Гаплогруппа U
| Гаплогруппа U | |
|---|---|
| Возможное время происхождения | 46 500 ± 3 300 лет назад [ 1 ] |
| Возможное место происхождения | Западная Азия, [ 2 ] |
| Предок | Р |
| Потомки | U1, U5, U6, U2'3'4'7'8'9 |
| Определение мутаций | 11467, 12308, 12372 [ 3 ] |
Гаплогруппа U — это гаплогруппа митохондриальной ДНК человека (мтДНК). Клада возникла из гаплогруппы R , вероятно, в период раннего верхнего палеолита . Его различные субклады (обозначенные U1–U9, расходившиеся на протяжении верхнего палеолита) широко распространены в Северной и Восточной Европе , Центральной , Западной и Южной Азии , а также в Северной Африке , на Африканском Роге и на Канарских островах. .
Происхождение
[ редактировать ]Гаплогруппа U происходит от ветви мтДНК гаплогруппы R филогенетического дерева. По оценкам, определяющие мутации (A11467G, A12308G, G12372A) возникли между 43 000 и 50 000 лет назад, в раннем верхнем палеолите (около 46 530 ± 3 290 лет до настоящего времени, с 95% доверительным интервалом согласно Behar et al., 2012). .
Древняя ДНК , отнесенная к митохондриальной гаплогруппе U*, была обнаружена в останках человеческого скелета, найденных в Западной Сибири и датированных ок. 45 000 лет назад. [ 4 ] Митогеном (33-кратное покрытие) особи Peştera Muierii 1 (PM1) из Румынии (35 тысяч лет назад) был идентифицирован как базальная гаплогруппа U6 *, ранее не встречавшаяся ни у одного древнего или современного человека. [ 5 ]
Гаплогруппа U была обнаружена среди иберомаврских экземпляров, датируемых эпипалеолитом, на доисторических стоянках Тафоральт и Афалу . [ 6 ] Среди особей Тафоральта около 13% наблюдаемых гаплотипов принадлежали к различным субкладам U, включая U4a2b (1/24; 4%), U4c1 (1/24; 4%) и U6d3 (1/24; 4%). Еще 41% проанализированных гаплотипов можно отнести либо к гаплогруппе U, либо гаплогруппе H. к Среди особей Афалу 44% проанализированных гаплотипов можно отнести либо к гаплогруппе U, либо к гаплогруппе H (3/9; 33%). [ 7 ]
Гаплогруппа U также наблюдалась среди древнеегипетских мумий, раскопанных на археологическом памятнике Абусир-эль-Мелек в Среднем Египте, датированных I тысячелетием до нашей эры, 13 из 90 мумий, несущих гаплогруппу U (все носители U позднего периода) и различные субклады. из них U, U1, U3, U5, U6, U7 и U8. [ 8 ] а в отдельном исследовании ДНК извлекли из зуба мумифицированной головы гораздо более древней мумии, существовавшей около 4000 лет назад Джехутинахта самого конца 11-й или начала 12-й династии, принадлежавшего к гаплогруппе мтДНК U5b2b5 (точных совпадений не обнаружено). в современной популяции носителей U5) из статьи Одиль Лорей с соавт., 2018 г. [ 9 ]
Кроме того, гаплогруппа U была обнаружена в древних окаменелостях гуанчей , раскопанных на Гран-Канарии и Тенерифе на Канарских островах , которые были датированы радиоуглеродом между 7 и 11 веками нашей эры. Все особи, являющиеся носителями клады, были захоронены на стоянке Тенерифе, причем эти экземпляры оказались принадлежащими к субкладам U6b1a (4/7; 57%) и U6b (1/7; 14%). [ 10 ]
Распределение
[ редактировать ]Гаплогруппа U встречается у 15% индийских каст и 8% индийских племен. [ 11 ] Гаплогруппа U встречается примерно у 11% коренных европейцев и считается старейшей материнской гаплогруппой, обнаруженной в этом регионе. [ 11 ] [ 12 ] [ 13 ] В исследовании 2013 года все древние современные человеческие последовательности из Европы, кроме одной, принадлежали к материнской гаплогруппе U, что подтверждает предыдущие выводы о том, что гаплогруппа U была доминирующим типом митохондриальной ДНК (мтДНК) в Европе до распространения сельского хозяйства в Европу из Ближний Восток. [ 14 ]
Гаплогруппа U имеет различные субклады с номерами от U1 до U9. Гаплогруппа K является субкладом U8. [ 15 ] Старость привела к широкому распространению потомков подгрупп по Западной Евразии, Северной Африке и Южной Азии. Некоторые субклады гаплогруппы U имеют более конкретный географический ареал.
Субклады
[ редактировать ]Субклады обозначены U1 – U9; Гаплогруппа K является субкладом U8.
Ван Овен и Кайзер (2009) предложили субклады «U2'3'4'7'8» и «U4'9». [ 3 ] Бехар и др. (2012) внесли поправки, сгруппировав «U4'9» как подчиненный «U2'3'4'7'8» для нового промежуточного субклада «U2'3'4'7'8'9».
Гаплогруппа U
[ редактировать ]Базальный U был найден в останках древнего североевразийского мальтийского мальчика возрастом 26 000 лет (МА1).
Гаплогруппа U1
[ редактировать ]Субклады U1: U1a (с глубокими субкладами U1a1, U1a1a, U1a1a1, U1a1b). [ 16 ] и U1b. [ 16 ]
По оценкам, гаплогруппа U1 возникла между 26 000 и 37 000 лет назад. Встречается очень редко по всей Европе. Чаще наблюдается в Восточной Европе, Анатолии и на Ближнем Востоке. Он также встречается на низких частотах в Индии. U1 встречается в Сванетском регионе Грузии в количестве 4,2%. Субклад U1a встречается от Индии до Европы, но крайне редко встречается на северных и атлантических окраинах Европы, включая Британские острова и Скандинавию. В Индии U1a был обнаружен в регионе Керала . U1b имеет такое же распространение, но встречается реже, чем U1a. Разновидность субклада U1b1 с мутациями G14070A! и A3426G обнаружен у евреев-ашкенази . [ 17 ] Субклады U1a и U1b встречаются с одинаковой частотой в Восточной Европе. [ 18 ]
Редкая клада U1 также встречается у алжирцев в Оране (0,83–1,08%) и регибат сахари племени (0,93%). [ 19 ]
Субклад U1a1a наблюдался у древнего человека, раскопанного на кладбище Келлис 2 в оазисе Дакле , расположенном в юго-западной пустыне Египта. 21 захоронение в Келлисе было датировано радиоуглеродом примерно 80–445 гг. н. э., то есть периодом романо-христианского периода. [ 20 ] Гаплогруппа U1 также была обнаружена среди образцов на материковом кладбище в Кулубнарти , Судан , которые датируются раннехристианским периодом (550-800 гг. н. э.). [ 21 ]
Анализ ДНК раскопанных останков, которые сейчас находятся в руинах церкви Святого Августина в Гоа , Индия, также выявил уникальный субклад мтДНК U1b. Эта подлиния отсутствует в Индии, но присутствует в Грузии и прилегающих регионах. [ 22 ] Поскольку генетический анализ подтверждает археологические и литературные данные, считается, что раскопанные останки принадлежат Кетеван Мученице , царице Грузии. [ 22 ]
Гаплогруппа U5
[ редактировать ]Возраст U5 оценивается от 25 000 до 35 000 лет. [ 23 ] примерно соответствует Граветтской культуре. Примерно 11% европейцев (10% американцев европейского происхождения) имеют тот или иной вариант гаплогруппы U5. Гаплогруппа, скорее всего, возникла в Европе. [ 24 ] [ 25 ]
U5 был преобладающей мтДНК мезолитических западных охотников-собирателей (WHG).
U5 был обнаружен в человеческих останках эпохи мезолита в Англии, Германии, Литве, Польше, Португалии, России, [ 26 ] Швеция, [ 27 ] Франция [ 28 ] и Испания. [ 29 ] Неолитические скелеты (возрастом около 7000 лет), которые были раскопаны в пещере Авелланер в Каталонии , на северо-востоке Испании , включали образец, несущий гаплогруппу U5. [ 30 ]
Гаплогруппа U5 и ее субклады U5a и U5b сегодня образуют самую высокую концентрацию населения на крайнем севере среди саамов , финнов и эстонцев . Однако он широко распространен на более низких уровнях по всей Европе. Такое распределение и возраст гаплогруппы указывают на то, что особи, принадлежащие к этой кладе, были частью первоначальной экспансии, сопровождавшей отступление ледниковых щитов из Европы около 10 000 лет назад.
Современные баски и кантабрийцы обладают почти исключительно линиями U5b (U5b1f, U5b1c1, U5b2). [ 31 ] [ 32 ]
Кроме того, гаплогруппа U5 встречается с небольшой частотой и в гораздо меньшем разнообразии на Ближнем Востоке и в некоторых частях Северной Африки (районы со значительной концентрацией U6), что предполагает обратную миграцию людей из Европы на юг. [ 33 ]
Митохондриальная гаплогруппа U5a также была связана с ВИЧ- инфицированными людьми, у которых наблюдалось ускоренное развитие СПИДа и смерть. [ 34 ]
U5 была основной гаплогруппой мезолитических европейских охотников-собирателей. Гаплогруппы U присутствовали у 83% европейских охотников-собирателей до того, как приток ближневосточных фермеров и степных индоевропейских предков снизил их частоту до менее чем 21%. [ 20 ]
- U5
- U5a возник около 17 000 и 27 000 лет назад.
- U5a1 возник между 14 000 и 20 000 лет назад. Найден у этрусского человека (700-600 гг. до н.э.) из южной Этрурии , Италия . [ 35 ]
- U5a1a возник между 8000 и 16000 лет назад.
- U5a1a1 возник между 3000 и 11 000 лет назад.
- U5a1a1a возникла менее 6000 лет назад. [ 36 ]
- U5a1a1b возник примерно между 600 и 6000 лет назад.
