatalis существовал 1,6–10 000 лет назад (от позднего Ирвингтона до Ранчолабрея ) и заменил S. gracilis в Северной Америке. [ 8 ] Популяция S. существовала 1–10 000 лет назад ( от Энсенадана до Луджана ); это произошло в восточных частях Южной Америки. [ 24 ]
Описание
[ редактировать ]
Скелет
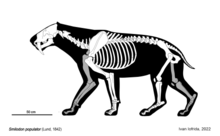
[ редактировать ]Смилодон был размером с современных больших кошек , но имел более крепкое телосложение. У него была уменьшенная поясничная область , высокая лопатка , короткий хвост и широкие конечности с относительно короткими ступнями. [ 25 ] [ 26 ] Смилодон наиболее известен своими относительно длинными клыками, которые являются самыми длинными у саблезубых кошек: у самого крупного вида S.populator их длина составляет около 28 см (11 дюймов) . [ 25 ] Клыки были тонкими и имели мелкие зазубрины на передней и задней стороне. [ 27 ] Череп имел крепкие пропорции, морда короткая и широкая. Скулы ( скулы ) глубокие и широко изогнутые, сагиттальный гребень выражен, лобная область слегка выпуклая. Нижняя челюсть имела выступы с каждой стороны спереди. Верхние резцы были большими, острыми и наклоненными вперед. имеется диастема Между резцами и молярами нижней челюсти (промежуток). Нижние резцы были широкими, загнутыми назад и располагались по прямой линии. p3 Премоляр нижней челюсти присутствовал у большинства ранних экземпляров, но утрачивался у более поздних экземпляров; он присутствовал только в 6% выборки Ла Бреа. [ 3 ] Существует некоторый спор о том, ли Смилодон был сексуально диморфным . Некоторые исследования окаменелостей S. Fatalis не выявили особых различий между полами. [ 28 ] [ 29 ] И наоборот, исследование 2012 года показало, что, хотя окаменелости S. Fatalis демонстрируют меньшие различия в размерах среди особей, чем современные Panthera , они, похоже, демонстрируют те же различия между полами по некоторым признакам. [ 30 ]
S. gracilis был самым маленьким видом, вес которого оценивался от 55 до 100 кг (от 121 до 220 фунтов), что примерно соответствовало размеру ягуара . Он был похож на своего предшественника Megantereon того же размера, но его зубной ряд и череп были более развитыми, приближаясь к S. Fatalis . [ 31 ] [ 4 ] S. Fatalis занимал промежуточное положение по размеру между S. gracilis и S. populator . [ 25 ] Он варьировался от 160 до 280 кг (от 350 до 620 фунтов). [ 31 ] и достиг высоты в плечах 100 см (39 дюймов) и длины тела 175 см (69 дюймов). [ 32 ] Размерами он был похож на льва, но был более крепким и мускулистым, а потому имел большую массу тела. Его череп также был похож на череп Мегантереона , но более массивный и с более крупными клыками. [ 4 ] S. populator был одним из крупнейших известных кошачьих , с массой тела от 220 кг (490 фунтов) до более 400 кг (880 фунтов). [ 31 ] и по одной оценке, до 470 кг (1040 фунтов). [ 33 ] Особенно большой череп популяции S. из Уругвая длиной 39,2 см (15,4 дюйма) указывает на то, что этот человек мог весить до 436 кг (961 фунт). [ 34 ] Его высота в плечах составляла 120 см (47 дюймов). [ 25 ] По сравнению с S. Fatalis , S. populator был более крепким и имел более удлиненный и узкий череп с более прямым верхним профилем, более высоким расположением носовых костей , более вертикальным затылком , более массивными метаподиями и немного более длинными передними конечностями по сравнению с задними конечностями. [ 4 ] [ 8 ] Большие ископаемые следы из Аргентины (для которых ихнотаксона было предложено название Smilodonichium ) были отнесены к S. populator и имеют размеры 17,6 см (6,9 дюйма) на 19,2 см (7,6 дюйма). [ 35 ] Это больше, чем следы бенгальского тигра , с которыми сравнивали следы. [ 36 ]
Внешние особенности
[ редактировать ]

Смилодон и другие саблезубые кошки были реконструированы как с однотонной шерстью, так и с пятнистым рисунком (что, по-видимому, является предковым состоянием кошачьих ), оба из которых считаются возможными. [ 37 ] Исследования современных видов кошек показали, что виды, живущие на открытом воздухе, как правило, имеют однородную шерсть, в то время как те, которые живут в местах обитания с большей растительностью, имеют больше отметин, за некоторыми исключениями. [ 38 ] Некоторые особенности шерсти, такие как гривы львов-самцов или полосы тигра, слишком необычны, чтобы их можно было предсказать по окаменелостям. [ 37 ]
Традиционно саблезубые кошки были художественно восстановлены с внешними чертами, похожими на черты современных кошачьих, такими художниками, как Чарльз Р. Найт, в сотрудничестве с различными палеонтологами в начале 20 века. [ 37 ] В 1969 году палеонтолог Дж. Дж. Миллер вместо этого предположил, что Смилодон сильно отличался от типичной кошки и был похож на бульдога , с линией нижней губы (чтобы позволить его рту широко открываться, не разрывая лицевые ткани), более втянутым носом и более втянутым носом. низко расположенные уши. [ 39 ] Палеохудожник Маурисио Антон и соавторы оспорили это в 1998 году и утверждали, что черты лица смилодона в целом не сильно отличались от черт лица других кошек. Антон отметил, что современные животные, такие как бегемоты, способны чрезвычайно широко открывать рот, не разрывая ткани, благодаря сложенной круговой мышце рта , и такое расположение мышц существует у современных крупных кошачьих. [ 40 ] Антон заявил, что сохранившийся филогенетический брекетинг (когда в качестве эталона используются черты ближайших сохранившихся родственников ископаемого таксона) является наиболее надежным способом восстановления жизненного облика доисторических животных, и поэтому реставрации кошачьих смилодонов , проведенные Найтом, являются все еще точно. [ 37 ] Исследование, проведенное Антоном и его коллегами в 2022 году, пришло к выводу, что верхние клыки Smilodon были бы видны, когда рот был закрыт, а у Homotherium - нет, после изучения окаменелостей и дошедших до нас больших кошек. [ 41 ]
Палеобиология
[ редактировать ]Диета
[ редактировать ]
Высший хищник , смилодон в основном охотился на крупных млекопитающих. Изотопы, сохранившиеся в костях S. Fatalis в смоляных ямах Ла-Бреа, показывают, что жвачных животных, таких как бизон ( Bison antiquus , который был намного крупнее современного американского бизона ) и верблюдов ( Camelops ). кошки чаще всего поедали там [ 42 ] Smilodon Fatalis, возможно, также иногда охотился на Glyptotherium , судя по черепу молоди Glyptotherium texanum, извлеченному из плейстоценовых отложений в Аризоне, которые имеют характерные эллиптические следы от проколов, лучше всего соответствующие меткам Smilodon , что указывает на то, что хищник успешно прокусил череп через глиптодонта. бронированный головной щит. [ 43 ] Кроме того, изотопы, сохранившиеся в зубной эмали экземпляров S. gracilis показывают, что этот вид питался пекарием Platygonus и ламоподобным из Флориды , Hemiauchenia . [ 44 ] Измерения стабильных изотопов углерода остатков S. gracilis во Флориде значительно различались в разных местах и показывают, что этот вид был гибким в своих пищевых привычках. [ 45 ] Изотопные исследования костей лютого волка ( Aenocyon dirus ) и американского льва ( Panthera atrox ) показывают совпадение с S. Fatalis в добыче, что позволяет предположить, что они были конкурентами. [ 42 ] Однако более детальный изотопный анализ показывает, что Smilodon Fatalis предпочитал лесную добычу, такую как тапиры, олени и лесные бизоны, в отличие от предпочтений ужасных волков в отношении добычи, обитающей на открытых территориях, таких как луга. [ 46 ] Доступность добычи в районе Ранчо Ла Бреа , вероятно, была сопоставима с современной Восточной Африкой . [ 47 ]

Когда смилодон мигрировал в Южную Америку, его рацион изменился; бизоны отсутствовали, лошади и хоботные были другими, а местные копытные, такие как токсодонты и литоптерны, были совершенно незнакомы, однако S. populator процветал там так же хорошо, как и его родственники в Северной Америке. [ 14 ] Изотопный анализ популяции S. показывает, что ее основными видами-жертвами были Toxodon Platensis , Pachyarmatherium , Holmesina , виды рода Panochthus , Palaeolama , Catonyx , Equus neogeus и крокодил Caiman latirostris . Этот анализ его рациона также указывает на то, что популяция S. охотилась как на открытых, так и в лесных местообитаниях. [ 48 ] Различия между видами Северной и Южной Америки могут быть связаны с разницей в добыче на двух континентах. [ 8 ] Смилодон , возможно, избегал употребления в пищу костей и оставил бы достаточно еды для падальщиков. [ 49 ] Копролиты, отнесенные к популяции S., обнаруженной в Аргентине, сохраняют Mylodon остеодермы и ладьевидную кость ламы . В дополнение к этому недвусмысленному свидетельству потребления костей, копролиты предполагают, что у смилодона была более универсальная диета, чем считалось ранее. [ 50 ] Исследования стоматологической микроодежды из Ла Бреа также позволяют предположить, что смилодон поедал и плоть, и кости. [ 51 ] Сам Смилодон , возможно, собирал добычу ужасных волков. [ 52 ] Было высказано предположение, что смилодон был чистым падальщиком, который использовал свои клыки для демонстрации своего господства над тушами, но эта теория сегодня не поддерживается, поскольку ни одно современное наземное млекопитающее не является чистым падальщиком. [ 53 ]
Хищническое поведение
[ редактировать ]
