Предуравральные гены
Предуравральные гены кодируют транскрипционные факторы базового класса спиральной спирали (BHLH), которые отвечают за развитие нейроэктодермальных клеток-предшественников. Предуравральные гены имеют множество функций в развитии нервного нерва . Они интегрируют позиционную информацию и вносят свой вклад в спецификацию идентичности предшественников. [ 1 ] Из тех же типов эктодермальных клеток нервные или эпидермальные клетки могут развиваться на основе взаимодействия между пронуравральными и нейрогенными генами. [ 2 ] Нейрогенные гены называются так, потому что потеря функциональных мутантов показывает увеличение числа развитых нейронных предшественников. С другой стороны, у мутантов -у мутантов -предшественных генов не могут быть развиты нейронные клетки предшественников. [ 3 ]
Проневральные гены экспрессируются в группах клеток (пронейральные кластеры), из которых одна клетка -предшественника - обычно в середине - будет выделяться, что приводит к образованию многих различных типов нейронов в центральной и периферической нервной системах . [ 4 ] [ 5 ] Проневральные гены кодируют группу белков BHLH, которые играют важную роль в контроле клеточной судьбы в различных типах тканей. [ 6 ] Основные белки спиральной спирали-спирали характеризуются двумя альфа-спиралами, разделенными петлей. Спирали опосредуют димеризацию , и прилегающая базовая область необходима для связывания ДНК . [ 1 ] Геном человека содержит приблизительно 125 факторов BHLH. [ 7 ]
Открытие
[ редактировать ]Проневральные гены были впервые идентифицированы в 1920 -х годах [ кем? ] Когда мутанты , у которых не хватало подмножествами внешних органов или щетины , были обнаружены. [ 8 ] Позже, в 1970-х годах, комплекс Achaete-Scute , комплекс генов, которые участвуют в регулировании ранних этапов развития нейронного развития у дрозофилы , были идентифицированы [ кем? ] . [ 9 ] Используя молекулярные инструменты, можно было выделить первые четыре гена этого комплекса: ахате (AC) , Scute (SC) , смертельный из Scute (LSC) и Asense (ASE) . [ 9 ] Еще один поневральный ген, атональный (ATO) [ 10 ] был изолирован недавно [ когда? ] и два гена, связанных с ATO, Amos и Cato, были позже изолированы, определяя второе семейство пронуравральных генов-атональный комплекс. [ 11 ] Недавно [ когда? ] Первым гомологом генов пронераврала мух, который можно было бы найти у млекопитающих, был Mash1 . [ 4 ]
СПИСОК ПРОНУРАЛЬНЫХ Генов
[ редактировать ]Этот список относится к белкам BHLH , обнаруженным у беспозвоночных и позвоночных . Они сгруппированы в отдельных семействах на основе более тесных сходств последовательностей в домене BHLH: [ 1 ]
| Организмы | Электронные белковые | Атональная семья | Семья НАТО | Семья Олиго | Семейство нейродов | нейрогенинов Семейство | Ахат-Семья | Семья NSCL |
|---|---|---|---|---|---|---|---|---|
| Беспозвоночные | Без дочери | Atonal, Amos, Cato, Lin32 | НАТО3 | Двупарный | CNASH, Achaete (AC) , Asense (ASE) , Scute (SC) , смертельный из Scute (LSC) | |||
| позвоночные | E-12 | Math1, Math5 | НАТО3 | Beta3, Beta4, Olig1 , Olig2 , Olig3 | NeuroD1 (NeuroD) , NeuroD2 (NDRF) , NeuroD4 (Math3), NeuroD6 (Math2, Nex1) | amphioxngn, ngn1 , ngn2 , ngn3 | Cash4, Mash1 , Mash2, Hash3 | NSCL1, NSCL2 |
Предуравральные гены функционируют
[ редактировать ]Гены семейств ASC и нейрогенинов , и, возможно, члены семейства гомологов ATO, имеют сходную проннуральную функцию у позвоночных с функционированием своих коллег -дрозофилов , тогда как другие нейронные гены BHLH участвуют в определении существенных существа или в дифференцировке нейронов, но имеют Нет предстерарной роли. [ 1 ]
Нейронные функции
[ редактировать ]Проневральные белки связывают ДНК в виде гетеродимерных комплексов, которые образуются белками BHLH или E -белками. Поскольку гетеродимеризация является предпосылкой для связывания ДНК , факторы, которые мешают димеризации, эффективно действуют как пассивные репрессоры активности генов. Предуравральные белки специально связывают последовательности ДНК , которые содержат сердечный гексануклеотидный мотив, Canntg, известный как E-Box . Основная область и спираль 1 домена BHLH образуют длинную альфа-спираль , которая связана с областью петли с спиралью 2. Прямые контакты между BHLH остатками и ДНК отвечают за общую способность нейронных белков BHLH