- U5a1a1c
- U5a1a1d возник менее 4300 лет назад
- U5a1a2 возник между 7000 и 14000 лет назад.
- U5a1a2a возникла менее 5400 лет назад.
- U5a1a2a1 возник менее 3400 лет назад.
- U5a1a2a возникла менее 5400 лет назад.
- U5a1a1 возник между 3000 и 11 000 лет назад.
- U5a1b возник между 6000 и 11000 лет назад.
- U5a1b1 возник между 5000 и 9000 лет назад.
- U5a1b1a возник между 2500 и 7500
- U5a1b1a1 менее 4000 лет назад
- U5a1b1b возник менее 8000 лет назад.
- U5a1b1c возник между 3000 и 7000 лет назад.
- U5a1b1c1 возник менее 5000 лет назад.
- U5a1b1c2 возник менее 5000 лет назад.
- U5a1b1d
- U5a1b1e
- U5a1b1a возник между 2500 и 7500
- U5a1b2
- U5a1b3
- U5a1b1 возник между 5000 и 9000 лет назад.
- u5a1c
- U5a1c1
- U5a1c2
- U5a1d возник около 19 000 лет назад.
- U5a1d1
- U5a1d2
- U5a1d2a
- U5a1e
- U5a1f
- U5a1a возник между 8000 и 16000 лет назад.
- U5a2 возник около 14000 лет назад.
- U5a2a возникла около 6000 лет назад. Он был найден в древнем мезолитическом образце (6000-5000 лет до нашей эры) из пещеры Сантимаминье в Стране Басков , Испания . [ 37 ]
- U5a2b возник около 8000 лет назад.
- U5a2c возник около 13000 лет назад.
- U5a2d Найден на мезолитической стоянке Хусеби Клев на западе Швеции.
- U5a2e
- U5a1 возник между 14 000 и 20 000 лет назад. Найден у этрусского человека (700-600 гг. до н.э.) из южной Этрурии , Италия . [ 35 ]
- U5b возник между 19 000 и 26 000 лет назад. [ 38 ] и имеет полиморфизмы в 150 7768 14182 (+ U5 полиморфизмы). Встречается среди сива-берберов оазиса Сива . [ 39 ]
- U5b1 возник между 11 000 и 20 000 лет назад. [ 40 ]
- U5b1a
- U5b1b: был обнаружен у саамов Скандинавии, финнов и берберов Северной Африки, у которых, как выяснилось, была одна очень молодая ветвь, возраст которой составлял всего около 9000 лет. U5b1b также был обнаружен у фульбе и папеля в Гвинее-Бисау и якутов северо-восточной Сибири . [ 41 ] [ 42 ] Он возник около 11000 лет назад.
- U5b1b1
- U5b1b1b Основной элемент материнской западно-евразийской линии в Пуэрто-Рико , соответствующий образцам из Сенегамбии и северного Камеруна, что указывает на его присутствие как продукт ранней колонизации и порабощения сенегамбийцев. [ 43 ]
- U5b1b1
- U5b1c возник около 13 000 лет назад.
- U5b1d
- U5b1e возник около 6600 лет назад. U5b1e в основном встречается в Центральной Европе среди чехов, словаков, венгров и южных русских. [ 44 ]
- U5b1g
- U5b2 возник между 17 000 и 23 000 лет назад. [ 45 ] и имеет полиморфизмы 1721 13637(+ U5b полиморфизмы). Клада была обнаружена в останках, датируемых доисторическими временами в Европе, таких как субклад U5b2c1 человека из Ла-Бранья (найденный на стоянке Ла-Бранья в Испании). U5b2 редко встречается у французских басков (2,5%) и чаще встречается у испанских басков.
- U5b2a между 12 000 и 19 000 лет назад, [ 46 ] распространен в Центральной Европе. [ 44 ]
- U5b2a1 между 9 000 и 18 000 лет назад.
- U5b2a2 между 7000 и 14000 лет назад,
- U5b2a3 между 3000 и 14 000 лет назад.
- U5b2a4 между 1000 и 10 000 лет назад
- U5b2a5 менее 2600 лет назад
- U5b2a6 менее 12 000 лет назад.
- U5b2b между 12 000 и 17 000 лет назад. [ 47 ] Эта клада была тесно связана с Неве, которая на момент ее открытия была старейшим обнаруженным захоронением младенцев женского пола в Европе, датированным радиоуглеродом примерно 10 000 лет назад. [ 48 ] [ 49 ]
- U5b2c между 7000 и 18000 лет назад. [ 50 ]
- U5b2c1 менее 8000 лет назад. [ 51 ] Найден у финикийца из карфагенской гробницы на холме Бирса , Тунис. [ 52 ]
- U5b2a между 12 000 и 19 000 лет назад, [ 46 ] распространен в Центральной Европе. [ 44 ]
- U5b3: Субклад, вероятно, происходит с итальянского полуострова; [ 44 ] его наибольшее распространение наблюдается в юго-западной Европе, с пиком среди сардинцев (3,84%), за которыми следуют жители Балеарских островов (1,56%) и северной части материковой части Португалии (1,09%). [ 53 ] Согласно другому исследованию, U5b3 встречается с частотой 2,53% среди жителей Майорки и 0,96% среди сефардских чуэта . [ 54 ]
- U5b3b: этот субклад, вероятно, имеет сходное западно-средиземноморское/иберо-итальянское происхождение, но распространился на части северо-западной и средней Европы посредством римской экспансии, образцы найдены на Крите (Греция) , в Испании , Центральной Италии , Англии , Немецком Пфальце и Богемии. . [ 55 ]
- U5b1 возник между 11 000 и 20 000 лет назад. [ 40 ]
- U5a возник около 17 000 и 27 000 лет назад.
Гаплогруппа U6
[ редактировать ]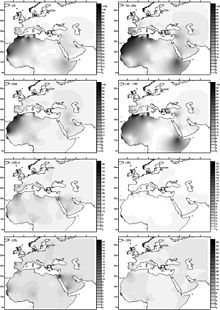
Бехар и др. датировали гаплогруппу U6 периодом от 31 000 до 43 000 лет назад. (2012). Базальный U6* был обнаружен в румынском образце древней ДНК ( Peştera Muierilor ), датированном 35 000 лет назад. [ 56 ] Хервелла и др. (2016) рассматривают эту находку как свидетельство палеолитической обратной миграции Homo sapiens из Евразии в Африку. Открытие базального U6* в древней ДНК способствовало тому, что предполагаемый возраст U6 был отброшен примерно на 46 000 лет назад. [ 57 ]
Обычно генетическую историю U6 рассматривают как миграцию из Юго-Западной Азии через Северную Африку. Эта гипотеза основана на общем происхождении субкладов гаплогруппы U в Юго-Западной Азии, которая также является центром географического распространения субкладов U: Европа, Индия, Центральная Азия, Восточная Африка и Северная Африка. Можно выдвинуть два возможных сценария развития первого гаплотипа U6 (несущего мутации 3348 и 16172): i) эти мутации возникли в регионе-основателе, но не оставили там никакого генетического наследия в нынешних человеческих популяциях; ii) они возникли, вероятно, где-то в Северной Африке, после прибытия гаплотипа-основателя U6. В Северной Африке U6 встречается значительно лишь на ее западной окраине (а также в Юго-Западной Европе). Что еще более важно, все самые базальные ветви практически ограничены этой областью (U6b, U6c и U6d), что может указывать на ее западное происхождение. Тем не менее, нельзя исключить основной субклад U6a, который демонстрирует богатство субкладов в Северо-Западной Африке, хотя некоторые производные ветви также включают последовательности из популяций Восточной Африки и Ближнего Востока (например, U6a2).
Гаплогруппа U6 распространена (распространенность около 10%). [ 33 ] в Северо-Западной Африке (максимум 29% у алжирских мозабитов [ 58 ] ) и Канарские острова (в среднем 18% с пиковой частотой 50,1% на Ла-Гомере ). Он также встречается на Пиренейском полуострове , где он имеет наибольшее разнообразие (10 из 19 подлиний встречаются только в этом регионе, а не в Африке). [ 59 ] Северо-Восточная Африка и иногда в других местах. U6 также встречается на низких частотах в бассейне Чада , включая редкую Канарскую ветвь. Это предполагает, что древние носители клады U6, возможно, населяли бассейн Чада или проходили через него на пути на запад к Канарским островам. [ 60 ]
Считается, что U6 проник в Северную Африку с Ближнего Востока около 30 000 лет назад. Он был найден среди иберомаврских экземпляров, датируемых эпипалеолитом, на доисторическом стоянке Тафоральт . [ 61 ] Несмотря на самое высокое разнообразие иберийского U6, Мака-Мейер утверждает, что эта клада имеет ближневосточное происхождение, основываясь на самом высоком разнообразии субклада U6a в этом регионе. [ 59 ] куда он прибыл бы из Западной Азии, при этом иберийская заболеваемость в первую очередь представляет собой миграцию из Магриба , а не сохранение европейской коренной популяции. [ почему? ]
По данным Эрнандеса и др. 2015 г. «предполагаемое проникновение североафриканских линий U6 в Иберию в 10 тыс. лет хорошо коррелирует с другими L-африканскими кладами, указывая на то, что U6 и некоторые линии L переместились вместе из Африки в Иберию в раннем голоцене ». [ 62 ]
U6 имеет четыре основных субклада: [ 59 ] [ 61 ]
Подгруппа U6a отражает первую африканскую экспансию из Магриба, возвращающуюся на восток. Производная клада U6a1 сигнализирует о движении назад из Восточной Африки обратно в Магриб и на Ближний Восток. Эта миграция совпадает с вероятной афроазиатской языковой экспансией. Клады U6b и U6c, ограниченные Западной Африкой, имели более локализованное распространение. U6b, вероятно, достиг Пиренейского полуострова во время распространения капсов в Северной Африке. Две автохтонные производные этих клад (U6b1 и U6c1) указывают на прибытие североафриканских поселенцев на Канарский архипелаг в доисторические времена, скорее всего, из-за высыхания Сахары. Отсутствие этих канарских линий в настоящее время в Африке предполагает важные демографические движения в западной части этого континента.