Мозг , смилодона сходную с имел структуру бороздок таковой у современных кошек, что предполагает повышенную сложность областей, контролирующих слух, зрение и координацию конечностей. У кошачьих саблезубых в целом были относительно маленькие глаза , не такие обращенные вперед, как у современных кошек, у которых хорошее бинокулярное зрение, позволяющее им передвигаться по деревьям. [ 53 ] Смилодон , вероятно, был хищником, устраивавшим засады и скрывавшимся в густой растительности, поскольку пропорции его конечностей были похожи на современных лесных кошек. [ 54 ] а короткий хвост не помог бы ему сохранять равновесие во время бега. [ 55 ] В отличие от своего предка Мегантереона , который был хотя бы частично сканзором и, следовательно, мог лазить по деревьям, Смилодон , вероятно, был полностью наземным существом из-за своего большего веса и отсутствия приспособлений для лазания. [ 56 ] Треки из Аргентины под названием Felipeda miramarensis в 2019 году, возможно, были спродюсированы Smilodon . Если следы правильно идентифицированы, то следы указывают на то, что у животного были полностью втягивающиеся когти, стопоходящие лапы, отсутствовала сильная способность супинации лап, особенно крепкие передние конечности по сравнению с задними конечностями и, вероятно, он охотился из засады. [ 57 ]
Пяточная кость смилодона была довольно длинной, что позволяет предположить, что он был хорошим прыгуном. [ 25 ] Хорошо развитые мышцы-сгибатели и разгибатели предплечий, вероятно , позволяли ему тянуть вниз и надежно удерживать крупную добычу. Анализ поперечных срезов S. Fatalis Hueri показал, что они были усилены корковым утолщением до такой степени, что могли выдерживать большую нагрузку, чем у современных больших кошек или вымершего американского льва. Кортикальная стенка плечевой кости у S. Fatalis была на 15 % толще, чем ожидалось у современных крупных кошек аналогичного размера. Утолщение S. Fatalis бедренных костей было в пределах современных кошачьих. [ 58 ] [ 59 ] Его клыки были хрупкими и не могли прокусить кость; из-за риска сломаться этим кошкам приходилось подчинять и сдерживать свою добычу своими мощными передними конечностями, прежде чем она могла использовать свои клыки, и, вероятно, использовали быстрые режущие или колющие укусы, а не медленные, удушающие укусы, обычно используемые современными кошками. [ 58 ] В редких случаях, о чем свидетельствуют окаменелости, смилодон был готов рискнуть прокусить кость своими клыками. Возможно, это было больше сосредоточено на конкуренции, такой как другие смилодоны , или на потенциальных угрозах, таких как другие хищники, чем на добыче. [ 56 ]

Продолжаются споры о том, как смилодон убил свою жертву. Традиционно самая популярная теория состоит в том, что кошка нанесла глубокий колющий укус или колющий удар с открытой пастью в горло, очень быстро убивая жертву. [ 58 ] [ 60 ] Другая гипотеза предполагает, что Смилодон нацелился на брюхо своей добычи. Это оспаривается, поскольку искривление живота жертвы, вероятно, помешало бы кошке получить хороший укус или удар. [ 61 ] Что касается того, как смилодон нанес укус, предпочтение отдавалось гипотезе «собачьего сдвига укуса», согласно которой сгибание шеи и вращение черепа помогали укусить добычу, но это может быть механически невозможно. Однако данные сравнения с Homotherium позволяют предположить, что Smilodon был полностью способен использовать собачий сдвиговый укус в качестве основного средства убийства добычи, основываясь на том факте, что у него был толстый череп и относительно небольшая трабекулярная кость, в то время как Homotherium имел и то, и другое. трабекулярная кость и более львиный зажимной укус в качестве основного средства нападения на добычу. Открытие, сделанное Фигейридо и Лаутеншлагером и др. и опубликованное в 2018 году, предполагает совершенно разные экологические адаптации у обоих махайродонтов. [ 62 ] Края нижней челюсти, возможно, помогали противостоять изгибающим силам, когда нижняя челюсть прижималась к шкуре животного-жертвы. [ 63 ] («Робокот») на тушах бизона и лося, экспериментально доказано, С помощью машины, которая воссоздает зубы и имитирует движения челюстей и шеи Smilodon Fatalis что колющий укус в горло является гораздо более правдоподобным. и практическая техника убийства, чем колющий укус в живот. [ 64 ]


Выступающие резцы располагались дугой и использовались для удержания добычи и стабилизации ее во время укуса собаки. Контактная поверхность между коронкой собаки и десной была увеличена, что помогло стабилизировать зуб и помочь кошке почувствовать, когда зуб прорезался в максимальную степень. Поскольку у саблезубых кошек обычно было относительно большое подглазничное отверстие (отверстие) в черепе, в котором располагались нервы, связанные с усами, было высказано предположение, что улучшенные чувства помогли бы кошкам повысить точность при кусании вне поля зрения, и тем самым предотвратить поломку клыков. Похожие на лезвия хищные зубы использовались для разрезания кожи и доступа к мясу, а уменьшенные коренные зубы позволяют предположить, что они были менее приспособлены для дробления костей, чем современные кошки. [ 53 ] Поскольку пища современных кошек при резке попадает в рот сбоку хищными, а не передними резцами между клыками, животным не нужно широко разиваться, поэтому клыки смилодона также не были бы помехой при кормлении. [ 40 ] Опубликованное в 2022 году исследование того, как питались махайродонты, показало, что характер износа зубов S. Fatalis также позволяет предположить, что он способен поедать кости в такой же степени, как и львы. Это, а также сравнения со следами укусов, оставленными современным махайродонтом Xenosmilus, позволяют предположить, что смилодон и его родственники могли эффективно очищать мясную тушу во время кормления, не мешая своим длинным клыкам. [ 65 ]
Несмотря на более мощное телосложение, чем у других крупных кошек, смилодон имел более слабый укус. Современные большие кошки имеют более выраженные скуловые дуги они были меньше , в то время как у смилодонов , что ограничивало толщину и, следовательно, мощность височных мышц образом, уменьшало смилодона и, таким силу укуса . Анализ его узких челюстей показывает, что он мог произвести укус лишь на треть сильнее, чем у льва (коэффициент силы укуса, измеренный для льва, равен 112). [ 66 ] [ 67 ] Кажется, существует общее правило: саблезубые кошки с самыми большими клыками имеют пропорционально более слабый укус. Анализ силы изгиба клыков (способность клыков сопротивляться изгибающим силам, не ломаясь) и силы укуса показывает, что зубы саблезубых кошек были сильнее по сравнению с силой укуса, чем у современных больших кошек. [ 68 ] Кроме того, смилодона зияние могло достигать более 110 градусов. [ 69 ] а у современного льва достигает 65 градусов. [ 70 ] Это сделало зияние достаточно широким, чтобы смилодон мог схватить крупную добычу, несмотря на длинные клыки. [ 40 ] Исследование 2018 года сравнило убийственное поведение Smilodon Fatalis и сыворотки Homotherium и обнаружило, что у первого был крепкий череп с небольшим количеством трабекулярной кости для колющего укуса, тогда как у второго было больше трабекулярной кости, и он чаще использовал стиль зажима и удержания. похожи на львов. Таким образом, эти два вида заняли бы разные экологические ниши. [ 71 ]
Природные ловушки
[ редактировать ]
Многие экземпляры смилодонов были выкопаны из асфальтовых просачиваний , которые служили естественными ловушками для хищников. Животные случайно попали в канализацию и стали приманкой для хищников, пришедших собирать мусор, но затем они сами оказались в ловушке. Самые известные из таких ловушек находятся в Ла-Бреа в Лос-Анджелесе, где было произведено более 166 000 Smilodon Fatalis . экземпляров [ 72 ] которые составляют самую большую коллекцию в мире. Отложения ям накопились 40–10 тысяч лет назад, в позднем плейстоцене . Хотя пойманных животных быстро хоронили, хищникам часто удавалось вырвать у них кости конечностей, но они сами часто попадали в ловушку, а затем добывались другими хищниками; 90% раскопанных костей принадлежали хищникам. [ 73 ]
Смоляные просачивания Талара в Перу представляют собой аналогичный сценарий и также содержат окаменелости смилодона . В отличие от Ла Бреа, многие кости были сломаны или имеют следы выветривания. Возможно, это произошло потому, что слои были более мелкими, поэтому избиение пойманных животных повредило кости ранее пойманных животных. Многие из хищников в Таларе были молодыми, что, возможно, указывает на то, что неопытные и менее приспособленные животные имели больше шансов попасть в ловушку. Хотя Лунд считал, что скопления окаменелостей смилодона и травоядных в пещерах Лагоа-Санта произошли из-за того, что кошки использовали пещеры в качестве логовищ, это, вероятно, результат того, что животные умирали на поверхности, а потоки воды впоследствии утаскивали их кости на пол пещеры. , но некоторые люди могли также умереть, заблудившись в пещерах. [ 73 ]
Социальная жизнь
[ редактировать ]
Ученые спорят, ли смилодон был социальным . Одно исследование африканских хищников показало, что социальные хищники, такие как львы и пятнистые гиены, больше реагируют на сигналы бедствия , чем одиночные виды. Поскольку окаменелости S. Fatalis распространены в смоляных ямах Ла-Бреа и, вероятно, были привлечены криками о помощи застрявшей добычи, это может означать, что этот вид также был социальным. [ 74 ] В одном критическом исследовании утверждается, что в исследовании не учитываются другие факторы, такие как масса тела (более тяжелые животные с большей вероятностью застревают, чем более легкие), интеллект (некоторые социальные животные, такие как американский лев, возможно, избегали смолы, потому что они были лучше способны (чтобы распознать опасность), отсутствие визуальных и обонятельных приманок, тип звуковой приманки и продолжительность сигналов бедствия (фактические сигналы бедствия пойманных животных-жертв длились бы дольше, чем сигналы, использованные в исследовании). Автор этого исследования размышляет о том, как бы отреагировали хищники, если бы записи были проиграны в Индии, где, как известно, в остальном одиночные тигры собираются вокруг одной туши. [ 75 ] Авторы оригинального исследования ответили, что, хотя эффекты звонков в tar-ямах и экспериментов по воспроизведению не будут идентичными, этого будет недостаточно, чтобы опровергнуть их выводы. Кроме того, они заявили, что вес и интеллект вряд ли повлияют на результаты, поскольку более легкие плотоядные животные гораздо более многочисленны, чем тяжелые травоядные, а в ямах также встречается социальный (и, казалось бы, умный) лютоволк. [ 76 ]