связываться с сердечником E -Кабоба . [ 1 ] Клетки внутри кластера, которые экспрессируют пронуравральный ген (называемый предуравральным кластером), можно рассматривать как клетки группы эквивалентности. В пределах предтераврального кластера клетки конкурируют друг с другом, так что только подмножество клеток выделяется для развития в предшественниках нейронов. Этот процесс выявления опосредуется клеточными взаимодействиями, интерпретируемыми в результате действия нейрогенных генов. В нейроэктодерме нейрогенные гены необходимы для выделения клеток из пронейральных кластеров для образования предшественников нейронов, в результате чего оставшиеся клетки пронейральных кластеров развиваются в эпидермальные клетки. [ 12 ] Предуравральные гены могут функционировать в аналогичной моде у позвоночных и беспозвоночных , в частности, они были вовлечены в ранний нейрогенез . [ 13 ] Хотя предуравральные белки отвечают за триггер -нейрогенез необходимы различные белки нейронных и глиальных клеток , для различных типов . Это подразумевает, что каждый из этих белков способен регулировать как общие гены -мишени для нейрогенеза , так и уникальные гены -мишени для характеристик нейронального подтипа. [ 14 ] Пронеравральные BHLH факторы транскрипции , не только способствуют нейрогенезу путем активации экспрессии каскада нейрональных генов, но они ингибируют экспрессию глиальных генов. [ 15 ] Нейронные гены BHLH имеют разные функции в зависимости от: чувствительности к латеральному ингибированию , которое определяет, становится ли клетка эпидермальной или нейронной, и является ли ген экспрессируется в ЦНС до или после терминального митоза . [ 4 ]
Проневральные гены способствуют нейрогенезу и ингибируют глиогенез , но некоторые нейрогенные факторы могут регулировать оба этих процесса, в зависимости от концентрации пронуравральных генов. Например, BMP (кости морфогенетические белки) способствуют нейрогенезу у предшественников, которые экспрессируют высокие уровни нейрогена-1 и глиогенез у предшественников, которые экспрессируют низкие уровни нейрогена-1 . [ 5 ] Процессы глиогенеза зависят от низких концентраций или делубения пронуравральных генов и могут быть ускорены в зависимости от того, какие гены затрагивают пронуравральные гены. [ 16 ]
В беспозвоночных
[ редактировать ]У Drosophila гены проневрала сначала экспрессируются в покоящихся эктодермальных клетках, которые имеют как эпидермальный, так и нейрональный потенциал. Проневральная активность приводит к выбору предшественников, которые привержены нейронной судьбе, но остаются мультипотентными , причем прарогенщики органа чувствительности вызывают нейроны , глию и другие не-нейрональные типы клеток. Кроме того, некоторые нейробласты центральной нервной системы также генерируют как нейроны , так и глию . Предшественники периферической и центральной нервной системы начинают делиться только после того, как экспрессия гена пронураврала утихла. [ 1 ]
У позвоночных
[ редактировать ]Проневральные гены сначала экспрессируются в нейроэпителиальных клетках, которые уже указаны для нервной судьбы и являются самообновлением. Проневральная активность приводит к генерации и расслаиванию предшественников, которые ограничены судьбой нейронов и имеют ограниченный митотический потенциал. В некоторых линиях, по крайней мере, проневральные гены участвуют в приверженности нейронных предшественников к судьбе нейронов за счет глиальной судьбы. [ 1 ]
В процессе бокового ингибирования
[ редактировать ]
Боковое ингибирование -это клеточное взаимодействие , которое происходит в группе проннурации, чтобы определить и ограничить клетки, которые вызывают нейробласту . [ 16 ] Во время этого взаимодействия зарождающиеся нейробласты экспрессируют пронуравральные гены выше определенного порога и, в то же время, они экспрессируют мембранный лиганд, называемый «дельта», который связывает и активирует рецепторы Notch, экспрессируемые в соседних клетках. Как только Notch активируется, активность проневральных генов уменьшается в этих клетках, вероятно, из -за активации генов в «энхансере комплекса разделения E (SPL)», кодируя в ингибирующих BHLH факторах транскрипции . [ 17 ] При ингибировании пронуравральные гены предотвращают нервные клетки, но также снижают их уровни «дельта». Эти конкретные взаимодействия ограничивают проннуральную активность одной клеткой в каждой проннуральной кластере, что приводит к ростам паттерна соли и перца. [ 2 ] [ 16 ]