- Бут-Мейер 2003 г.
- U6a: субклад является наиболее распространенным, простирающимся от Канарских островов и Пиренейского полуострова до Африканского Рога и Ближнего Востока. Субгаплогруппа имеет наибольшее разнообразие в Северо-Восточной Африке. Анализ древней ДНК иберомаврских скелетных останков на стоянке Тафоральт в Марокко , которые были датированы поздним каменным веком между 15 100 и 13 900 лн, выявил субклад U6a среди большинства окаменелостей (6/7; ~ 86%). [ 63 ] Было обнаружено, что окаменелости на стоянке раннего неолита Ифри-н'Амр или Мусса в Марокко, датированные примерно 5000 годом до нашей эры, также содержат субгаплогруппу U6a. Эти древние люди несли автохтонный геномный компонент Северо-Западной Африки, который достигает максимума среди современных берберов , что указывает на то, что они были предками населения в этом районе. [ 64 ] Предполагаемый возраст U6a составляет 24–27 500 лет назад. Он имеет один основной субклад:
- U6b: показывает более исправленный и западный дистрибутив. На Пиренейском полуострове U6b чаще встречается на севере, тогда как U6a чаще встречается на юге. Он также был обнаружен на низких частотах в Марокко, Алжире, Сенегале и Нигерии. Предполагаемый возраст: 8 500–24 500 лет назад. Имеет один субклад:
- U6b1: встречается только на Канарских островах и Пиренейском полуострове. Предполагаемый возраст: гр. 6000 лет назад.
- U6c: встречается только в Марокко и на Канарских островах. Предполагаемый возраст: 6 000–17 500 лет назад.
- U6d: наиболее тесно связан с U6b. Локализуется в Магрибе, присутствует в Европе. Он возник между 10 000 и 13 000 лет назад.
U6a, U6b и U6d имеют общую базальную мутацию (16219), которой нет в U6c, тогда как U6c имеет 11 уникальных мутаций. U6b и U6d имеют общую мутацию (16311), которой нет у U6a, который имеет три уникальные мутации.
U2'3'4'7'8'9
[ редактировать ]Субклады U2, U3, U4, U7, U8 и U9 теперь считаются монофилетическими, их общим предком. «U2'3'4'7'8'9» определяется мутацией A1811G, возникшей примерно между 42 000 и 48 000 лет назад (Behar et al., 2012). Внутри U2'3'4'7'8'9 U4 и U9 могут быть монофилетическими, как «U4'9» (мутации T195C!, G499A, T5999C), возникшие между 31 000 и 43 000 лет назад (Behar et al., 2012). .
U2'3'4'7'8'9 был найден в останках двух древних жителей Северной Сибири (ANS) возрастом 32 000 лет на стоянке RHS Яна на реке Яна . [ 66 ]
Гаплогруппа U2
[ редактировать ]Гаплогруппа U2 наиболее распространена в Южной Азии. [ 67 ] но также встречается с низкой частотой в Центральной и Западной Азии, а также в Европе как U2e (европейская разновидность U2 называется U2e). [ 68 ] Общая частота U2 в Южной Азии в основном объясняется группой U2i в Индии, тогда как гаплогруппа U2e, распространенная в Европе, встречается редко; учитывая, что эти линии разошлись примерно 50 000 лет назад, эти данные были интерпретированы как указывающие на очень низкий поток генов по материнской линии между Южной Азией и Европой на протяжении всего этого периода. [ 67 ] Примерно половина U мтДНК в Индии принадлежит к специфичным для Индии ветвям гаплогруппы U2 (U2i: U2a, U2b и U2c). Гаплогруппа U2b2 была обнаружена в останках 4500-летней женщины, раскопанных на стоянке Ракхигархи цивилизации долины Инда , в современном штате Харьяна, Индия. [ 69 ] Хотя U2 обычно встречается в Индии, [ 67 ] он присутствует и у ногайцев , потомков различных монгольских и тюркских племен, образовавших Ногайскую Орду . [ 70 ] И У2, и У4 встречаются у кетов и нганасан , коренных жителей бассейна реки Енисей и полуострова Таймыр . [ 71 ]
Субклады U2: U2a, [ 72 ] и2б, [ 73 ] и2с, [ 74 ] U2d, [ 75 ] и U2e. [ 76 ] Поскольку специфичные для Индии субклады U2a, U2b и U2c вместе называются U2i, евразийская гаплогруппа U2d, по-видимому, является сестринской кладой индийской гаплогруппы U2c. [ 77 ] в то время как U2e считается субкладом, специфичным для Европы, но также встречается в Южной Индии. [ 68 ]
Гаплогруппа U2 была обнаружена в останках 37 000 человек. [ 78 ] и 30 000-летний охотник-собиратель из Костёнок Воронежской области в Центрально-Южно-Европейской части России. [ 79 ] в человеческих останках возрастом от 4800 до 4000 лет из памятника культуры кубков эпохи позднего неолита в Кромсдорфе , Германия, [ 80 ] и в человеческих останках возрастом 2000 лет из Бегебьергорда на юге Дании. Однако гаплогруппа U2 у современных скандинавов встречается редко. [ 81 ] Останки 2000-летнего западноевразийского мужчины гаплогруппы U2e1 были найдены на кладбище хунну на северо-востоке Монголии. [ 82 ]
Гаплогруппа U3
[ редактировать ]Гаплогруппа U3 делится на два субклада:: U3a. [ 83 ] и U3b. [ 83 ]
Возраст слияния U3a оценивается от 18 000 до 26 000 лет назад, а возраст слияния U3b - от 18 000 до 24 000 лет назад. U3a встречается в Европе, на Ближнем Востоке, на Кавказе и в Северной Африке. Почти полностью европейский субклад U3a1, датируемый 4000–7000 лет назад, предполагает относительно недавнее (поздний голоцен или позже) распространение этих линий в Европе. Существует минорный субклад U3c (производный от U3a), представленный единственной азербайджанской мтДНК с Кавказа. U3b широко распространен на Ближнем Востоке и на Кавказе, особенно в Иране, Ираке и Йемене, с небольшим европейским субкладом U3b1b, датируемым 2000–3000 лет назад. [ 84 ] Гаплогруппа U3 определяется HVR1 переходом A16343G. Он встречается в небольших количествах по всей Европе (около 1% населения), на Ближнем Востоке (около 2,5% населения) и в Центральной Азии (около 1% населения). U3 присутствует у сванов из региона Сванети (около 4,2% населения), а также среди литовских цыган, польских цыган и испанских цыган (36-56%). [ 85 ] [ 86 ] [ 87 ] [ 88 ]
Клада U3 также встречается у мозабитских берберов (10,59%), [ 19 ] а также египтяне в Эль-Хайезе (2,9%) [ 89 ] и оазисы Гурна (2,9%), [ 90 ] и алжирцы в Оране (1,08–1,25%). [ 19 ] Редкий субклад U3a встречается среди туарегов, населяющих Нигер (3,23%). [ 91 ] и среди сомалийцев (1,6%). [ 92 ]
Гаплогруппа U3 была обнаружена в некоторых из 6400-летних останков (U3a), обнаруженных в пещерах Вади-эль-Маккух недалеко от Иерихона, относящихся к периоду энеолита . [ 93 ] Гаплогруппа U3 уже присутствовала в генофонде Западной Евразии около 6000 лет назад, а также, вероятно, также ее субклад U3a. [ 93 ]
Гаплогруппа U4
[ редактировать ]Гаплогруппа U4 возникла между 21 000 и 14 000 лет назад. Его распространение связано с узким местом численности населения из-за последнего ледникового максимума . [ 85 ]
U4 был обнаружен в древней ДНК, [ 94 ] и это относительно редко встречается в современных популяциях, [ 44 ] хотя он встречается в значительных количествах у некоторых коренных народов Северной Азии и Северной Европы, поскольку связан с остатками древних европейских охотников-собирателей, сохранившимися у коренного населения Сибири. [ 95 ] [ 96 ] [ 97 ] U4 встречается у исчезающего народа нганасан полуострова Таймыр. [ 71 ] [ 98 ] у манси (16,3%), [ 97 ] и у кетов (28,9%) Енисея. [ 97 ] Он встречается в Европе, с самой высокой концентрацией в Скандинавии и странах Балтии. [ 99 ] и встречается у саамского населения Скандинавского полуострова (хотя у U5b более высокая представленность). [ 100 ] U4 также сохранился у народа калашей (текущая численность населения 3700 человек). [ 101 ] уникальное племя среди индоарийских народов Пакистана, где U4 (субклад U4a1 [ 102 ] ) достигает максимальной частоты 34%. [ 86 ] [ 103 ]
Субклады U4: U4a, [ 104 ] U4b, [ 105 ] U4c, [ 106 ] и U4d. [ 107 ]
возрастом от 7200 до 6000 лет Гаплогруппа U4 связана с древними европейскими охотниками-собирателями и была обнаружена в останках культуры каменной керамики на Готланде, Швеция, а также в останках возрастом от 4400 до 3800 лет из стоянки Дамсбо датской культуры кубков. . [ 108 ] [ 27 ] [ 109 ] Останки, идентифицированные как субклад U4a2, связаны с культурой шнуровой керамики , которая процветала от 5200 до 4300 лет назад в Восточной и Центральной Европе и охватывала большую часть континентальной северной Европы от реки Волги на востоке до Рейна на западе. [ 110 ] Митохондриальная ДНК, обнаруженная из останков возрастом от 3500 до 3300 лет на стоянке Бредтофтегард в Дании, связанной с скандинавским бронзовым веком, включает гаплогруппу U4 с 16179T в ее HVR1, указывающей на субклад U4c1. [ 109 ] [ 111 ] [ 112 ] 2 из 9 останков возрастом 1700 лет на крайнем юго-западе Ивановской области имели U4c1. [ 113 ]