Другой аргумент в пользу социальности основан на заживших травмах на нескольких окаменелостях смилодона , что позволяет предположить, что животным нужны были другие люди, чтобы обеспечить их пищей. [ 77 ] [ 78 ] Этот аргумент был подвергнут сомнению, поскольку кошки могут быстро восстановиться даже после серьезного повреждения костей, а раненый смилодон мог бы выжить, если бы у него был доступ к воде. [ 79 ] Однако смилодон , страдающий дисплазией тазобедренного сустава в молодом возрасте и доживший до взрослой жизни, предполагает, что он не мог бы дожить до взрослой жизни без помощи социальной группы, поскольку этот человек не мог охотиться или защищать свою территорию из-за серьезности врожденной проблемы. . [ 80 ] Мозг смилодона был относительно небольшим по сравнению с другими видами кошек. Некоторые исследователи утверждают, что был мозг смилодона слишком мал, чтобы он мог быть социальным животным. [ 81 ] Анализ размера мозга ныне живущих больших кошек не выявил корреляции между размером мозга и социальностью. [ 82 ] Еще один аргумент против социальной принадлежности смилодона заключается в том, что охота из засады в закрытой среде обитания, вероятно, сделала бы ненужной групповую жизнь, как у большинства современных кошек. [ 79 ] Тем не менее, также было высказано предположение, что, будучи крупнейшим хищником в среде, сравнимой с африканской саванной, смилодон, возможно, имел социальную структуру, подобную современным львам, которые, возможно, живут группами, главным образом, для защиты оптимальной территории от других львов (львы — это сегодня только социальные большие кошки). [ 53 ]

Был ли смилодон сексуально диморфным, это имеет значение для его репродуктивного поведения. Основываясь на своих выводах об отсутствии полового диморфизма , Ван Валкенбург и Сакко предположили в 2002 году, что, если бы кошки были социальными, они, вероятно, жили бы моногамными парами (вместе с потомством) без острой конкуренции между самцами за самок. [ 28 ] менее выражена, Аналогичным образом, Мичен-Сэмюэлс и Биндер в 2010 году пришли к выводу, что агрессия между самцами у S. Fatalis чем у американского льва. [ 29 ] Кристиансен и Харрис обнаружили в 2012 году, что, поскольку S. Fatalis действительно проявлял некоторый половой диморфизм, для конкуренции между самцами должен был существовать эволюционный отбор. [ 30 ] Некоторые кости свидетельствуют о том, что их укусил другой смилодон , возможно, в результате территориальных сражений, конкуренции за права размножения или из-за добычи. [ 53 ] На двух черепах популяций S. из Аргентины видны, по-видимому, смертельные незаживающие раны, которые, по-видимому, были причинены клыками другого смилодона (хотя нельзя исключать, что они были вызваны ударом ногой добычи). Если это вызвано внутривидовой борьбой, это также может указывать на то, что их социальное поведение могло привести к смерти, как это наблюдается у некоторых современных кошек (а также на то, что клыки могут проникать в кости). [ 83 ] Было высказано предположение, что увеличенные клыки саблезубых кошек эволюционировали для сексуальной демонстрации и конкуренции, но статистическое исследование корреляции между собаками и размером тела у S. populator не обнаружило различий в масштабировании между размером тела и собак и пришло к выводу, что это больше вероятно, они эволюционировали исключительно ради хищнической функции. [ 84 ]
Набор из трех связанных скелетов S. Fatalis, найденных в Эквадоре и описанных в 2021 году Рейнольдсом, Сеймуром и Эвансом, позволяет предположить, что смилодон находился под длительной родительской опекой . У двух обнаруженных несовершеннолетних особей есть уникальная унаследованная черта зубных костей, что позволяет предположить, что они были братьями и сестрами; редкий случай семейных отношений, обнаруженный в летописи окаменелостей. Предполагается также, что полувзрослые экземпляры были мужского и женского пола соответственно, в то время как останки взрослых особей, найденные на этом месте, предположительно принадлежали их матери. По оценкам, на момент смерти подросткам было около двух лет, но они все еще росли. [ 85 ] S. Fatalis имел пропорционально большие подъязычные кости , чем современные виды кошачьих, и поэтому, вероятно, издавал более глубокие вокалы. Хотя смилодон имел такое же количество подъязычных костей, как и « ревущие » кошки, их форма была ближе к форме « мурлыкающих » видов. [ 86 ]
Разработка
[ редактировать ]
У смилодона начали развиваться саблезубы, когда животное достигло возраста от 12 до 19 месяцев, вскоре после завершения прорезывания молочных зубов у кошки . И детские, и взрослые клыки присутствовали во рту бок о бок в течение примерно 11 месяцев, а мышцы, используемые для мощного укуса, также были развиты примерно в полтора года, восемью месяцами ранее. чем у современного льва. После того, как смилодон достиг возраста 23–30 месяцев, молочные зубы выпадали, в то время как взрослые клыки росли со средней скоростью 7 мм (0,28 дюйма) в месяц в течение 12-месячного периода. Они достигли своего полного размера примерно в 3-летнем возрасте, позже, чем современные виды больших кошек. Молодые и подростки смилодонов крайне редки на Ранчо Ла Бреа, где проводилось исследование, что указывает на то, что они оставались скрытыми или в местах убежищ во время охоты и зависели от заботы родителей, пока их клыки развивались. [ 87 ] [ 88 ] [ 89 ]
Исследование 2024 года обнаружило доказательства того, что подростки -смилодоны хранили свои молочные сабли в течение длительного периода времени (по оценкам, 30 месяцев), чтобы помочь укрепить своих взрослых клыков по мере их роста. В результате молочные сабли действовали как структурная поддержка, позволяя им начать охоту. с минимальным риском для их зрелого набора сабель. В результате сохранение сабли из кошачьего молока уменьшило изгибающую нагрузку на прорезывающиеся взрослые зубы кошки, когда она прикусывала их, поскольку было обнаружено, что прорезывающиеся сабли были гораздо более уязвимы для поломки по мере роста, чем во время взросления. Это также привело бы к тому, что на этой стадии роста у смилодона были «двойные клыки», что подтверждается обнаружением особей на этой онтогенной стадии на Ранчо Ла Бреа. [ 90 ] [ 91 ]
Исследование 2017 года показывает, что подростки рождаются с крепким телосложением, похожим на взрослых. Сравнение костей молодых экземпляров S. Fatalis из Ла-Бреа с костями современного американского льва показало, что обе кошки имеют схожую кривую роста. Развитие передних конечностей кошачьих в онтогенезе (изменения во время роста) остается жестко ограниченным. Кривая аналогична кривой современных кошек, таких как тигры и пумы, но больше смещается в сторону более сильного направления осей, чем это наблюдается у современных кошачьих. [ 92 ] Исследования Рейнольдса, Сеймура и Эванса (2021) показывают, что смилодон имел уникальную и быструю скорость роста, аналогичную тигру, но что у этого рода был длительный период роста, подобный тому, что наблюдается у львов, и что детеныши зависели от своих родителей, пока этот период роста не закончился. [ 85 ]
Палеопатология
[ редактировать ]
На нескольких окаменелостях смилодона наблюдаются признаки болезни Бехтерева , гиперостоза и травмы. [ 93 ] Одно исследование 1000 черепов смилодонов показало, что у 36% из них были разрушены теменные кости , где прикрепляются самые большие мышцы челюсти. У них также были обнаружены признаки микропереломов, а также ослабление и истончение костей, возможно, вызванное механическим воздействием из-за постоянной необходимости совершать колющие движения клыками. [ 94 ] Костные наросты там, где дельтовидная мышца прикрепляется к плечевой кости, являются распространенной патологией для особей из Ла Бреа, которая, вероятно, возникла из-за неоднократного напряжения, когда смилодон пытался стянуть добычу передними конечностями. Травмы грудины также распространены, вероятно, из-за столкновения с добычей. [ 95 ]
Частота травм у экземпляров S. Fatalis составила 4,3% по сравнению с 2,8% у лютого волка, что означает, что хищническое поведение первого из засад привело к большему риску травм, чем хищническое поведение последнего. На останках смилодона плечевых и поясничных позвонков . обнаружено относительно больше травм [ 96 ] Исследование 2023 года зафиксировало высокую степень субхондральных дефектов на поверхности конечностей и суставов S. Fatalis и особей лютого волка из ям Ла-Бреа-Тар, которые напоминали рассекающий остеохондроз . Поскольку современные собаки с этим заболеванием являются инбредными , исследователи предположили, что то же самое могло произойти с доисторическими видами, когда они были на грани исчезновения, но предупредили, что необходимы дополнительные исследования, чтобы определить, имело ли это место и с образцами из других частей Земли. Америки. [ 97 ]
Остеомиелит левой четвертой пястной кости был зарегистрирован у образца популяции S., относящегося к 5-й стадии морского изотопа . Эта патология привела к тому, что особь махайродонта стала неспособной сгибать пальцы ног и серьезно снизила бы ее способность охотиться на добычу. [ 98 ]
Распространение и среда обитания
[ редактировать ]