Не все пронивральные гены одинаково чувствительны к боковому ингибированию . Например, в Xenopus Chitnis и Kintner продемонстрировали, что «Xash-3» и Neurod ( комплекс Achaete-Scute ) по-разному реагируют на латеральное ингибирование , что отражает различную способность активировать гены-мишени и дифференциальную восприимчивость этих генов-мишеней к репрессии нотч [ 16 ] Задние исследования показали, что даже когда сигнальный путь Notch/Delta блокируется, Wnt2b способен ингибировать дифференцировку нейронов посредством подавления экспрессии мРНК множественных проневральных генов, а также Notch1 . С помощью этого механизма Wnt2b поддерживает клетки -предшественники, недифференцированные путем ослабления экспрессии пронуравральных и нейрогенных генов, предотвращая попадание клеток в каскад дифференцировки, регулируемую пронурранными генами и Notch . [ 18 ] Хотя передача сигналов Notch участвует в контроле экспрессии генов проннуральной школы, петли с положительной обратной обратной обратной обратной обратной обратной обратной обратнойцей для увеличения или поддержания уровней пронуравральных генов необходимы . Факторы транскрипции, ответственные за это поддержание, могут действовать посредством ингибирования сигнального пути Notch в конкретных клетках или на посттранскрипционном уровне, влияя на транскрипцию и функцию проневральных генов. [ 1 ]
В нейрогенезе
[ редактировать ]Нейрогенез в нервной системе беспозвоночных
[ редактировать ]У беспозвоночных генов проневрала, в частности, члены комплекса ахэте-дисков (AS-C) способствуют нейрогенезу , в то время как нейрогенные гены предотвращают нейрогенез и облегчают эпидермальное развитие . The formation of neuroblasts depends on the Achaete-scute complex genes – achaete (ac) , scute (sc) , lethal of scute (lsc) and ventral nervous system defective (''vnd''). Однако только «VND» может контролировать этот процесс формирования, потому что этот ген активирует экспрессию других. [ 19 ] [ 20 ] Факторы AC , SC , LSC изначально экспрессируются в перчарте эмбриональной центральной нервной системы ( нейроэктодерма ) в пронейральных кластерах, из которых отдельные нейробласты . возникают [ 20 ] Каждая клетка пронурранного кластера имеет общий потенциал нейробластам. Локальное ингибирование оставшихся клеток путем увеличивающихся нейробластов гарантирует, что только один нейробласт возникает из -за проннуральной кластера. Все клетки кластера сохраняют свой потенциал формирования NB, по крайней мере, когда NB увеличивается, но теряет этот потенциал к тому времени, когда ячейка собирается разделить. Паттерны экспрессии генов проневрала приводят к различным способам образования нейробластов в голове и стволе. Коэкспрессия проннуральных генов в мозга нейробластах является временной и варьируется в зависимости от стадии развития. [ 21 ]
Нейрогенез утоматогастральной нервной системы у дрозофилы
[ редактировать ]Экспрессия генов в проневральных генах в нейроэктодермальных клетках, которые составляют пронуравральные кластеры, становится компетентным для расслаивания в качестве нейробластов. Хотя нейробласты являются предшественниками Drosophila центральной нервной системы (CNS), экспрессия пронураврала гена также участвует в контрольной спецификации и морфогенезе предшественников устьил -нервных клеток. Эти гены экспрессируются и требуются на всех этапах развития устьительной нервной системы (SNS), чтобы регулировать количество, паттерн и структурные характеристики субпопуляций SNS. Правильный баланс между пронуравральным и нейрогенным экспрессией генов на плакодах SNS участвует в контроле сложной последовательности морфогенетических движений (расслаивание, инвагинация и диссоциация), с помощью которой эти плакаты вызывают различные субпопуляции SNS . [ 22 ]
Нейрогенез в центральной нервной системе позвоночных