Гаплогруппа U7
[ редактировать ]Гаплогруппа U7 считается специфичной для Западной Евразии гаплогруппой мтДНК, которая, как полагают, возникла в районе Черного моря примерно 30 000 лет назад. [ 67 ] [ 114 ] [ 115 ] В современных популяциях U7 встречается с низкой частотой на Кавказе. [ 115 ] западносибирские племена, [ 116 ] Западная Азия (около 4% на Ближнем Востоке, пик - 10% у иранцев), [ 67 ] Южная Азия (около 12% в Гуджарате, самом западном штате Индии, тогда как для всей Индии его частота держится около 2%, а в Пакистане - 5% ), [ 67 ] и народ ведды в Шри-Ланке, где он достигает самой высокой частоты - 13,33% (субклад U7a). [ 117 ] Одна треть мтДНК, специфичных для Западной Евразии, обнаруженных в Индии, относится к гаплогруппам U7 R2 и W. , Предполагается, что крупномасштабная иммиграция привела эти митохондриальные гаплогруппы в Индию. [ 67 ]
Субклады U7 - это U7a (с глубокими субкладами U7a1, U7a2, U7a2a, U7a2b). [ 118 ] и U7b. [ 118 ]
Генетический анализ лиц, связанных с поздней гальштатской культурой из земли Баден-Вюртемберг (Германия), считающихся примерами железного века, включал гаплогруппу U7. «княжеских захоронений» [ 119 ] Сообщается, что гаплогруппа U7 была обнаружена в человеческих останках возрастом 1200 лет (около 834 года) у женщины, предположительно принадлежащей к королевскому клану, которая была похоронена на корабле викингов Осеберг в Норвегии. [ 120 ] Гаплогруппа U7 была обнаружена в 1000-летних человеческих останках (датируемых примерно 1000–1250 гг. н. э.) на христианском кладбище в Конгемаркене, Дания. Однако U7 редко встречается среди современных этнических скандинавов. [ 116 ]
Субклад U7a особенно распространен среди саудовцев , составляя около 30% материнских линий в Восточной провинции . [ 121 ]
Гаплогруппа U8
[ редактировать ]- Гаплогруппа U8a: Баски имеют самую предковую филогению в Европе для митохондриальной гаплогруппы U8a. Это редкая подгруппа U8, что позволяет отнести баскское происхождение этой линии к верхнему палеолиту. Отсутствие линий U8a в Африке позволяет предположить, что их предки, возможно, произошли из Западной Азии. [ 15 ]
- Гаплогруппа U8b: эта клада была найдена в Италии и Иордании. [ 15 ]
- Гаплогруппа U8b'K: Эта клада может быть синонимом гаплогруппы K и гаплогруппы UK . [ нужна ссылка ]
Наиболее распространенным субкладом гаплогруппы U8b является гаплогруппа K , которая, по оценкам, возникла от 30 000 до 22 000 лет назад. [ нужна ссылка ] Гаплогруппа K составляет значительную часть линий мтДНК Европы и Западной Азии. Теперь известно, что на самом деле это субклад гаплогруппы U8b'K. [ 15 ] и, как полагают, впервые возник на северо-востоке Италии. Гаплогруппа Великобритании демонстрирует некоторые доказательства своей высокой защиты от прогрессирования СПИДа. [ 34 ]
Гаплогруппа U9
[ редактировать ]Гаплогруппа U9 представляет собой редкую кладу в филогении мтДНК, охарактеризованную лишь недавно в нескольких популяциях Пакистана (Quintana-Murci et al. 2004). Его присутствие в Эфиопии и Йемене , а также некоторые специфичные для Индии линии M в йеменской выборке указывают на поток генов вдоль побережья Аравийского моря. Гаплогруппы U9 и U4 имеют две общие мутации, лежащие в основе их филогении. Интересно, что в Пакистане U9 часто встречается только среди так называемой популяции Макрани . В этой конкретной популяции линии, специфичные для частей Восточной Африки, встречаются примерно в 39%, что позволяет предположить, что линии U9 в Пакистане могут иметь африканское происхождение (Quintana-Murci et al. 2004). Независимо от того, какое побережье Аравийского моря могло быть источником U9, его распространение в бассейне Эфиопии, Южной Аравии и Инда намекает на то, что диверсификация субклада от U4 могла произойти в регионах, далеких от нынешней области наибольшего разнообразия и частоты встречаемости. гаплогруппа U4 — Восточная Европа и Западная Сибирь. [ 122 ]
Известные участники
[ редактировать ]- Доктор Санджай Гупта является носителем гаплогруппы U2c. [ 123 ]
- Политический аналитик Линда Чавес является членом гаплогруппы U5a1a1. [ 124 ]
- Специалист по генетической генеалогии СиСи Мур принадлежит к гаплогруппе U5b1b2. [ 125 ]
См. также
[ редактировать ]- Генеалогический ДНК-тест
- Генетическая генеалогия
- Митохондриальная генетика человека
- Популяционная генетика
- Семь дочерей Евы
|
Филогенетическое древо гаплогрупп митохондриальной ДНК (мтДНК) человека | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева ( L ) | |||||||||||||||||||||||||||||||||||||||
| Л0 | Л1–6 | ||||||||||||||||||||||||||||||||||||||
| Л1 | Л2 | Л3 | Л4 | Л5 | Л6 | ||||||||||||||||||||||||||||||||||
| М | Н | ||||||||||||||||||||||||||||||||||||||
| Чехия | Д | И | Г | вопрос | ТО | А | С | Р | я | В | Х | И | |||||||||||||||||||||||||||
| С | С | Б | Ф | Р0 | до JT | П | В | ||||||||||||||||||||||||||||||||
| Х.В. | Джей Ти | К | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Ссылки
[ редактировать ]- ^ Бехар и др. (2012), обратите внимание на пересмотренную оценку 42–58 ключей, основанную на древней ДНК, сделанную Hervella et al. (2016).
- ^ Хервелла и др. (2016) «Люди, несущие гаплогруппу U, возможно, распространились на запад из Западной Азии около 39–52 тыс. лет назад, достигнув Европы, о чем свидетельствует гаплогруппа U5, и Северной Африки, о чем свидетельствует гаплогруппа U6»
- ^ Перейти обратно: а б ван Овен М., Кайзер М. (февраль 2009 г.). «Обновленное комплексное филогенетическое древо глобальных вариаций митохондриальной ДНК человека» . Человеческая мутация . 30 (2): Е386-94. дои : 10.1002/humu.20921 . ПМИД 18853457 . S2CID 27566749 .
- ^ Ларруга Дж.М., Марреро П., Абу-Амеро К.К., Голубенко М.В., Кабрера В.М. (май 2017 г.). «Носители митохондриальной ДНК макрогаплогруппы R колонизировали Евразию и Австралазию из центральной области Юго-Восточной Азии» . Эволюционная биология BMC . 17 (1): 115. Бибкод : 2017BMCEE..17..115L . дои : 10.1186/s12862-017-0964-5 . ПМЦ 5442693 . ПМИД 28535779 . Марреро П., Абу-Амеро К.К., Ларруга Х.М., Кабрера В.М. (ноябрь 2016 г.). «Носители митохондриальной ДНК человека макрогаплогруппы М колонизировали Индию из Юго-Восточной Азии» . Эволюционная биология BMC . 16 (1): 246. Бибкод : 2016BMCEE..16..246M . дои : 10.1186/s12862-016-0816-8 . ПМК 5105315 . ПМИД 27832758 .
- ^ Хервелла М., Свенссон Э.М., Альберди А., Гюнтер Т., Изагирре Н., Мунтерс А.Р. и др. (май 2016 г.). «Митогеном 35 000-летнего человека разумного из Европы поддерживает палеолитическую обратную миграцию в Африку» . Научные отчеты . 6 : 25501. Бибкод : 2016NatSR...625501H . дои : 10.1038/srep25501 . ПМЦ 4872530 . ПМИД 27195518 . .
- ^ Сечер Б., Фрегель Р., Ларруга Х.М., Кабрера В.М., Эндикотт П., Пестано Дж.Дж., Гонсалес А.М. (май 2014 г.). «История потока гена гаплогруппы U6 североафриканской митохондриальной ДНК на африканский, евразийский и американский континенты» . Эволюционная биология BMC . 14 (1): 109. Бибкод : 2014BMCEE..14..109S . дои : 10.1186/1471-2148-14-109 . ПМК 4062890 . ПМИД 24885141 .
- ^ Кефи Р., Хечми М., Науали С., Джмел Х., Хсуна С., Бузаид Е. и др. (январь 2018 г.). «О происхождении иберомавров: новые данные, основанные на древней митохондриальной ДНК и филогенетическом анализе популяций афалу и тафоральта». Митохондриальная ДНК Часть А. 29 (1): 147–157. дои : 10.1080/24701394.2016.1258406 . ПМИД 28034339 . S2CID 4490910 .
- ^ Шунеманн В.Дж., Пельцер А., Вельте Б., ван Пелт В.П., Молак М., Ван CC и др. (май 2017 г.). «Геномы древнеегипетских мумий предполагают увеличение африканского происхождения к югу от Сахары в постримские периоды» . Природные коммуникации . 8 : 15694. Бибкод : 2017NatCo...815694S . дои : 10.1038/ncomms15694 . ПМЦ 5459999 . ПМИД 28556824 .
- ^ Лорей, О.; Ратнаяке, С.; Базинет, Алабама; Стоквелл, ТБ; Соммер, Д.Д.; Роланд, Н.; Маллик, С.; Джонсон, Польша; Скоглунд, П.; Онорато, Эй Джей; Бергман, Нью-Хэмпшир; Райх, Д.; Ирвин, Дж. А. (2018). «Биологическое определение пола головы египетской мумии возрастом 4000 лет для оценки возможности восстановления ядерной ДНК из наиболее поврежденных и ограниченных судебно-медицинских образцов» . Гены . 9 (3): 135. doi : 10.3390/genes9030135 . ПМЦ 5867856 . ПМИД 29494531 .