Смилодон жил в эпоху плейстоцена (2,5 млн лет назад – 10 000 лет назад) и был, пожалуй, самым последним из саблезубых кошек. [ 25 ] S. Fatalis обитал в самых разных средах обитания, будучи способен населять открытые луга и парки. [ 99 ] окраинные лесисто-луговые массивы, [ 100 ] и закрытые леса. [ 101 ] Окаменелости этого рода были обнаружены по всей Америке. [ 3 ] Самыми северными остатками этого рода являются окаменелости S. Fatalis из Альберты, Канада . [ 102 ] самые южные останки известны с крайнего юга Патагонии. [ 103 ] Ареал обитания Северной Америки варьировался от субтропических лесов и саванн на юге до безлесных мамонтовых степей на севере. Мозаичная растительность лесов, кустарников и трав на юго-западе Северной Америки поддерживала крупных травоядных животных, таких как лошади, бизоны, антилопы , олени , верблюды, мамонты , мастодонты и наземные ленивцы . Северная Америка также поддерживала других саблезубых кошек, таких как Homotherium и Xenosmilus , а также других крупных хищников, включая ужасных волков, короткомордого медведя ( Arctodus simus ) и американского льва. [ 14 ] [ 73 ] [ 104 ] Конкуренция со стороны таких хищников, возможно, помешала североамериканскому S. Fatalis достичь размера популяции S. Fatalis в Южной Америке . Сходство размеров S. Fatalis и американского льва предполагает перекрытие ниш и прямую конкуренцию между этими видами, и, по-видимому, они питались добычей одинакового размера. [ 105 ]

S. gracilis проник в Южную Америку в период от раннего до среднего плейстоцена, где, вероятно, дал начало S. populator , обитавшему в восточной части континента. S. Fatalis также проник в западную часть Южной Америки в позднем плейстоцене, и считалось, что эти два вида разделены горами Анд . [ 8 ] [ 20 ] [ 25 ] Однако в 2018 году было сообщено о черепе S. Fatalis, найденном в Уругвае к востоку от Анд, что ставит под сомнение идею о том, что эти два вида были аллопатрическими (географически разделенными). [ 106 ] Американский обмен привел к тому, что смешались местные и инвазивные виды в прериях и лесах Южной Америки ; Травоядные животные Северной Америки включали хоботных, лошадей, верблюдовых и оленей, травоядные Южной Америки включали токсодонтов, литоптерн, наземных ленивцев и глиптодонтов. Местные метатерийские хищники (в том числе саблезубые тилакосмилиды ) вымерли в плиоцене и были заменены североамериканскими хищниками, такими как псовые, медведи и крупные кошки. [ 14 ]
S. populator оказался очень успешным, тогда как Homotherium так и не получил широкого распространения в Южной Америке. Вымирание тилакосмилид приписывают конкуренции со смилодонами , но это, вероятно, неверно, поскольку они, по-видимому, исчезли до прибытия крупных кошек. Форусрацидные «птицы-террористы» , возможно, доминировали в нише крупных хищников в Южной Америке до прибытия смилодона . [ 14 ] S. populator , возможно, смог достичь большего размера, чем S. Fatalis, из-за отсутствия конкуренции в плейстоценовой Южной Америке; S. populator появился после исчезновения Arctotherium angustidens , одного из крупнейших хищников за всю историю, и поэтому смог занять нишу мегахищника. [ 105 ] Популятор S. предпочитал крупную добычу из открытых мест обитания, таких как луга и равнины, основываясь на данных, полученных на основе соотношений изотопов, определяющих рацион животного. В этом отношении южноамериканский вид Smilodon , вероятно, был похож на современного льва. S. populator, вероятно, конкурировал здесь с псовым Protocyon , но не с ягуаром, который питался преимущественно более мелкой добычей. [ 107 ] [ 108 ] С другой стороны, морфометрия указывает на то, что S. populator лучше всего приспособлен к более закрытой среде. [ 109 ]
Вымирание
[ редактировать ]
Наряду с большей частью мегафауны плейстоцена Нового Света, смилодон вымер 10 000 лет назад на этапах вымирания позднего плейстоцена в Северной и Южной Америке . Его исчезновение было связано с упадком и исчезновением крупных травоядных животных. Следовательно, смилодон мог быть слишком специализирован на охоте на крупную добычу и не смог адаптироваться. [ 58 ] Действительно, во время потепления Бёллинга-Аллерёда и перед в раннем дриасе похолоданием у S. Fatalis наблюдались изменения в морфологии черепа, которые намекают на усиление специализации на более крупной добыче и/или эволюции в ответ на конкуренцию с другими хищниками. [ 110 ] Однако исследование стираемости зубов смилодонов, проведенное в 2012 году , не выявило доказательств того, что они были ограничены пищевыми ресурсами. [ 111 ] Другие объяснения включают изменение климата и конкуренцию с Homo Sapiens. [ 111 ] (которые проникли в Америку примерно в то время, когда исчез смилодон ), или сочетание нескольких факторов, каждый из которых относится к общему вымиранию в позднем плейстоцене , а не конкретно к вымиранию саблезубых кошек. [ 112 ] Одним из часто упоминаемых здесь факторов является похолодание в Младшем дриасе , которое, возможно, резко сократило обитаемое пространство для многих видов. Что касается влияния человека, то есть свидетельства смены режима, вызванной пожаром, на Ранчо ла Бреа, которая предшествовала истреблению мегафауны в этом районе, причем люди, скорее всего, несут ответственность за увеличение интенсивности пожаров. [ 113 ]
Писатели первой половины двадцатого века предположили, что последние саблезубые кошки, Smilodon и Homotherium , вымерли в результате конкуренции с пришедшими им на смену более быстрыми и генерализованными кошачьими. Было даже высказано предположение, что саблезубые хищники уступают современным кошкам, поскольку считалось, что постоянно растущие клыки мешают их владельцам правильно питаться. Однако с тех пор было показано, что рацион махайродонтинов, таких как Smilodon и Homotherium, был разнообразным. Судя по всему, они не ограничивались добычей гигантских животных, как предполагалось ранее, а питались всем, что было доступно, включая крупного рогатого скота , лошадей и верблюдов . [ 114 ] [ 115 ] Кроме того, немахайродонтные кошачьи, такие как американский лев и мирациноникс, также вымерли в позднем плейстоцене, а саблезубые и коническизубые кошачьи раньше сосуществовали более миллиона лет. [ 116 ] Тот факт, что саблезубые зубы эволюционировали много раз в несвязанных линиях, также свидетельствует об успехе этой особенности. [ 112 ]
Самая молодая прямая радиоуглеродная дата S. Fatalis отличается от датировки S. populatus на тысячи лет: первая - незадолго до похолодания в раннем дриасе, а вторая - на начало голоцена. [ 117 ] Последний экземпляр S. Fatalis , обнаруженный в смоляных ямах Ранчо Ла Бреа, датирован 13 025 лет назад. [ 118 ] Образец S. Fatalis из Айовы датируется 13 605–13 455 годами ранее (BP). [ 119 ] Последние останки популяции смилодонов, найденные в пещере Куэва-дель-Медио, недалеко от города Сория, на северо-востоке провинции Эльтима-Эсперанса , регион Магальянес на юге Чили , датируются 10 935–11 209 годами назад. [ 120 ] Самая последняя достоверная дата появления S. Fatalis по углероду-14 составляет 11 130 лет назад. [ 121 ] Однако такие радиоуглеродные даты, скорее всего, не откалиброваны, а это означает, что они не были скорректированы с календарных лет на обычные годы. В результате даты кажутся моложе, чем они есть на самом деле. Таким образом, экземпляр S. Fatalis с Ранчо Ла Бреа является самым молодым из зарегистрированных видов. [ 117 ] предполагая вымирание до Младшего дриаса на основании его последнего появления в Калифорнии, в отличие от других регионов, где мегафауна сократилась из-за Младшего дриаса. [ 113 ]
См. также
[ редактировать ]- Список крупнейших хищников
- Список крупнейших доисторических хищников
- Мегафауна
- Позднеплейстоценовые вымирания
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Антон 2013 , с. 3–26.
- ^ Лунд, PW (1842 г.). Посмотрите на животный мир Бразилии в поисках последнего земного переворота (на датском языке). Копенгаген: Естествознание и математические тезисы Датского королевского общества наук. стр. 54–57.
- ^ Jump up to: а б с д и Берта, А. (1985). «Статус смилодона в Северной и Южной Америке» (PDF) . Вклад в науку, Музей естественной истории округа Лос-Анджелес . 370 : 1–15. Архивировано из оригинала (PDF) 28 мая 2014 года.
- ^ Jump up to: а б с д и ж г час я дж Антон 2013 , с. 108–154.
- ^ Лейди, Дж. (1869). «Вымершая фауна млекопитающих Дакоты и Небраски: включая отчет о некоторых родственных формах из других мест, а также краткий обзор останков млекопитающих Северной Америки» . Журнал Академии естественных наук Филадельфии . 7 : 366–367. дои : 10.5962/bhl.title.20910 .
- ^ «Саблезубый» . Нэшнл Географик. 21 сентября 2011 года . Проверено 13 октября 2021 г.
- ^ Jump up to: а б Коуп, ЭД (декабрь 1880 г.). «О вымерших кошках Америки» . Американский натуралист . 14 (12): 833–858. дои : 10.1086/272672 . JSTOR 2449549 .
- ^ Jump up to: а б с д и Куртен, Б.; Верделин, Л. (1990). «Отношения между североамериканским и южноамериканским смилодоном ». Журнал палеонтологии позвоночных . 10 (2): 158–169. Бибкод : 1990JVPal..10..158K . дои : 10.1080/02724634.1990.10011804 . JSTOR 4523312 .
- ^ Верделин, Л.; Макдональд, Х.Г.; Шоу, Калифорния (2018). Смилодон : культовый саблезубый ( смилодон из Южной Каролины: значение для таксономии рода под ред.). Балтимор: Издательство Университета Джонса Хопкинса. стр. 76–95. ISBN 978-1-4214-2557-3 .
- ^ Куртен, Б.; Андерсон, Э. (1980). Плейстоценовые млекопитающие Северной Америки . Нью-Йорк: Издательство Колумбийского университета. стр. 186–188. ISBN 978-0-231-03733-4 .
- ^ Берта, А. (1987). «Саблезубый кот Smilodon gracilis из Флориды и обсуждение его родственных связей (Mammalia, Felidae, Smilodontini)» . Бюллетень Государственного музея Флориды . 31 : 1–63.
- ^ Черчер, CS (1984). «Статус Smilodontopsis (Brown, 1908) и Ischyrosmilus (Merriam, 1918): таксономический обзор двух родов саблезубых кошек (Felidae, Machairodontinae)» . Взносы Королевского музея Онтарио в области наук о жизни . 140 : 14–34 . дои : 10.5962/bhl.title.52222 . ISBN 978-0-88854-305-9 .