[ редактировать ]В центральной нервной системе не все гены BHLH участвуют в нейрогенезе, потому что семейства Neurod и '' Math3/NeuRom 'также участвуют в решении о судьбе нейронал против мрлиальной клетки. Другая проневральная семья (которая включает в себя «Math1» и «Math5») имеет важное значение для развития небольшого числа нейронных линий, тогда как «Math1» также играет роль в идентичности интернейрона спецификации . Типы клеток, которые зависят от экспрессии «математика», относятся к проприоцептивному сенсорному пути . [ 1 ] Bertrand et al. (2002) подтвердили проннуральную активность «MASH1», NGN1 и NGN2 и, возможно, Math1 и «Math5» в мышке. [ 1 ] Нейрогенез в центральной нервной системе зависит от ингибирования гена пронураврала путем сигнального пути Notch , а отсутствие этого ключевого регулятора приводит к преждевременной дифференцировке нейронов . Для поддержания клеток нервных предшественников между соседними клетками происходит регуляторная петля, которая включает в себя боковый процесс ингибирования (см. Боковое ингибирование). [ 23 ] В отсутствие бокового ингибирования некоторые проннуральные гены, такие как ASCL1 или «нейрога», способны индуцировать экспрессию нейрон-специфических генов, приводящих к преждевременному образованию раннего родового нейронов. [ 24 ] Ratié и коллеги (2013) включали в себя то, что сеть генов Notch предзнаменования играет важную роль в обновлении и переходе на клеточную судьбу и переходу на мыши. [ 23 ]
Нейрогенез в периферической нервной системе позвоночных
[ редактировать ]В периферической нервной системе NGN участвуют в определении всех черепных и позвоночных сенсорных предшественников. Проневральные гены, такие как MASH1 , NGN1 и NGN2, в основном экспрессируются в большинстве предшественников спинного мозга , а также совместны в дорсальном телефонах . Вместе эти группы факторов BHLH способствуют генерации всех предшественников коры головного мозга . MASH1 - единственный ген, экспрессируемый в вентральном домене . Однако в вентральных и дорсальных концах нейронной трубки экспрессируется другой тип пронивраленных генов, таких как NGN3 и «Math1». [ 1 ]
В глиогенезе
[ редактировать ]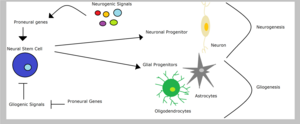
Нейронные стволовые клетки могут вызывать нейрональные или глиальные предшественники, в зависимости от типа сигналов, которые они получают - глиогенные или нейрогенные сигналы, соответственно. Глиальные клетки -предшественники могут дифференцироваться в олигодендроциты или астроциты . Тем не менее, приверженность линии нервных предшественников включает подавление альтернативных существ. Следовательно, гены пронераврала позвоночных способствуют судьбам нейронов и одновременно ингибируют глиальные судьбы. Например, подавление экспрессии пронураврального гена NGN2 в спинном мозге подавляет олигодендроцитов дифференцировку . In the context of restricted glial progenitors, proneural genes might have functions that are distinct from their better-characterized role in lineage specification, perhaps in the differentiation of glial lineages . [ 1 ] Sun and colleagues showed that proneural ngn1 inhibits gliogenesis by binding transcriptional co-activators like CBP/Smad1 or p300/Smad1 preventing the transcription of glialdifferentiation genes.[ 15 ] С другой стороны, сигнальный путь Notch способен способствовать глиогенезу в стволовых клетках посредством ингибирования проневральных генов, таких как MASH1 и нейрогенины . [ 5 ]
В регламенте клеточного цикла судьба
[ редактировать ]У позвоночных , хотя проневральные гены определяют нейронную судьбу предшественников, они также способствуют остановке своей стадии деления путем выделения уже указанных клеток-предшественников от влияния внешних сигналов, определяющих судьбу. Проневральные гены регулируют клеточный цикл путем активации циклин-зависимой киназы ('' CDK ') в некоторых линиях на уровне генов дифференцировки нейронов. С другой стороны, в таких беспозвоночных , как Drosophila , проневральные гены экспрессируются в основном в не дидифицированных клетках, но также могут быть экспрессируются в разделенных клетках, где комплекса Achaete-Scute было показано, что гены проневрала ингибируют прогрессирование клеточного цикла . [ 1 ]
В разработке сенсорных органов