- ^ Родригес-Варела Р., Гюнтер Т., Кшевинска М., Стора Дж., Гиллингуотер Т.Х., МакКаллум М. и др. (ноябрь 2017 г.). «Геномный анализ человеческих останков с Канарских островов доевропейского завоевания показывает близкое родство с современными жителями Северной Африки» . Современная биология . 27 (21): 3396–3402.e5. Бибкод : 2017CBio...27E3396R . дои : 10.1016/j.cub.2017.09.059 . hdl : 2164/13526 . ПМИД 29107554 .
- ^ Перейти обратно: а б Кармин М (2005). Гаплогруппа R митохондриальной ДНК человека в Индии (PDF) (магистерская диссертация). Тартуский университет.
- ^ Сайкс Б. (2001). Семь дочерей Евы . Лондон; Нью-Йорк: Бантам Пресс. ISBN 978-0-393-02018-2 .
- ^ «Материнское происхождение» . Оксфордские предки. Архивировано из оригинала 15 июля 2017 года . Проверено 7 февраля 2013 г.
- ^ Фу К., Миттник А., Джонсон П.Л., Бос К., Лари М., Боллонгино Р. и др. (апрель 2013 г.). «Пересмотренная временная шкала эволюции человека, основанная на древних митохондриальных геномах» . Современная биология . 23 (7): 553–559. Бибкод : 2013CBio...23..553F . дои : 10.1016/j.cub.2013.02.044 . ПМК 5036973 . ПМИД 23523248 .
- ^ Перейти обратно: а б с д Гонсалес А.М., Гарсиа О, Ларруга Х.М., Кабрера В.М. (май 2006 г.). «Митохондриальная линия U8a указывает на палеолитическое поселение в Стране Басков» . БМК Геномика . 7 (1): 124. дои : 10.1186/1471-2164-7-124 . ПМЦ 1523212 . ПМИД 16719915 .
- ^ Перейти обратно: а б Скотт Р. (2010). «Филогения mt-hg U3» (PDF) .
- ^ Брук, Кевин Алан (2022). Материнские генетические линии евреев-ашкенази . Пресса академических исследований. п. 100-101. ISBN 978-1-64469-984-3 .
- ^ «Проект гаплогруппы U1 мтДНК» . СемейноеДревоДНК .
- ^ Перейти обратно: а б с Бекада А., Арауна Л.Р., Деба Т., Калафель Ф., Бенхамамуш С., Комас Д. (24 сентября 2015 г.). «Генетическая гетерогенность в человеческой популяции Алжира» . ПЛОС ОДИН . 10 (9): e0138453. Бибкод : 2015PLoSO..1038453B . дои : 10.1371/journal.pone.0138453 . ПМЦ 4581715 . ПМИД 26402429 .
- ^ Перейти обратно: а б Молто Дж.Э., Лорей О., Маллотт Е.К., Малхи Р.С., Фаст С., Дэниелс-Хиггинботэм Дж. и др. (октябрь 2017 г.). «Полное секвенирование митохондриального генома захоронения на романо-христианском кладбище в оазисе Дахле, Египет: предварительные показания» . Гены . 8 (10): 262. doi : 10.3390/genes8100262 . ПМК 5664112 . ПМИД 28984839 .
- ^ Сирак К., Френандес Д., Новак М., Ван Гервен Д., Пинхаси Р. (2016). «Реферат Межконгресса IUAES 2016 - Сообщество разделено? Выявление генома(ов) сообщества средневекового Кулубнарти с помощью секвенирования следующего поколения» . Тезисы докладов Межконгресса IUAES 2016 . ИУАЭС: 115.
- ^ Перейти обратно: а б Рай Н., Тахер Н., Сингх М., Чауби Г., Джа Ан, Сингх Л., Тангарадж К. (январь 2014 г.). «Реликвия, раскопанная в западной Индии, вероятно, принадлежит грузинской царице Кетеван» Митохондрия 14 (1): 1–6 дои : 10.1016/j.myth.2013.12.002 . ПМИД 24355295 .
- ^ «Гаплогруппа U5» . haplogroup.org . 06.06.2016. Архивировано из оригинала 3 июля 2017 г. Проверено 13 июля 2017 г.
30 248,3 ± 5 330,5; ДИ=95% (Бехар и др., 2012)
- ^ Брандштеттер, Анита; Циммерманн, Беттина; Вагнер, Джанин; Гебель, Таня; Рёк, Александр В.; Салас, Антонио; Карраседо, Анхель; Парсон, Вальтер (4 июля 2008 г.). «Время и расшифровка изменчивости макрогаплогруппы R0 митохондриальной ДНК в Центральной Европе и на Ближнем Востоке» . Эволюционная биология BMC . 8 (1): 191. Бибкод : 2008BMCEE...8..191B . дои : 10.1186/1471-2148-8-191 . ISSN 1471-2148 . ПМЦ 2491632 . ПМИД 18601722 .
- ^ Соарес, Педро; Ахилли, Алессандро; Семино, Орнелла; Дэвис, Уильям; Маколей, Винсент; Бандельт, Ханс-Юрген; Торрони, Антонио; Ричардс, Мартин Б. (23 февраля 2010 г.). «Археогенетика Европы» . Современная биология . 20 (4): Р174–Р183. Бибкод : 2010CBio...20.R174S . дои : 10.1016/j.cub.2009.11.054 . ПМИД 20178764 .
- ^ Браманти Б., Томас М.Г., Хаак В., Унтерлендер М., Хорес П., Тамбетс К. и др. (октябрь 2009 г.). «Генетический разрыв между местными охотниками-собирателями и первыми земледельцами Центральной Европы» . Наука 326 (5949): 137–40. Бибкод : 2009Sci...326..137B . дои : 10.1126/science.1176869 . ПМИД 19729620 . S2CID 206521424 .
- ^ Перейти обратно: а б Мальмстрем Х., Гилберт М.Т., Томас М.Г., Брандстрем М., Стора Дж., Молнар П. и др. (ноябрь 2009 г.). «Древняя ДНК показывает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавами» . Современная биология . 19 (20): 1758–62. Бибкод : 2009CBio...19.1758M . дои : 10.1016/j.cub.2009.09.017 . ПМИД 19781941 . S2CID 9487217 .
- ^ Дегийу М.Ф., Солер Л., Пемонж М.Х., Скарр К., Жуссом Р., Лапорт Л. (январь 2011 г.). «Новости с запада: древняя ДНК из французской мегалитической погребальной камеры». Американский журнал физической антропологии . 144 (1): 108–18. дои : 10.1002/ajpa.21376 . ПМИД 20717990 .
- ^ Санчес-Куинто Ф., Шредер Х., Рамирес О., Авила-Аркос М.К., Пибус М., Олальде И. и др. (август 2012 г.). «Геномное сходство двух иберийских охотников-собирателей возрастом 7000 лет» (PDF) . Современная биология . 22 (16): 1494–9. Бибкод : 2012CBio...22.1494S . дои : 10.1016/j.cub.2012.06.005 . ПМИД 22748318 . S2CID 14913530 .
- ^ Лакан М., Кейзер С., Рико FX, Брукато Н., Таррус Дж., Бош А. и др. (ноябрь 2011 г.). «Древняя ДНК позволяет предположить ведущую роль, которую играли мужчины в распространении неолита» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (45): 18255–9. Бибкод : 2011PNAS..10818255L . дои : 10.1073/pnas.1113061108 . ПМК 3215063 . ПМИД 22042855 .
- ^ «Гаплогруппа U5» . Евпедия .
- ^ «Баскская генетика: Рефераты и резюме» . Кажария .
- ^ Перейти обратно: а б «Генографический проект» . Нэшнл Географик . Архивировано из оригинала 13 апреля 2005 года.
- ^ Перейти обратно: а б Хендриксон С.Л., Хатчесон Х.Б., Руис-Песини Е., Пул Дж.К., Лаутенбергер Дж., Сезгин Е. и др. (ноябрь 2008 г.). «Гаплогруппы митохондриальной ДНК влияют на прогрессирование СПИДа» . СПИД . 22 (18): 2429–39. дои : 10.1097/QAD.0b013e32831940bb . ПМЦ 2699618 . ПМИД 19005266 .
- ^ Антонио, Маргарет Л.; Гао, Зиюэ; М. Мутс, Ханна (2019). «Древний Рим: генетический перекресток Европы и Средиземноморья» . Наука . 366 (6466). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки (опубликовано 8 ноября 2019 г.): 708–714. Бибкод : 2019Sci...366..708A . дои : 10.1126/science.aay6826 . ПМК 7093155 . ПМИД 31699931 .
- ^ Возраст: 1398,1 ± 1872,3; ДИ=95% (Бехар и др., 2012) haplogroup.org [ постоянная мертвая ссылка ]
- ^ Паленсия-Мадрид Л, Кардосо С, Кейсер С, Лопес-Кинтана ХК, Генага-Лисасу А, де Панкорбо ММ (май 2017 г.). «Древние митохондриальные линии подтверждают доисторические материнские корни басков на севере Пиренейского полуострова» . Европейский журнал генетики человека . 25 (5): 631–636. дои : 10.1038/ejhg.2017.24 . ПМЦ 5437903 . ПМИД 28272540 .
- ^ 22794,0 ± 3590,3; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 11 октября 2017 г. на Wayback Machine.
- ^ Кудре С., Оливьери А., Ахилли А., Пала М., Мелхауи М., Шеркауи М. и др. (март 2009 г.). «Сложный и разнообразный митохондриальный генофонд берберского населения». Анналы генетики человека . 73 (2): 196–214. дои : 10.1111/j.1469-1809.2008.00493.x . ПМИД 19053990 . S2CID 21826485 .