- ^ Михан, Т.; Мартин, LDJ (2003). «Вымирание и реэволюция подобных адаптивных типов (экоморфов) у кайнозойских копытных и плотоядных животных Северной Америки отражают циклы ван дер Хаммена». Die Naturwissenschaften . 90 (3): 131–135. Бибкод : 2003NW.....90..131M . дои : 10.1007/s00114-002-0392-1 . ПМИД 12649755 . S2CID 21117744 .
- ^ Jump up to: а б с д и ж г Антон 2013 , с. 65–76.
- ^ «Что такое саблезубый?» . Музей палеонтологии Калифорнийского университета. Декабрь 2005 года . Проверено 12 июня 2012 г.
- ^ Янчевский, Д.Н.; Юки, Н.; Гилберт, округ Колумбия; Джефферсон, GT; О'Брайен, SJ (1992). «Молекулярно-филогенетический вывод на основе окаменелостей саблезубых кошек с Ранчо Ла Бреа» . Труды Национальной академии наук . 89 (20): 9769–9773. Бибкод : 1992PNAS...89.9769J . дои : 10.1073/pnas.89.20.9769 . ПМЦ 50214 . ПМИД 1409696 .
- ^ Барнетт, Р.; Барнс, И.; Филлипс, MJ; Мартин, LD; Харингтон, Чехия; Леонард, Дж.А.; Купер, А. (2005). «Эволюция вымерших саблезубых и американской гепардообразной кошки» . Современная биология . 15 (15): 589–590 рэндов. Бибкод : 2005CBio...15.R589B . дои : 10.1016/j.cub.2005.07.052 . ПМИД 16085477 . S2CID 17665121 .
- ^ ван ден Хук Остенде, левый фланговый игрок; Морло, М.; Нагель, Д. (2006). «Величественные убийцы: саблезубые кошки (Ископаемые объяснили 52)». Геология сегодня . 22 (4): 150–157. дои : 10.1111/j.1365-2451.2006.00572.x . S2CID 128960196 .
- ^ Jump up to: а б Пайманс, JLA; Барнетт, Р.; Гилберт, MTP; Сепеда-Мендоса, ML; Реймер, JWF; де Вос, Дж.; Зазула, Г.; Нагель, Д.; Барышников Г.Ф.; Леонард, Дж.А.; Роланд, Н.; Вестбери, Миннесота; Барлоу, А.; Хофрейтер, М. (19 октября 2017 г.). «Эволюционная история саблезубых кошек на основе древней митогеномики» . Современная биология . 27 (21): 3330–3336.e5. Бибкод : 2017CBio...27E3330P . дои : 10.1016/j.cub.2017.09.033 . ПМИД 29056454 .
- ^ Jump up to: а б с Ринкон, А.; Превости, Ф.; Парра, Г. (2011). «Новые записи саблезубых кошек (Felidae: Machairodontinae) для плейстоцена Венесуэлы и Великого американского биотического обмена». Журнал палеонтологии позвоночных . 31 (2): 468–478. Бибкод : 2011JVPal..31..468R . дои : 10.1080/02724634.2011.550366 . hdl : 11336/69016 . JSTOR 25835839 . S2CID 129693331 .
- ^ Кристиансен, П. (2008). «Эволюция формы черепа и нижней челюсти у кошек (Carnivora: Felidae)» . ПЛОС ОДИН . 3 (7): e2807. Бибкод : 2008PLoSO...3.2807C . дои : 10.1371/journal.pone.0002807 . ПМЦ 2475670 . ПМИД 18665225 .

- ^ Кристиансен, Пер (октябрь 2010 г.). «Филогения саблезубых кошачьих (Carnivora: Felidae: Machairodontinae)» . Кладистика . 29 (5): 543–559. дои : 10.1111/cla.12008 . ISSN 0748-3007 .
- ^ Пирас, Паоло; Сильвестро, Даниэле; Каротенуто, Франческо; Кастильоне, Сильвия; Коцакис, Анастасиос; Майорино, Леонардо; Мельчионна, Марина; Монданаро, Алессандро; Сансалоне, Габриэле; Серьезно, Кармела; Верно, Вероника Анна; Райя, Паскуале (май 2018 г.). «Эволюция нижней челюсти саблезубого зуба: смертельная экоморфологическая специализация» . Палеогеография, Палеоклиматология, Палеоэкология . 496 : 166–174. дои : 10.1016/j.palaeo.2018.01.034 . hdl : 2158/1268434 .
- ^ де Кастро, Мариэла Кордейро; Лангер, Макс Кардозо (2008). «Новые посткраниальные останки популяции Smilodon Lund, 1842 г., из южно-центральной Бразилии» . Бразильский журнал палеонтологии . 11 (3): 199–206. дои : 10.4072/rbp.2008.3.06 .
- ^ Jump up to: а б с д и ж г Тернер, А.; Антон, М. (1997). Большие кошки и их ископаемые родственники: иллюстрированный путеводитель по их эволюции и естественной истории . Издательство Колумбийского университета. стр. 57–58, 67–68. ISBN 978-0-231-10229-2 . OCLC 34283113 .
- ^ «Что такое саблезубый?» . Музей палеонтологии Калифорнийского университета . Проверено 08 апреля 2013 г.
- ^ Слейтер, Дж.Дж.; Валкенбург, Б.В. (2008). «Длинный в зубе: эволюция формы черепа саблезубой кошки». Палеобиология . 34 (3): 403–419. Бибкод : 2008Pbio...34..403S . дои : 10.1666/07061.1 . ISSN 0094-8373 . S2CID 85353590 .
- ^ Jump up to: а б Ван Валкенбург, Б.; Сакко, Т. (2002). «Половой диморфизм, социальное поведение и внутриполовая конкуренция у крупных хищников плейстоцена». Журнал палеонтологии позвоночных . 22 (1): 164–169. doi : 10.1671/0272-4634(2002)022[0164:sdsbai]2.0.co;2 . JSTOR 4524203 . S2CID 86156959 .
- ^ Jump up to: а б Мичен-Сэмюэлс, Дж.; Биндер, В. (2010). «Половой диморфизм и онтогенетический рост американского льва и саблезубой кошки с ранчо Ла Бреа». Журнал зоологии . 280 (3): 271–279. дои : 10.1111/j.1469-7998.2009.00659.x .
- ^ Jump up to: а б Кристиансен, Пер; Харрис, Джон М. (2012). «Изменения в краниомандибулярной морфологии и половом диморфизме у пантерин и саблезубого кота Smilodon Fatalis» . ПЛОС ОДИН . 7 (10): е48352. Бибкод : 2012PLoSO...748352C . дои : 10.1371/journal.pone.0048352 . ПМЦ 3482211 . ПМИД 23110232 .
- ^ Jump up to: а б с Кристиансен, Пер; Харрис, Джон М. (2005). «Размер тела смилодона (Mammalia: Felidae)». Журнал морфологии . 266 (3): 369–84. дои : 10.1002/jmor.10384 . ПМИД 16235255 . S2CID 27233870 .
- ^ «Саблезубый кот, Smilodon Fatalis » . Глобальный зоопарк Сан-Диего. Январь 2009 г. Архивировано из оригинала 3 февраля 2013 г. Проверено 7 мая 2013 г.
- ^ Соркин, Б. (2008). «Биомеханическое ограничение массы тела у наземных хищников-млекопитающих». Летайя . 41 (4): 333–347. Бибкод : 2008Лета..41..333С . дои : 10.1111/j.1502-3931.2007.00091.x .
- ^ Манцуэтти, А.; Переа, Д.; Джонс, В.; Убилла, М.; Риндеркнехт, А. (2020). «Чрезвычайно большой череп саблезубой кошки из Уругвая (поздний плейстоцен – ранний голоцен, формация Долорес): размер тела и палеобиологические значения». Алчеринга: Австралазийский журнал палеонтологии . 44 (2): 332–339. Бибкод : 2020Алч...44..332М . дои : 10.1080/03115518.2019.1701080 . S2CID 216505747 .
- ^ «Неопубликованная находка в Мирамаре: окаменелые следы большого саблезубого тигра» . 0223.com.ar (на испанском языке). 0223. 26 мая 2016 . Проверено 28 мая 2016 г.
- ^ Перкинс, Сид (10 июня 2016 г.). «Первые ископаемые следы саблезубых кошек больше, чем лапы бенгальского тигра» . Наука . дои : 10.1126/science.aag0602 .
- ^ Jump up to: а б с д Антон 2013 , с. 157–176.
- ^ Аллен, WL; Катхилл, IC; Скотт-Сэмюэл, штат Невада; Бэддели, Р. (2010). «Почему у леопарда появились пятна: связь развития узора с экологией кошачьих» . Труды Королевского общества Б. 278 (1710): 1373–1380. дои : 10.1098/rspb.2010.1734 . ПМК 3061134 . ПМИД 20961899 .
- ^ Миллер, Дж. Дж. (1969). «Новая гипотеза, объясняющая метод приема пищи, используемый Smilodon Californicus Bovard». Тебива . 12 :9–19.
- ^ Jump up to: а б с Антон, М.; Гарсиа-Переа, Р.; Тернер, А. (1998). «Реконструированный облик лица саблезубого кошачьего смилодона ». Зоологический журнал Линнеевского общества . 124 (4): 369–386. дои : 10.1111/j.1096-3642.1998.tb00582.x .
- ^ Антон, Морис; Кремнезем, драгоценный камень; Пастор Джон Ф.; Салеса, Мануэль Дж. (2022). «Скрытое оружие: пересмотренная реконструкция анатомии лица и жизненного облика саблезубой кошки Homotherium latidens (Felidae, Machairodontinae)» . Четвертичные научные обзоры . 284 : 107471. Бибкод : 2022QSRv..28407471A . doi : 10.1016/j.quascirev.2022.107471 . hdl : 10261/270770 . S2CID 248168629 .
- ^ Jump up to: а б Колтрейн, Дж. Б.; Харрис, Дж. М.; Серлинг, TE; Элерингер-младший; Диринг, доктор медицины; Уорд, Дж.; Аллен, Дж. (2004). «Биогеохимия стабильных изотопов Ранчо Ла Бреа и ее значение для палеоэкологии позднего плейстоцена, прибрежной южной Калифорнии» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 205 (3–4): 199–219. Бибкод : 2004PPP...205..199C . дои : 10.1016/j.palaeo.2003.12.008 . Архивировано из оригинала (PDF) 11 ноября 2011 г.