[ редактировать ]Предуравральные гены также играют важную роль в разработке различных типов сенсорных органов , а именно аккордотональных органов ( ProprioeptorsThat определяют механические и звуковые вибрации) и внешние сенсорные органы . Члены комплекса Achaete-Scute , такие как Achaete и Scute , а также «атональные» и «без дочери», придают эктодермальные клетки способность становиться сенсорными материнскими клетками (SMC). [ 6 ] [ 10 ] При разработке сенсорных органов существует две основные фазы: определение и дифференциация, которые не могут быть механически разделены. Проневральные белки участвуют в обоих процессах посредством активации нисходящих «дифференцирующих генов» [ 25 ] Это, в свою очередь, регулирует индукцию характеристик сенсорно-органа-сабтипа. The specification of sensory organs by proneural genes is a complex process, since they elicit different cellular contexts. Например, у Drosophila атональный (ATO) может способствовать развитию хордотональных органов для рецепторов органов обонятельных чувств , в зависимости от изображного диска , в котором он экспрессируется. [ 1 ] В Drosophila эмбриогенезе ген- ахтет проневрал экспрессируется в хорошо определенных областях, как в эндодерме , отвечая за образование конкретных сенсорных органов у взрослых и личинок. Согласно Ruíz-Gomes and Ghysen (1993), эта экспрессия возникает в двух отдельных фазах: компетентном состоянии, в котором проннуральный ген экспрессируется в клеточном кластере; Определенное состояние, в котором специфическая клетка накапливает высокие уровни транскриптов «переменного тока», инициируя нейронного предшественника. [ 26 ] Функция каждого из комплексных генов ASC варьируется в зависимости от состояния развития (личинок или взрослых). [ 26 ] For example, in the adult state the ac and sc genes promote the differentiation of two sets of complementary sensory organs and ase gene as a minimal function, while in the larvae state ac and sc genes affect the same set of sensory organs and ase is responsible Для определения дополнительного набора. [ 27 ]
В кортикогенезе
[ редактировать ]В неокортексе существует широкая нейрональная сеть, поддерживаемая астроцитами и олигодендроцитами ( глиальные клетки ) с различными функциями. Во время развития коры факторы BHLH контролируют пролиферацию и дифференцировку нейронных клеток и их функции в любое время и место и место, зависит от их клеточного контекста. NeuroD , Ngns , Mash , ''Olig'' and other proneural gene families have a crucial role in cell fate decision during corticogenesis and different combinations of them regulate the choice and the timing of differentiation into a neuron , an astrocyte or an oligodendrocyte . [ 28 ] Высокие уровни NGN1 и NGN2 необходимы для определения нейрональной идентичности кортикальных предшественников только на ранних стадиях развития неокортекса . [ 29 ] В частности, NGN2 также важен для регулирования перехода кортикальных предшественников из желудочковой зоны в субвентрикулярную зону . С другой стороны, Mash1 участвует в ранней дифференциации стриатума нейронов [ 30 ] и достаточное для продвижения базальных клеточных делений независимо от его роли в спецификации существе нейрональных клеток на более поздних стадиях. Сотрудничество между NGN2 и пронуррельными генами MASH1 регулирует переход кортикальных предшественников от апикальных к базально -клеточным компартментам. [ 28 ] Спецификация различных нейрональных подтипов зависит от группы вовлеченных генов -проневралов. [ 31 ] Низкие уровни проеневральных транскриптов в желудочковой зоне экспрессируются, когда возникает спецификация предшественника, и увеличение их экспрессии приводит к началу нейрогенеза . NGN ответственны за образование глутаматергических нейронов , тогда как MASH1 порождает ГАМКергические и холинергические нейроны . [ 31 ]
При Drosophila развитии фоторецепторов
[ редактировать ]Атональный белок -белок (ATO) отвечает за развитие Drosophila R8 фоторецепторов . Тем не менее, он не действует в одиночку, поскольку он димеризируется со вторым идентичным белком, «без дочери» (DA) », который усиливает его экспрессию. Коэкспрессия «ато» и '' da '' 'важна для миграции различных типов клеток оматидия и для репрессии атональных в межкластерных пространствах, функционируя как ингибирующие сигналы, которые регулируют как число, так и число. положение зарождающихся клеток R8. Более того, эта экспрессия приводит к активации киназы MAPK , которая важна для рекрутирования клеток и эффекта репрессора, один раз, когда эта киназа неактивна, экспрессия «ато» обнаруживается в каждой клетке. [ 32 ] [ 33 ] В начальном состоянии Hedgehog ('' HH '' ') и DecapentAplegic ответственны за активацию' 'ato' 'в мелких клеточных кластерах, что приводит к активации MAPK . Во -первых, Hedgehog индуцирует экспрессию «ато», а не просто