- ^ Возраст: 15.529,7 ± 4.889,9; ДИ=95% «Гаплогруппа мтДНК U5b1» . haplogroup.org . 06.06.2016. Архивировано из оригинала 22 марта 2017 г. Проверено 13 июля 2017 г.
- ^ Роза А., Орнелас С., Джоблинг М.А., Брем А., Виллемс Р. (июль 2007 г.). «Y-хромосомное разнообразие населения Гвинеи-Бисау: многоэтническая перспектива» . Эволюционная биология BMC . 7 (1): 124. Бибкод : 2007BMCEE...7..124R . дои : 10.1186/1471-2148-7-124 . ЧВК 1976131 . ПМИД 17662131 .
- ^ Ахилли А., Ренго С., Батталья В., Пала М., Оливьери А., Форнарино С. и др. (май 2005 г.). «Саамы и берберы – неожиданная связь митохондриальной ДНК» . Американский журнал генетики человека . 76 (5): 883–6. дои : 10.1086/430073 . ПМК 1199377 . ПМИД 15791543 .
- ^ Эктор Д.З., Мария Н.К., Хуан М.К. (апрель 2017 г.). «В основном околосредиземноморское происхождение мтДНК Западной Евразии и Северной Африки в Пуэрто-Рико с сильным вкладом Канарских островов и Западной Африки». Биология человека . 89 (2): 125–155. doi : 10.13110/humanbiology.89.2.04 . ПМИД 29299964 . S2CID 21941155 .
- ^ Перейти обратно: а б с д Малярчук Б, Деренко М, Гжибовский Т, Перкова М, Рогалла У, Ванечек Т и др. (апрель 2010 г.). Гилберт (ред.). «Заселение Европы с точки зрения митохондриальной гаплогруппы U5» . ПЛОС ОДИН . 5 (4): е10285. Бибкод : 2010PLoSO...510285M . дои : 10.1371/journal.pone.0010285 . ПМК 2858207 . ПМИД 20422015 .
Субкластер U5b2a характеризуется преимущественно центральноевропейским распространением, поскольку в этот подкластер попадает большое количество образцов U5b из Польши, Словакии и Чехии. Например, субкластер U5b2a2 часто встречается в Центральной Европе (с наибольшей частотой его субкластера U5b2a2a1 у поляков) и датируется возникновением между 12–18 тыс. лет назад [...] Примечательно также, что внутри U5b2a1a субкластеру U5b2a1a1 предшествует средиземноморская ветвь. , что характерно для популяций Центральной и Восточной Европы (рис. 1). Другой субкластер U5b, U5b3, скорее всего, имеет свою родину на Итальянском полуострове, откуда он в голоцене распространился вдоль средиземноморского побережья [11]. Таким образом, в целом можно заключить, что первоначальная диверсификация U5b произошла в Южной и Центральной Европе, за которой последовало распространение определенных субкластеров U5b в Восточную Европу.
- ^ 20 040,2 ± 3 208,6; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 17 июня 2017 г. на Wayback Machine.
- ^ 14 938,0 ± 3 656,4; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 17 июня 2017 г. на Wayback Machine.
- ^ Возраст: 14 676,9 ± 2716,6; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 27 июля 2017 г. на Wayback Machine.
- ^ Меткалф, Том (14 декабря 2021 г.). «Самое раннее современное захоронение младенцев женского пола, найденное в Европе» . Нэшнл Географик . Архивировано из оригинала 14 декабря 2021 года . Проверено 16 декабря 2021 г.
- ^ Ходжкинс, Джейми (2021). «Младенческое захоронение в Арма-Вейране на северо-западе Италии дает представление о погребальных практиках и женской личности в Европе раннего мезолита» . Научные отчеты . 11 (1): 23735. Бибкод : 2021NatSR..1123735H . дои : 10.1038/s41598-021-02804-z . ПМЦ 8671481 . ПМИД 34907203 .
- ^ Возраст: 12 651,1 ± 5 668,2; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 28 декабря 2017 г. на Wayback Machine.
- ^ 4015,9 ± 3898,7; CI = 95% (Behar et al., 2012) haplogroup.org. Архивировано 6 сентября 2017 г. на Wayback Machine.
- ^ Матисоо-Смит Э.А., Гослинг А.Л., Букок Дж., Кардаильский О., Курумилиан Ю., Рудесли-Чебби С. и др. Матисоо-Смит Э.А., Гослинг А.Л., Букок Дж., Кардаильский О., Курумилиан Ю., Рудесли-Чебби С. и др. (25 мая 2016 г.). «Европейский митохондриальный гаплотип, выявленный в древних финикийских останках из Карфагена, Северная Африка» . ПЛОС ОДИН . 11 (5): e0155046. Бибкод : 2016PLoSO..1155046M . дои : 10.1371/journal.pone.0155046 . ПМК 4880306 . ПМИД 27224451 .
Образец молодого человека из Бирсы датируется концом VI века до нашей эры, и считается, что его происхождение отражает ранний поток генов из Иберии в Магриб.
- ^ Пала М., Ачилли А., Оливьери А., Хушиар Кашани Б., Перего У.А., Санна Д. и др. (июнь 2009 г.). «Митохондриальная гаплогруппа U5b3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев» (PDF) . Американский журнал генетики человека . 84 (6): 814–21. дои : 10.1016/j.ajhg.2009.05.004 . ПМК 2694970 . ПМИД 19500771 . Архивировано из оригинала (PDF) 22 июля 2018 г. Проверено 2 сентября 2009 г.
- ^ Фаррагут SJ (июнь 2017 г.). ГЕНЕТИЧЕСКОЕ НАСЛЕДИЕ ЕВРЕЕВ-СЕФАРДОВ: ОТЦОВСКАЯ И МАТЕРИНСКАЯ ЛИНИЯ НАСЕЛЕНИЯ ЧУЭТА (PDF) . Университет Балеарских островов (Диссертация).
- ^ Пала М., Ачилли А., Оливьери А., Хушиар Кашани Б., Перего У.А., Санна Д. и др. (июнь 2009 г.). «Митохондриальная гаплогруппа U5b3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев» (PDF) . Американский журнал генетики человека . 84 (6): 814–21. дои : 10.1016/j.ajhg.2009.05.004 . ПМК 2694970 . ПМИД 19500771 .
- ^ Софикару А., Добос А., Тринкаус Э. (ноябрь 2006 г.). «Ранние современные люди из Пестера Муиери, Бая-де-Фьер, Румыния» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (46): 17196–201. Бибкод : 2006PNAS..10317196S . дои : 10.1073/pnas.0608443103 . ПМЦ 1859909 . ПМИД 17085588 .
- ^ Хервелла М., Свенссон Э.М., Альберди А., Гюнтер Т., Изагирре Н., Мунтерс А.Р. и др. (май 2016 г.). «Митогеном 35 000-летнего человека разумного из Европы поддерживает палеолитическую обратную миграцию в Африку» . Научные отчеты . 6 : 25501. Бибкод : 2016NatSR...625501H . дои : 10.1038/srep25501 . ПМЦ 4872530 . ПМИД 27195518 . «Люди, несущие гаплогруппу U, возможно, распространились на запад из Западной Азии около 39–52 тыс. лет назад, достигнув Европы, о чем свидетельствует гаплогруппа U5, и Северной Африки, о чем свидетельствует гаплогруппа U6, что, вероятно, представляет собой генетический сигнал EUP-возвращения Homo sapiens из Евразии на Север. Африка Время появления самого недавнего общего предка (TMRCA) для U6 оценивается в 35,3 (24,6–46,4) тыс. лет назад [...] Наши оценки гаплогруппы U6 TMRCA, включающей древние геномы (включая PM1), определяют формирование. линии U6 обратно к 49,6 тыс. лет назад (95% HPD: 42–58 тыс. лет назад)»
- ^ Плаза С., Калафель Ф., Хелал А., Бузерна Н., Лефранк Дж., Бертранпети Дж., Комас Д. (июль 2003 г.). «Присоединение к Геркулесовым столпам: последовательности мтДНК демонстрируют разнонаправленный поток генов в западном Средиземноморье» (PDF) . Анналы генетики человека . 67 (Часть 4): 312–28. дои : 10.1046/j.1469-1809.2003.00039.x . ПМИД 12914566 . S2CID 11201992 .
- ^ Перейти обратно: а б с Мака-Мейер Н., Гонсалес А.М., Пестано Х., Флорес К., Ларруга Х.М., Кабрера В.М. (октябрь 2003 г.). «Транзит митохондриальной ДНК между Западной Азией и Северной Африкой, сделанный на основе филогеографии U6» . БМК Генетика . 4 (1): 15. дои : 10.1186/1471-2156-4-15 . ПМК 270091 . ПМИД 14563219 .
- ^ Сересо М., Черный В., Карраседо А., Салас А. (апрель 2011 г.). «Новые данные о структуре населения бассейна озера Чад, полученные с помощью высокопроизводительного генотипирования митохондриальной ДНК, кодирующей SNP» . ПЛОС ОДИН . 6 (4): e18682. Бибкод : 2011PLoSO...618682C . CiteSeerX 10.1.1.291.8871 . дои : 10.1371/journal.pone.0018682 . ПМК 3080428 . ПМИД 21533064 .
- ^ Перейти обратно: а б Сечер Б., Фрегель Р., Ларруга Дж.М., Кабрера В.М., Эндикотт П., Пестано Дж.Дж. и др. (май 2014 г.). «История потока гена гаплогруппы U6 североафриканской митохондриальной ДНК на африканский, евразийский и американский континенты» . Эволюционная биология BMC . 14 (1): 109. Бибкод : 2014BMCEE..14..109S . дои : 10.1186/1471-2148-14-109 . ПМК 4062890 . ПМИД 24885141 .