- ^ Антон 2013 , стр. 203–204.
- ^ Феранец, Р.С. (2005). «Скорость роста и продолжительность роста взрослых собак S. gracilis и выводы о диете на основе анализа стабильных изотопов» (PDF) . Бюллетень Музея естественной истории Флориды . 45 (4): 369–377. doi : 10.58782/flmnh.psyo5090 – через Университет Флориды.
- ^ Феранец, Роберт С.; ДеСантис, Лариса Р.Г. (лето 2014 г.). «Понимание особенностей универсального рациона хищных животных путем анализа значений стабильных изотопов углерода у плейстоценовых млекопитающих Флориды» . Палеобиология . 40 (3): 477–493. Бибкод : 2014Pbio...40..477F . дои : 10.1666/13055 . ISSN 0094-8373 . Проверено 21 января 2024 г. - через Cambridge Core.
- ^ ДеСантис, Лариса Р.Г.; Критс, Джонатан М.; Феранец, Роберт С.; Фокс-Доббс, Кена; Фаррелл, Эслинг Б.; Харрис, Джон М.; Такеучи, Гэри Т.; Серлинг, Туре Э. (2019). «Причины и последствия вымирания плейстоценовой мегафауны, выявленные на млекопитающих ранчо Ла Бреа» . Современная биология . 29 (15): 2488–2495. Бибкод : 2019CBio...29E2488D . дои : 10.1016/j.cub.2019.06.059 . PMID 31386836 .
- ^ Ванвалкенбург, Б.; Хертель, Ф. (1993). «Трудные времена в Ла-Бреа: поломка зубов у крупных хищников позднего плейстоцена». Наука . 261 (5120): 456–459. Бибкод : 1993Sci...261..456V . дои : 10.1126/science.261.5120.456 . ПМИД 17770024 . S2CID 39657617 .
- ^ Дантас, Марио Андре Триндаде; Черкинский, Александр; Лесса, Карлос Микаэль Бонфим; САНТОС, Лучано Вилабойм; Коссуоль, Марио Альберто; ОМЕНА, Эрика Кавальканте; СИЛЬВА, Хорхе Луис Лопес; Сиал, Альсидес Нобрега; Бочеренс, Эрве (14 июля 2020 г.). «Изотопная пеоэкология (δ13C, δ18O) сообщества позвоночных позднего плейстоцена из межтропического региона Бразилии» . Бразильский журнал палеонтологии . 23 (2): 138–152. дои : 10.4072/rbp.2020.2.05 . ISSN 2236-1715 .
- ^ Ван Валкенбург, Б.; Тифорд, МФ; Уокер, А. (1990). «Молярная микроодежда и диета крупных хищников: выводы относительно диеты саблезубой кошки Smilodon Fatalis ». Журнал зоологии . 222 (2): 319–340. дои : 10.1111/j.1469-7998.1990.tb05680.x .
- ^ Морено Родригес, Ханна П.; Цемент, Николас Р.; Агнолин, Фредерик Л.; Джоффри, Уильям; Джентиле, Адриэль (2022). «Возможный копролит Smilodon (Mammalia, Felidae) из плейстоцена Аргентины» . ПАЛЕОС 37 (7): 402–410. Бибкод : 2022Представитель..37..402М . дои : 10.2110/figure.2021.056 . ISSN 1938-5323 . S2CID 251078622 .
- ^ ДеСантис, Лариса Р.Г.; Шоу, Кристофер А. (2018). «Саблезубые кошки с зубной болью: влияние травм зубов на пищевое поведение Smilodon Fatalis (Mammalia, Felidae) позднего плейстоцена с Ранчо ла Бреа (Лос-Анджелес, Калифорния)» . Геологическое общество Америки Рефераты с программами . 50 (6): 322567. doi : 10.1130/abs/2018AM-322567 .
- ^ Ван Валкенбург, Б. (1991). «Итеративная эволюция гиперплотоядности у псовых (Mammalia: Carnivora): эволюционные взаимодействия между симпатрическими хищниками». Палеобиология . 17 (4): 340–362. Бибкод : 1991Pbio...17..340В . дои : 10.1017/S0094837300010691 . JSTOR 2400749 . S2CID 251052044 .
- ^ Jump up to: а б с д и Антон 2013 , с. 176–216.
- ^ Гония, WJ (1976). «Поведенческие последствия морфологии саблезубых кошачьих». Палеобиология . 2 (4): 332–342. Бибкод : 1976Pbio....2..332G . дои : 10.1017/S0094837300004966 . JSTOR 2400172 . S2CID 87481727 .
- ^ «Что такое саблезубый?» . Беркли.edu . Проверено 1 июня 2017 г.
- ^ Jump up to: а б Антон, Маурисио (2013). Саблезубый .
- ^ Агнолин, Флорида; Цемент, Северная Каролина; Филд, ДХ; Магнуссен, М.; Бох, Д.; Де Чианни, Ф. (2019). «Большие следы хищников из позднего плейстоцена Аргентины» Ихнос 26 (2): 119–126. Бибкод : 2019Ично..26..119А дои : 10.1080/10420940.2018.1479962 . S2CID 134190731 .
- ^ Jump up to: а б с д Мичен-Сэмюэлс, Дж.А.; Ван Валкенбург, Б. (2010). «Рентгенограммы показывают исключительную силу передних конечностей у саблезубого кота Smilodon Fatalis » . ПЛОС ОДИН . 5 (7): е11412. Бибкод : 2010PLoSO...511412M . дои : 10.1371/journal.pone.0011412 . ISSN 1932-6203 . ПМК 2896400 . ПМИД 20625398 .
- ^ Кришнасвами, Долли Дж. (2 июля 2010 г.). «ScienceShot: Саблезубые тигры добавляют в свой арсенал мощное оружие» . Наука .
- ^ МакГенри, ЧР; Роу, С.; Клаузен, PD; Морено, К.; Каннингем, Э. (2007). «Супермоделированная саблезубая кошка, хищническое поведение Smilodon Fatalis, выявленное с помощью 3D-компьютерного моделирования высокого разрешения» . ПНАС . 104 (41): 16010–16015. Бибкод : 2007PNAS..10416010M . дои : 10.1073/pnas.0706086104 . ПМК 2042153 . ПМИД 17911253 .
- ^ Эньонг, В. (1996). «Микроизнос на клыках и убийственное поведение крупных хищников: функция сабли у Smilodon Fatalis » . Журнал маммологии . 77 (4): 1059–1067. дои : 10.2307/1382786 . JSTOR 1382786 .
- ^ Фигейридо, Б.; Лаутеншлагер, С.; Перес-Рамос, А.; Ван Валкенбург, Б. (2018). «Отличительное хищническое поведение ятаганных и кинжалозубых саблезубых кошек» . Современная биология . 28 (20): 3260–3266.e3. Бибкод : 2018CBio...28E3260F . дои : 10.1016/j.cub.2018.08.012 . hdl : 10630/29727 . ПМИД 30293717 .
- ^ Браун, Дж. Г. (2014). «Функция челюсти у Smilodon Fatalis : переоценка сдвигового прикуса собак и предложение новой модели рычага класса 1 с приводом от передних конечностей» . ПЛОС ОДИН . 9 (10): е107456. Бибкод : 2014PLoSO...9j7456B . дои : 10.1371/journal.pone.0107456 . ISSN 1932-6203 . ПМЦ 4182664 . ПМИД 25272032 .
- ^ Уиллер, Х. Тодд (2011). «Экспериментальная палеонтология смертельных укусов ятагана и кинжала» . В Л. Неаполе, Вирджиния; Д. Мартин, Ларри; П. Бабиарц, Джон (ред.). Другие саблезубые: ятаганзубые кошки западного полушария . Издательство Университета Джонса Хопкинса. стр. 19–35.
- ^ Домингес-Родриго, Мануэль; Эгеланд, Чарльз П.; Кобо-Санчес, Люсия; Бакедано, Энрике; Халберт, Ричард К. (2022). «Поведение в потреблении туш саблезубых особей и динамика крупных гильдий хищников плейстоцена» . Научные отчеты . 12 (1): 6045. Бибкод : 2022NatSR..12.6045D . дои : 10.1038/s41598-022-09480-7 . ПМК 9061710 . ПМИД 35501323 .
- ^ Хехт, Дж. (1 октября 2007 г.). «Саблезубый кот обладал удивительно тонким укусом» . Новый учёный .
- ^ Роу, С.; МакГенри2, К.; Томасон, Дж. (2004). «Клуб укусов: сравнительная сила укуса крупных кусающих млекопитающих и прогноз хищнического поведения ископаемых таксонов» (PDF) . Труды Королевского общества . 272 (1563): 619–25. дои : 10.1098/рспб.2004.2986 . ПМК 1564077 . ПМИД 15817436 . Архивировано из оригинала (PDF) 25 августа 2013 г.
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Кристиансен, П. (2007). «Сравнительная сила укуса и сила изгиба собак у кошачьих и саблезубых кошачьих: значение для экологии хищников» . Зоологический журнал Линнеевского общества . 151 (2): 423–437. дои : 10.1111/j.1096-3642.2007.00321.x .
- ^ Андерссон, К.; Норман, Д.; Верделин, Л. (2011). «Саблезубые хищники и убийство крупной добычи» . ПЛОС ОДИН . 6 (10): e24971. Бибкод : 2011PLoSO...624971A . дои : 10.1371/journal.pone.0024971 . ПМК 3198467 . ПМИД 22039403 .
- ^ Мартин, LD (1980). «Функциональная морфология и эволюция кошек» . Труды Академии наук Небраски . 8 : 141–154.
- ^ Фигейридо, Б.; Лаутеншлагер, С.; Перес-Рамос, А.; Ван Валкенбург, Б. (2018). «Отличительное хищническое поведение ятаганных и кинжалозубых саблезубых кошек» . Современная биология . 28 (20): 3260–3266.e3. Бибкод : 2018CBio...28E3260F . дои : 10.1016/j.cub.2018.08.012 . hdl : 10630/29727 . ПМИД 30293717 .
- ^ Шоу, Кристофер А.; Куинн, Джеймс П. (15 сентября 2015 г.). Джон М. Харрис (ред.). «Добавление Smilodon Fatalis (Mammalia: Carnivora: Felidae) к биоте позднеплейстоценовых асфальтовых отложений Carpinteria в Калифорнии с онтогенетическими и экологическими последствиями для этого вида» (PDF) . Научная серия 42: Вклад в науку (специальный том под названием «Ла Бреа и за его пределами: палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия раскопок Музея естественной истории округа Лос-Анджелес на ранчо Ла Бреа). Музей естественной истории округа Лос-Анджелес: 91–95. Архивировано из оригинала (PDF) 25 декабря 2016 года . Проверено 30 сентября 2017 г.
- ^ Jump up to: а б с Антон 2013 , с. 30–33.
- ^ Карбоне, К.; Мэддокс, Т.; Фанстон, Пи Джей; Миллс, MGL; Гретер, ГФ; Ван Валкенбург, Б. (2009). «Параллели между воспроизведением и просачиванием смолы плейстоцена позволяют предположить социальность вымершего саблезубого кота Смилодона » . Письма по биологии . 5 (1): 81–85. дои : 10.1098/rsbl.2008.0526 . ПМЦ 2657756 . ПМИД 18957359 .
- ^ Киффнер, К. (2009). «Совпадение или свидетельство: был ли саблезубый кот Смилодон общительным?» . Письма по биологии . 5 (4): 561–562. дои : 10.1098/rsbl.2009.0008 . ПМК 2781900 . ПМИД 19443504 .
- ^ Ван Валкенбург, Б.; Мэддокс, Т.; Фанстон, Пи Джей; Миллс, MGL; Гретер, ГФ; Карбоне, К. (2009). «Социальность на ранчо Ла Бреа Смилодон : аргументы в пользу «доказательств» над «совпадением» » . Письма по биологии . 5 (4): 563–564. дои : 10.1098/rsbl.2009.0261 . ПМК 2781931 .
- ^ Хилд, Ф. (1989). « Травмы и заболевания Smilodon Californicus Bovard, 1904, (Mammalia, Felidae) из Ранчо Ла Бреа, Калифорния». Журнал палеонтологии позвоночных . 9 (3): 24A.
- ^ Балиси, Массачусетс; Шарма, АК; Ховард, CM; Шоу, Калифорния; Клэппер, Р.; Линдси, Эмили Л. (2020). «Компьютерная томография выявляет дисплазию тазобедренного сустава у смилодона: последствия для социального поведения вымершего хищника плейстоцена». bioRxiv 10.1101/2020.01.07.897348 .
- ^ Jump up to: а б МакКолл, С.; Неаполь, В.; Мартин, Л. (2003). «Оценка поведения вымерших животных: был ли Смилодон социальным?» . Мозг, поведение и эволюция . 61 (3): 159–164. дои : 10.1159/000069752 . ПМИД 12697957 . S2CID 2756104 .
- ^ Балиси, Майрин А.; Шарма, Абхинав К.; Ховард, Кэрри М.; Шоу, Кристофер А.; Клэппер, Роберт; Линдси, Эмили Л. (2021). «Компьютерная томография выявила дисплазию тазобедренного сустава у вымершего плейстоценового саблезубого кота Смилодона» . биоRxiv . 11 (1): 21271. doi : 10.1101/2020.01.07.897348 . ПМЦ 8553773 . ПМИД 34711910 . S2CID 235663241 .
- ^ Радинский, Л.Б. (1975). «Эволюция кошачьего мозга». Мозг, поведение и эволюция . 11 (3–4): 214–254. дои : 10.1159/000123636 . ПМИД 1181005 .
- ^ Ямагучи, Н.; Китченер, AC; Гилиссен, Э.; Макдональд, Д.В. (2009). «Размер мозга льва ( Panthera leo ) и тигра ( P. tigris ): значение для внутриродовой филогении, внутривидовые различия и последствия содержания в неволе». Биологический журнал Линнеевского общества . 98 (1): 85–93. дои : 10.1111/j.1095-8312.2009.01249.x .
- ^ Чименто, Северная Каролина; Агнолин, Флорида; Сойбельзон, Л.; Очоа, JG; Буйде, В. (2019). «Доказательства внутривидовых агонистических взаимодействий в популяции смилодонов (Carnivora, Felidae)» . Аккаунты Палевол . 18 (4): 449–454. Бибкод : 2019CRPal..18..449C . дои : 10.1016/j.crpv.2019.02.006 .
- ^ О'Брайен, DM (2019). «Статическое масштабирование и эволюция экстремальных размеров клыков у саблезубой кошки ( Smilodon Fatalis )» . Интегративная и сравнительная биология . 59 (5): 1303–1311. дои : 10.1093/icb/icz054 . ПМИД 31120517 .
- ^ Jump up to: а б Рейнольдс, Эшли Р.; Сеймур, Кевин Л.; Эванс, Дэвид К. (7 января 2021 г.). «Братцы и сестры Smilodon Fatalis раскрывают историю жизни саблезубой кошки» . iScience . 24 (1): 101916. Бибкод : 2021iSci...24j1916R . дои : 10.1016/j.isci.2020.101916 . ПМЦ 7835254 . ПМИД 33532710 .
- ^ Дойч, Арканзас; Лангерганс, РБ; Флорес, Д; Хартстон-Роуз, А (2023). «Рев Ранчо Ла Бреа? Сравнительная анатомия современных и ископаемых подъязычных костей кошачьих» . Журнал морфологии . 284 (10): e21627. дои : 10.1002/jmor.21627 . ПМИД 37708512 . S2CID 261090355 .
- ^ «Кинжалообразные клыки саблезубых кошек вырастали годами» . ScienceDaily . 01 июля 2015 г. Архивировано из оригинала 02 июля 2015 г. Проверено 2 июля 2015 г.
- ^ Мильбахлер, MC; Высоцкий, Массачусетс; Феранец, РС; Ценг, З.Дж.; Бьернссон, CS (01 июля 2015 г.). «Использование нового метода абсолютного онтогенетического определения возраста для расчета времени прорезывания зубов у саблезубой кошки Smilodon Fatalis » . ПЛОС ОДИН . 10 (7): e0129847. Бибкод : 2015PLoSO..1029847W . дои : 10.1371/journal.pone.0129847 . ПМЦ 4489498 . ПМИД 26132165 .
- ^ Феранец, Р.К. (2004). «Изотопные доказательства развития саблезубых зубов, скорости роста и диеты взрослой собаки Smilodon Fatalis с Ранчо Ла Бреа». Палеогеография, Палеоклиматология, Палеоэкология . 206 (3–4): 303–310. Бибкод : 2004PPP...206..303F . дои : 10.1016/j.palaeo.2004.01.009 .
- ^ «Факты свидетельствуют о том, что саблезубые кошки держались за молочные зубы, чтобы стабилизировать свои сабли» .
- ^ Ценг, ZJ (апрель 2024 г.). «Изменение характеристик изгиба во время продолжительной прорезывания клыков у саблезубых хищников: исследование Smilodon Fatalis » . Анатомическая запись . 307 (5). дои : 10.1002/ar.25447 . ПМИД 38588019 .
- ^ Лонг, К.; Протеро, Д.; Мадан, М.; Сиверсон, VJP; Смит, Т. (2017). «Вырастали ли саблезубые котята мускулистыми? Исследование постнатальной аллометрии костей конечностей у кошачьих из плейстоцена Ранчо Ла Бреа» . ПЛОС ОДИН . 12 (9): e0183175. Бибкод : 2017PLoSO..1283175L . дои : 10.1371/journal.pone.0183175 . ПМК 5617143 . ПМИД 28953899 .
- ^ Бьоркенгрен, АГ; Сарторис, диджей; Шермис, С.; Резник, Д. (1987). «Особенности паравертебрального окостенения у доисторической саблезубой кошки». Американский журнал рентгенологии . 148 (4): 779–782. дои : 10.2214/ajr.148.4.779 . ПМИД 3103404 .
- ^ Даклер, GL (1997). «Теменные впадины в черепах вымерших саблезубых кошачьих Smilodon Fatalis : свидетельства механического напряжения». Журнал палеонтологии позвоночных . 17 (3): 600–609. Бибкод : 1997JVPal..17..600D . дои : 10.1080/02724634.1997.10011006 .
- ^ Антон 2013 , с. 199.
- ^ Браун, К.; Балиси, М.; Шоу, Калифорния; Ван Валкенбург, Б. (2017). «Скелетная травма отражает охотничье поведение вымерших саблезубых кошек и ужасных волков». Экология и эволюция природы . 1 (5): 0131. Бибкод : 2017NatEE...1..131B . дои : 10.1038/s41559-017-0131 . ПМИД 28812696 . S2CID 8008808 .
- ^ Шмёкель, Хью; Фаррелл, Эслинг; Балиси, Майрин Ф. (2023). «Субхондральные дефекты, напоминающие рассекающий остеохондроз, на суставных поверхностях вымершего саблезубого кота Smilodon Fatalis и лютого волка Aenocyon dirus » . ПЛОС ОДИН 18 (7): e0287656. Бибкод : 2023PLoSO..1887656S . дои : 10.1371/journal.pone.0287656 . ПМЦ 10337945 . ПМИД 37436967 .
- ^ Мун, Чарльз А.; Пул, Рой Р.; Эрколи, Марк Д.; Цемент, Николас Р.; Барбоза, Фердинанд Х. из С.; Зурита, Альфред Э.; Сорок, Питер (22 мая 2023 г.). «Остеомиелит кисти руки популяции смилодонов (Felidae, Machairodontinae) из позднего плейстоцена Южной Америки» . Палеомир . 33 (2): 517–525. дои : 10.1016/j.palwor.2023.05.001 . ISSN 1871-174X . Проверено 5 февраля 2024 г. - через Elsevier Science Direct.