- ^ Эрнандес К.Л., Соарес П., Дюгужон Х.М., Новеллетто А., Родригес Х.Н., Рито Т. и др. (2015). «Ранние голоценовые и исторические африканские признаки мтДНК на Пиренейском полуострове: Андалузский регион как парадигма» . ПЛОС ОДИН . 10 (10): e0139784. Бибкод : 2015PLoSO..1039784H . дои : 10.1371/journal.pone.0139784 . ПМЦ 4624789 . ПМИД 26509580 .
- ^ ван де Лоосдрехт М., Бузуггар А., Хамфри Л., Пост С., Бартон Н., Аксиму-Петри А. и др. (май 2018 г.). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ПМИД 29545507 .
- ^ Фрегель Р., Мендес Ф.Л., Бокбот Ю., Мартин-Сокас Д., Камалич-Массие, доктор медицинских наук, Сантана Дж. и др. (июнь 2018 г.). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (26): 6774–6779. Бибкод : 2018PNAS..115.6774F . дои : 10.1073/pnas.1800851115 . ПМК 6042094 . ПМИД 29895688 .
- ^ Мохамед Х.И. Генетические закономерности вариаций Y-хромосомы и митохондриальной ДНК, имеющие значение для заселения Судана (докторская диссертация). Хартумский университет. Архивировано из оригинала (PDF) 10 ноября 2020 года . Проверено 17 апреля 2016 г.
- ^ Сикора, Мартин; и др. (2019). «История населения северо-восточной Сибири со времен плейстоцена» (PDF) . Природа . 570 (7760): 182–188. Бибкод : 2019Natur.570..182S . bioRxiv 10.1101/448829 . дои : 10.1038/s41586-019-1279-z . ПМИД 31168093 . S2CID 174809069 .
- ^ Перейти обратно: а б с д и ж г Мецпалу М., Кивисилд Т., Мецпалу Е., Парик Дж., Худжашов Г., Калдма К. и др. (август 2004 г.). «Большая часть существующих границ мтДНК на юге и юго-западе Азии, вероятно, сформировалась во время первоначального заселения Евразии анатомически современными людьми» . БМК Генетика . 5:26 . дои : 10.1186/1471-2156-5-26 . ПМК 516768 . ПМИД 15339343 .
- ^ Перейти обратно: а б Маджи С., Критика С., Васулу Т.С. (март 2008 г.). «Распространение макрогаплогруппы N митохондриальной ДНК в Индии с особым акцентом на гаплогруппу R и ее субгаплогруппу U». (PDF) . Международный журнал генетики человека . 8 (1–2): 85–96. дои : 10.1080/09723757.2008.11886022 . S2CID 14231815 .
- ^ Шинде, В; Нарасимхан, В.М.; Роланд, Н.; Маллик, С; Мах, М; Липсон, М; Накацука, Н; Адамски, Н; Бруманд Хошбахт, Н.; Ферри, М; Лоусон, AM; Мишель, М; Оппенгеймер, Дж.; Стюардсон, К.; Джадхав, Н.; Ким, YJ; Чаттерджи, М; Мунши, А; Паньям, А; Вагмаре, П; Ядав, Ю; Патель, Х; Кошик, А; Тангарадж, К; Мейер, М; Паттерсон, Н.; Дождь; Райх, Д. (17 октября 2019 г.). «Древний хараппский геном не имеет происхождения от степных скотоводов или иранских фермеров» (PDF) . Ячейка 179 (3): 729–735.e10. дои : 10.1016/j.cell.2019.08.048 . ПМК 6800651 . ПМИД 31495572 .
- ^ Бермишева М.А., Кутуев И.А., Коршунова Т.И., Дубова Н.А., Виллемс Р., Хуснутдинова Е.К. (2004). «[Филогеографический анализ митохондриальной ДНК ногайцев: высокий уровень смешения материнских линий Восточной и Западной Евразии]». Молекулярная биология . 38 (4): 617–24. ПМИД 15456133 .
- ^ Перейти обратно: а б Дербенева О.А., Стариковская Е.Б., Володько Н.В., Уоллес Д.С., Сукерник Р.И. (ноябрь 2002 г.). «[Вариации митохондриальной ДНК у кетов и нганасан и ранних народов Северной Евразии] Генетика . 38 (11): 1554–60. ПМИД 12500682 .
- ^ Скотт Р. (2009). «Филогения mt-hg U2a» (PDF) .
- ^ Скотт Р. (2010). «Филогения mt-hg U2b» (PDF) .
- ^ Скотт Р. (2009). «Филогения mt-hg U2c» (PDF) .
- ^ Скотт Р. (2009). «Филогения mt-hg U2d» (PDF) .
- ^ Скотт Р. (2010). «Филогения mt-hg U2e» (PDF) .
- ^ Малярчук Б, Деренко М, Перкова М, Ванечек Т (октябрь 2008 г.). «Филогения и распространение митохондриальной гаплогруппы U2d». Биология человека . 80 (5): 565–71. дои : 10.3378/1534-6617-80.5.565 . ПМИД 19341323 . S2CID 9447306 .
- ^ А. Сеген-Орландо; и др. (6 ноября 2014 г.). «Геномная структура европейцев насчитывает не менее 36 200 лет» . Наука . 346 (6213): 1113–1118. Бибкод : 2014Sci...346.1113S . дои : 10.1126/science.aaa0114 . ПМИД 25378462 . S2CID 206632421 .
- ^ «Анализ ДНК ранних европейцев» . Новости Би-би-си . 2010.
- ^ Ли Э.Дж., Макаревич С., Реннеберг Р., Хардер М., Краузе-Кёра Б., Мюллер С., Остриц С., Ферен-Шмитц Л., Шрайбер С., Мюллер Дж., фон Вурмб-Шварк Н., Небель А. (август 2012 г.). «Новые генетические модели европейского неолита: перспективы захоронения колокольчиков позднего неолита в Германии» . Американский журнал физической антропологии . 148 (4): 571–9. дои : 10.1002/ajpa.22074 . ПМИД 22552938 .
- ^ Мельхиор Л., Гилберт М.Т., Кивисилд Т., Линнеруп Н., Диссинг Дж. (февраль 2008 г.). «Редкие гаплогруппы мтДНК и генетические различия в богатых и бедных датских деревнях железного века». Американский журнал физической антропологии . 135 (2): 206–15. дои : 10.1002/ajpa.20721 . ПМИД 18046774 .
- ^ Ким К., Бреннер Ч., Майр В.Х., Ли К.Х., Ким Дж.Х., Гелегдорж Э. и др. (июль 2010 г.). «Западно-евразийский мужчина найден на 2000-летнем элитном кладбище хунну на северо-востоке Монголии» (PDF) . Американский журнал физической антропологии . 142 (3): 429–40. дои : 10.1002/ajpa.21242 . ПМИД 20091844 .
- ^ Перейти обратно: а б Скотт Р. (2010). «Филогения mt-hg U3» (PDF) .
- ^ Деренко М., Малярчук Б., Бахманимер А., Денисова Г., Перкова М., Фарджадян С. и др. (2013). «Полное разнообразие митохондриальной ДНК у иранцев» . ПЛОС ОДИН . 8 (11): е80673. Бибкод : 2013PLoSO...880673D . дои : 10.1371/journal.pone.0080673 . ПМЦ 3828245 . ПМИД 24244704 .
- ^ Перейти обратно: а б Ричардс М., Маколей В., Хики Е., Вега Е., Сайкс Б., Гуида В. и др. (ноябрь 2000 г.). «Отслеживание линий европейских основателей в пуле мтДНК Ближнего Востока» (PDF) . Американский журнал генетики человека . 67 (5): 1251–76. дои : 10.1016/S0002-9297(07)62954-1 . ПМК 1288566 . ПМИД 11032788 . Архивировано из оригинала (PDF) 4 марта 2009 г. Проверено 19 марта 2007 г.
- ^ Перейти обратно: а б Кинтана-Мурси Л., Чейкс Р., Уэллс Р.С., Бехар Д.М., Саяр Х., Скозцари Р. и др. (май 2004 г.). «Там, где запад встречается с востоком: сложный ландшафт мтДНК юго-западного и центральноазиатского коридора» . Американский журнал генетики человека . 74 (5): 827–45. дои : 10.1086/383236 . ПМК 1181978 . ПМИД 15077202 .
- ^ Малярчук Б.А., Гжибовский Т, Деренко М.В., Чарный Ю, Мисцицка-Сливка Д (март 2006 г.). «Разнообразие митохондриальной ДНК у польских цыган». Анналы генетики человека . 70 (Часть 2): 195–206. дои : 10.1111/j.1529-8817.2005.00222.x . ПМИД 16626330 . S2CID 662278 .
- ^ Мендисабаль И., Валенте С., Гужман А., Алвеш С., Гомеш В., Гойос А. и др. (январь 2011 г.). «Реконструкция индийского происхождения и расселения европейских цыган: взгляд на материнскую генетику» . ПЛОС ОДИН . 6 (1): e15988. Бибкод : 2011PLoSO...615988M . дои : 10.1371/journal.pone.0015988 . ПМК 3018485 . ПМИД 21264345 .
- ^ Куянова М, Перейра Л, Фернандес В, Перейра ЖБ, Черны В (октябрь 2009 г.). «Генетический вклад ближневосточного неолита в небольшой оазис Западной египетской пустыни». Американский журнал физической антропологии . 140 (2): 336–46. дои : 10.1002/ajpa.21078 . ПМИД 19425100 .
- ^ Стеванович А., Жиль А., Бузед Э., Кефи Р., Пэрис Ф., Гайро Р.П. и др. (январь 2004 г.). «Разнообразие последовательностей митохондриальной ДНК у малоподвижного населения Египта». Анналы генетики человека . 68 (Часть 1): 23–39. дои : 10.1046/j.1529-8817.2003.00057.x . PMID 14748828 . S2CID 44901197 .
- ^ Перейра Л., Черны В., Сересо М., Сильва Н.М., Гаек М., Васикова А. и др. (август 2010 г.). «Связывание генофондов стран южнее Сахары и Западной Евразии: материнское и отцовское наследие кочевников-туарегов из африканского Сахеля» . Европейский журнал генетики человека . 18 (8): 915–23. дои : 10.1038/ejhg.2010.21 . ПМЦ 2987384 . ПМИД 20234393 .
- ^ «Судебно-медицинская и филогеографическая характеристика линий мтДНК из Сомали» .
- ^ Перейти обратно: а б Саламон М., Цур С., Аренсбург Б., Зиас Дж., Нагар Ю., Вайнер С., Боаретто Э. (январь 2010 г.). «Древние последовательности мтдны и радиоуглеродное датирование человеческих костей из энеолитических пещер Вади-эль-Маккух» (PDF) . Средиземноморская археология и археометрия . 10 (2): 1–4. Бибкод : 2010MAA....10....1S . Архивировано из оригинала (PDF) 03 марта 2016 г.
- ^ Буджа М (декабрь 2013 г.). «Неолитические горшки и гончары в Европе: конец миграционной модели« демического распространения »» . Документа Преисторика . 40 : 38–56. дои : 10.4312/dp.40.5 .
- ^ Дер Саркисян С, Балановский О, Брандт Г, Хартанович В, Бужилова А, Кошель С и др. (2013). «Древняя ДНК раскрывает доисторический поток генов из Сибири в сложной истории человеческого населения Северо-Восточной Европы» . ПЛОС Генетика . 9 (2): e1003296. дои : 10.1371/journal.pgen.1003296 . ПМЦ 3573127 . ПМИД 23459685 .
- ^ Хаак В., Балановский О., Санчес Дж.Дж., Кошель С., Запорожченко В., Адлер С.Дж. и др. (ноябрь 2010 г.). «Древняя ДНК европейских фермеров раннего неолита раскрывает их ближневосточное родство» . ПЛОС Биология . 8 (11): е1000536. дои : 10.1371/journal.pbio.1000536 . ПМЦ 2976717 . ПМИД 21085689 .
- ^ Перейти обратно: а б с Дербенева О.А., Стариковская Е.Б., Уоллес Д.С., Сукерник Р.И. (апрель 2002 г.). «Следы ранних евразийцев у манси Северо-Западной Сибири, выявленные методом анализа митохондриальной ДНК» . Американский журнал генетики человека . 70 (4): 1009–14. дои : 10.1086/339524 . ПМК 379094 . ПМИД 11845409 .
- ^ Володько Н.В., Стариковская Е.Б., Мазунин И.О., Ельцов Н.П., Найденко П.В., Уоллес Д.С. и др. (май 2008 г.). «Разнообразие митохондриального генома арктических сибиряков с особым упором на эволюционную историю Берингии и плейстоценового заселения Америки» . Американский журнал генетики человека . 82 (5): 1084–100. дои : 10.1016/j.ajhg.2008.03.019 . ПМК 2427195 . ПМИД 18452887 .
- ^ «Клан Ульрики» . Материнское происхождение . Оксфордские предки. 2013. Архивировано из оригинала 15 июля 2017 г. Проверено 7 февраля 2013 г.
- ^ «Материнское происхождение» . Оксфорд Предки Лтд . Архивировано из оригинала 15 июля 2017 г. Проверено 7 февраля 2013 г.
- ^ Башир М (2012). «Сохранение наследия калашей» . Пакистан сегодня .
- ^ Логан I (2014). «Последовательности генбанка U4a1» .
- ^ «Калаш» . Блог, посвященный гаплогруппе U4 .
- ^ Скотт Р. (2011). «Филогения mt-hg U4a» (PDF) .
- ^ Скотт Р. (2011). «Филогения mt-hg U4b» (PDF) .
- ^ Скотт Р. (2011). «Филогения mt-hg U4c» (PDF) .
- ^ Скотт Р. (2010). «Филогения mt-hg U4d» (PDF) .
- ^ «Доисторическая европейская ДНК - частоты гаплогрупп мтДНК и Y-ДНК по периодам» . Евпедия .
- ^ Перейти обратно: а б Мельхиор Л., Линнеруп Н., Зигизмунд Х.Р., Кивисилд Т., Диссинг Дж. (июль 2010 г.). «Генетическое разнообразие древних скандинавских популяций» . ПЛОС ОДИН . 5 (7): e11898. Бибкод : 2010PLoSO...511898M . дои : 10.1371/journal.pone.0011898 . ПМЦ 2912848 . ПМИД 20689597 .
- ^ Малярчук Б, Гжибовский Т, Деренко М, Перкова М, Ванечек Т, Лазур Дж, Гомолчак П, Цыбовский И (август 2008 г.). «Филогения митохондриальной ДНК у восточных и западных славян» . Молекулярная биология и эволюция . 25 (8): 1651–8. дои : 10.1093/molbev/msn114 . ПМИД 18477584 .
- ^ Манко Дж (2009). «Древняя западноевразийская ДНК медного и бронзового веков» . Путешествия предков . Архивировано из оригинала 22 января 2017 г.
- ^ Скотт Р. (2010). «Полный список мутаций hg U» (PDF) .
- ^ «Славянизация уральцев Волго-Окского междуречья. Языки и гены от железного века до славян» . 22 января 2023 г. Проверено 31 августа 2023 г. - через YouTube .
- ^ «Женщины Осеберга и мужчина Гокстада сохраняют свои генетические тайны» (PDF) . Культурно-исторический музей . Университет Осло. [ постоянная мертвая ссылка ]
- ^ Перейти обратно: а б «Проект митохондриальной ДНК гаплогруппы U7» . Архивировано из оригинала 2 июня 2014 года.
- ^ Перейти обратно: а б Рудбек Л., Гилберт М.Т., Виллерслев Э., Хансен А.Дж., Линнеруп Н., Кристенсен Т., Диссинг Дж. (октябрь 2005 г.). «Анализ мтДНК человеческих останков с раннего датского христианского кладбища». Американский журнал физической антропологии . 128 (2): 424–9. дои : 10.1002/ajpa.20294 . ПМИД 15838837 .
- ^ Ранавира Л., Каевсутти С., Вин Тун А., Буньярит Х., Пулсуван С., Лертрит П. (январь 2014 г.). «История митохондриальной ДНК этнических народов Шри-Ланки: их отношения внутри острова и с индийским субконтинентальным населением» . Журнал генетики человека . 59 (1): 28–36. дои : 10.1038/jhg.2013.112 . ПМИД 24196378 . S2CID 41185629 .
- ^ Перейти обратно: а б Скотт Р. (2010). «Филогения mt-hg U7» (PDF) .
- ^ Ли Э.Дж., Штеффен С., Хардер М., Краузе-Кёра Б., Фон Вурмб-Шварк Н., Небель А. (январь 2012 г.). «Древний взгляд на ДНК «княжеских захоронений» железного века из Баден-Вюртемберга, Германия» . Американский журнал физической антропологии . 147 : 190. дои : 10.1002/ajpa.22033 .
- ^ Берглунд Н. (26 марта 2007 г.). «Женщина-викинг имела корни у Черного моря –» . Aftenposten — Новости на английском языке . Aftenposten.no. Архивировано из оригинала 26 августа 2010 г. Проверено 11 апреля 2010 г.
- ^ Фаррелл Дж.Дж., Аль-Али А.К., Фаррер Л.А., Аль-Нафаи А.Н., Аль-Рубайш А.М., Мелиста Э. и др. «Геном Саудовской Аравии свидетельствует о двухэтапной миграции из Африки» . Американское общество генетики человека (ASHG). Архивировано из оригинала 13 сентября 2016 г. Проверено 12 мая 2016 г.
- ^ Кивисилд Т., Рейдла М., Метспалу Е., Роза А., Брем А., Пеннарун Е. и др. (ноябрь 2004 г.). «Наследие эфиопской митохондриальной ДНК: отслеживание потока генов через ворота слез и вокруг них» . Американский журнал генетики человека . 75 (5): 752–70. дои : 10.1086/425161 . ПМЦ 1182106 . ПМИД 15457403 .
- ^ Найдите свои корни . Сезон 1. Эпизод 8. 06.05.2012. ПБС.
- ^ Надин Эпштейн (сентябрь – октябрь 2012 г.). «Великий эксперимент с ДНК журнала Moment» . Журнал «Момент» . п. 45 . Проверено 7 марта 2024 г.
- ^ Мур, СиСи (31 декабря 2010 г.). «Моя полная митохондриальная последовательность, гетероплазмия и сравнение обозначений мтДНК на FTDNA и 23andMe» . Ваш генетический генеалог .
Дальнейшее чтение
[ редактировать ]- Бехар Д.М., ван Овен М., Россет С., Метспалу М., Лоогвяли Э.Л., Сильва Н.М., Кивисилд Т., Торрони А., Виллемс Р. (апрель 2012 г.). «Коперниканская» переоценка дерева митохондриальной ДНК человека от его корня» . Американский журнал генетики человека . 90 (4): 675–84. дои : 10.1016/j.ajhg.2012.03.002 . ПМЦ 3322232 . ПМИД 22482806 .
Внешние ссылки
[ редактировать ]- Общий
- Яна Логана Сайт митохондриальной ДНК
- Манниса ван Овена Филодерево
- Гаплогруппа U
- YFull MTree Гаплогруппа U
- MITOMAP Гаплогруппа U
- Распространение гаплогруппы U , от National Geographic
- Проект U1 мтДНК в ДНК Семейного древа
- Проект гаплогруппы мтДНК U3 в ДНК генеалогического древа
- Проект гаплогруппы мтДНК U4 в генеалогическом древе ДНК
- Проект мтДНК U5 в ДНК Семейного древа
- Проект мтДНК U6 в ДНК Семейного древа
- Гаплогруппа мтДНК U7 в ДНК генеалогического древа
- Гаплогруппа мтДНК U8 в ДНК генеалогического древа
- Проект мтДНК U9 в ДНК Семейного древа