- ^ Хилл, Мэтью Г.; Истерла, Дэвид А. (1 мая 2023 г.). «Полный череп саблезубой кошки со Среднего континента Северной Америки и его эволюционный и экологический контекст» . Четвертичные научные обзоры . 307 : 108045. Бибкод : 2023QSRv..30708045H . doi : 10.1016/j.quascirev.2023.108045 . Проверено 27 апреля 2024 г. - через Elsevier Science Direct.
- ^ Кон, Мэтью Дж.; Маккей, Мориа П.; Найт, Джеймс Л. (1 августа 2005 г.). «Обед в плейстоцене. Кто в меню?» . Геология . 33 (8): 649–652. дои : 10.1130/G21476AR.1 . ISSN 1943-2682 . Проверено 27 апреля 2024 г. - через GeoScienceWorld.
- ^ Шеллхорн, Рико; Санмугараджа, Маюран (16 апреля 2014 г.). «Адаптации к среде обитания предплечья кошачьих» . ПалЗ . 89 (2): 261–269. дои : 10.1007/s12542-014-0230-8 . ISSN 0031-0220 . Проверено 27 апреля 2024 г. - через Springer.
- ^ Рейнольдс, Арканзас; Сеймур, КЛ; Эванс, округ Колумбия (2019). «Записи о кошачьих из Медисин-Хет, Альберта, позднего плейстоцена, включая первую канадскую запись о саблезубом коте Smilodon Fatalis». Канадский журнал наук о Земле . 56 (10): 1052–1060. Бибкод : 2019CaJES..56.1052R . doi : 10.1139/cjes-2018-0272 . hdl : 1807/96725 . S2CID 134586651 .
- ^ Вильявисенсио, Наталья А.; Линдси, Эмили Л.; Мартин, Фабиана М.; Борреро, Луис А.; Морено, Патрисио И.; Маршалл, Чарльз Р.; Барноски, Энтони Д. (февраль 2016 г.). «Сочетание людей, климата и изменения растительности спровоцировало позднечетвертичное вымирание мегафауны в регионе Ультима-Эсперанса, южная Патагония, Чили» . Экография . 39 (2): 125–140. Бибкод : 2016Экогр..39..125В . дои : 10.1111/ecog.01606 . ISSN 0906-7590 .
- ^ ДеСантис, LRG; Шуберт, Б.В.; Шмитт-Линвилл, Э.; Унгар, П.; Донохью, С.; Хаупт, Р.Дж. (15 сентября 2015 г.). Джон М. Харрис (ред.). «Текстуры стоматологической микроодежды хищных животных из смоляных ям Ла-Бреа, Калифорния, и потенциальные последствия их вымирания» (PDF) . Научная серия 42: Вклад в науку (специальный том под названием «Ла Бреа и за его пределами: палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия раскопок Музея естественной истории округа Лос-Анджелес на ранчо Ла Бреа). Музей естественной истории округа Лос-Анджелес: 37–52. Архивировано из оригинала (PDF) 20 декабря 2016 года . Проверено 6 февраля 2017 г.
- ^ Jump up to: а б Шерани, С. (2016). Новый метод оценки массы тела кошачьих, зависящий от особи (№ e2327v2) . Препринты PeerJ.
- ^ Манцуэтти, А.; Переа, Д.; Убилла, М.; Риндеркнехт, А. (2018). «Первая запись Smilodon Fatalis Leidy, 1868 (Felidae, Machairodontinae) во внеандском регионе Южной Америки (поздний плейстоцен, формация Сопас), Уругвай: таксономические и палеобиогеографические значения». Четвертичные научные обзоры . 180 : 57–62. Бибкод : 2018QSRv..180...57M . doi : 10.1016/j.quascirev.2017.11.024 .
- ^ Бохеренс, Х.; Котт, М.; Бонини, Р.; Скиан, Д.; Страчча, П.; Сойбельзон, Л.; Превости, Ф.Дж. (24 апреля 2016 г.). «Палеобиология популяции саблезубых кошек Smilodon в Пампейском регионе (провинция Буэнос-Айрес, Аргентина) вокруг последнего ледникового максимума: данные о стабильных изотопах углерода и азота в костном коллагене». Палеогеография, Палеоклиматология, Палеоэкология . 449 : 463–474. Бибкод : 2016PPP...449..463B . дои : 10.1016/j.palaeo.2016.02.017 . hdl : 11336/43965 .
- ^ Хейс, Б. (21 марта 2016 г.). «Саблезубые кошки были львами доисторической Южной Америки» . Новости науки УПИ . УПИ . Проверено 19 апреля 2016 г.
- ^ Мелоро, Карло; Элтон, Сара; Луис, Жюльен; Бишоп, Лаура К.; Дитчфилд, Питер (18 марта 2013 г.). «Кошки в лесу: прогнозирование адаптации среды обитания на основе морфометрии плечевой кости у современных и ископаемых кошачьих (Carnivora)» . Палеобиология . 39 (3): 323–344. Бибкод : 2013Pbio...39..323M . дои : 10.1666/12001 . ISSN 0094-8373 . Проверено 21 января 2024 г. - через Cambridge Core.
- ^ Мичен, Джули А.; О'Киф, Ф. Робин; Садлер, Редьярд В. (2014). «Эволюция саблезубой кошки Smilodon Fatalis в ответ на изменение климата в плейстоцене». Журнал эволюционной биологии . 27 (4): 714–723. дои : 10.1111/jeb.12340 . ПМИД 24779050 .
- ^ Jump up to: а б ДеСантис, LRG; Шуберт, Б.В.; Скотт, младший; Унгар, П.С. (2012). «Последствия диеты для вымирания саблезубых кошек и американских львов» . ПЛОС ОДИН . 7 (12): е52453. Бибкод : 2012PLoSO...752453D . дои : 10.1371/journal.pone.0052453 . ПМК 3530457 . ПМИД 23300674 .
- ^ Jump up to: а б Антон 2013 , с. 217–230.
- ^ Jump up to: а б О'Киф, Ф. Робин; Данн, Риган Э.; Вайцель, Элик М.; Уотерс, Майкл Р.; Мартинес, Лиза Н.; Биндер, Венди Дж.; Саутон, Джон Р.; Коэн, Джошуа Э.; Мичен, Джули А.; ДеСантис, Лариса Р.Г.; Кирби, Мэтью Э.; Геццо, Елена; Колтрейн, Джоан Б.; Фуллер, Бенджамин Т.; Фаррелл, Эслинг Б.; Такуэчи, Гэри Т.; Макдональд, Глен; Дэвис, Эдвард Б.; Линдси, Эмили Л. (2023). «Искоренение мегафауны до раннего дриаса на ранчо Ла Бреа связано с изменением состояния, вызванным пожаром». Наука . 381 (6659): eabo3594. дои : 10.1126/science.abo3594 . ПМИД 37590347 .
- ^ Бочеренс, Эрве (01 июня 2015 г.). «Изотопное отслеживание палеоэкологии крупных хищников в гигантской степи» . Четвертичные научные обзоры . 117 : 42–71. Бибкод : 2015QSRv..117...42B . doi : 10.1016/j.quascirev.2015.03.018 . ISSN 0277-3791 .
- ^ Превости, Франциско Дж.; Мартин, Фабиана М. (14 августа 2013 г.). «Палеоэкология гильдии хищников-млекопитающих Южной Патагонии во время позднего плейстоцена: экоморфология, стабильные изотопы и тафономия» . Четвертичный интернационал . Ранжирование мест обитания и процесс колонизации человеком Южной Америки. 305 : 74–84. Бибкод : 2013QuInt.305...74P . дои : 10.1016/j.quaint.2012.12.039 . hdl : 11336/84524 . ISSN 1040-6182 .
- ^ Антон 2013 , с. 223.
- ^ Jump up to: а б Стюарт, Энтони Дж. (20 августа 2022 г.). «Глава 6. Северная Америка: мастодонты, наземные ленивцы и саблезубые кошки». Исчезнувшие гиганты: Затерянный мир ледникового периода . Издательство Чикагского университета. стр. 67–112. ISBN 978-0-226-82403-1 .
- ^ О'Киф, Франция; Фет, Е.В.; Харрис, Дж. М. (2009). «Компиляция, калибровка и синтез радиоуглеродных дат фауны и флоры, Ранчо Ла Бреа, Калифорния» . Вклад в науку . 518 : 1–16. дои : 10.5962/стр.226783 . S2CID 128107590 .
- ^ Хилл, Мэтью Г.; Истерла, Дэвид А. (май 2023 г.). «Полный череп саблезубой кошки со Среднего континента Северной Америки и его эволюционный и экологический контекст» . Четвертичные научные обзоры . 307 : 108045. Бибкод : 2023QSRv..30708045H . doi : 10.1016/j.quascirev.2023.108045 . S2CID 257861663 .
- ^ Прието, Альфред; Лабарка, Рафаэль; Сьерпе, Виктор (2010). «Новые свидетельства существования саблезубого кота смилодона (Carnivora: Machairodontinae) в позднем плейстоцене южной чилийской Патагонии» . Чилийский журнал естественной истории . 83 (2). дои : 10.4067/S0716-078X2010000200010 .
- ^ Фидель, Стюарт (2009). «Внезапные смерти: хронология терминального вымирания мегафауны плейстоцена» (PDF) . В Хейнсе, Гэри (ред.). Вымирание американской мегафауны в конце плейстоцена . Палеобиология и палеоантропология позвоночных. Спрингер . стр. 21–37. дои : 10.1007/978-1-4020-8793-6_2 . ISBN 978-1-4020-8792-9 .
Библиография
[ редактировать ]- Энтони, М. (2013). Саблезубый (1-е изд.). Блумингтон: Издательство Университета Индианы. ISBN 978-0-253-01042-1 . ОСЛК 857070029 .

- Смилодон
- Высшие хищники
- Голоценовые вымирания
- Ископаемые таксоны, описанные в 1842 году.
- Роды доисторических хищников
- Плейстоценовые хищники
- Первые появления в плейстоцене
- Вымирание родов плейстоцена
- Плейстоценовые млекопитающие Северной Америки
- Плейстоценовые млекопитающие Южной Америки
- Смилодонтини
- Таксоны, названные Питером Вильгельмом Лундом
- Формирование Сопас
- Символы Калифорнии