в клетках, которые в конечном итоге становятся фоторецепторными клетками , поэтому его экспрессия должна быть утончена и ограничена одной предполагаемой клеткой R8. Передача сигналов Hedgehog также необходима для подавления «атональной» экспрессии «атональная» выражение между зарождающимися предтеравральными кластерами, которая выявляет двойную роль, имеющую решающую роль в построении точности и геометрии в сетчатку взрослого . Следовательно, регуляция экспрессии «ато» зависит от уровней ежа : на низких уровнях (это далеко от источника) этот пронуклеарный ген активируется и на высоких уровнях (близко к источнику) он подавляется. Сочетая боковое ингибирование с двойной ролью, которую играет Hedgehog , можно представить, как может быть установлен шестиугольный массив клеток R8. [ 33 ]
НЕВЕРНАЯ ФУНКЦИИ
[ редактировать ]Помимо их роли в развитии нервной системы , пронуравральные гены также участвуют в процессах, связанных с инвазией трофобластов , дифференцировкой эндокринных клеток (а именно в поджелудочной железе и аденогипофизе ), определении пола и легких , щитовидной железой , надпочечниками и слюнных железами и развитию гастроиттоиджортальной системы. [ 34 ]
В инвазии трофобласта человека
[ редактировать ]В дополнение к их участии в нейрональной и глиальной дифференцировке, определении пола и развитии сенсорных органов , гены пронейрара также участвуют в дифференцировке трофобластов во время прогрессирования инвазии в плацентарной формировании. Исследования выявили экспрессию транскриптов NeuroD1 , NeuroD2 и «ATH2» в разных подмножествах инвазивного трофобласта . [ 35 ]
В развитии поджелудочной железы
[ редактировать ]В развивающейся поджелудочной железе фактор транскрипции NGN3 'отмечает популяции клеток, которые находятся в пути от недифференцированных эпителиальных клеток -предшественников в зрелые эндокринные клетки (предшественники эндокринных клеток поджелудочной железы), и, таким образом, не экспрессируют гормоны , которые предполагают, что этот ген является Выключен в дифференцированном гормонеположительном. [ 36 ] NGN3 '' является одновременно необходимым и достаточным для управления формированием островковых клеток во время развития поджелудочной железы, таким же, как спецификация нервной судьбы в нейроэктодерме . [ 37 ]
В определении пола
[ редактировать ]Хотя нейрогенез и определение пола , по -видимому, являются различными биологическими процессами, существуют доказательства того, что «без дочери (да») - важный ген для формирования всей Drosophila периферической нервной системы - также необходим для правильного определения пола . [ 38 ] В мухах Scute работает для прямого развития нейронов, но этот ген также приобрел роль в первичном событии определения пола - подсчет Х -хромосом - став элементом сигнала X -хромосом. [ 39 ]
В миогенезе
[ редактировать ]работают пронураранные гены Несмотря на то, что в эктодерме , леталь из Scute действует в соматической мезодерме для определения клеточного кластера, из которого предшественники мышц будут одиноки. [ 40 ] Взаимодействие между этими клетками и эктодермой приводит к образованию клеток -основателей мышц в аналогичном процессе с тем, который происходит в центральной нервной системе . [ 41 ]
В клеточной миграции
[ редактировать ]NGN1 и NGN2 регулировать механизмы миграции клеток и участвуют в первоначальном подавлении RHOA могут независимо прямо до того, как клетки нервных предшественников становятся постмитотическими, тогда как нейрод в первую очередь участвует в непрерывном подавлении RHOA во время миграции кортикальной миграции. Другие факторы BHLH , такие как «Math2», NeuroD2 и «NSCL1», могут подавлять экспрессию RHOA и регулировать механизм миграции в постмитотических нейронах . [ 42 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не Бертран, н; Кастро, DS; Guillemot, F (2002). «Предуравральные гены и спецификация типов нервных клеток». Nature Reports Neuroscience . 3 (7): 517–30. doi : 10.1038/nrn874 . PMID 12094208 . S2CID 205512016 .
- ^ Jump up to: а беременный Куниш, м; Haenlin, M; Кампос-Аорга, JA (1994). «Боковое ингибирование, опосредованное дельтой нейрогенного гена Drosophila, усиливается пронуравральными белками» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (21): 10139–43. Bibcode : 1994pnas ... 9110139K . doi : 10.1073/pnas.91.21.10139 . PMC 44973 . PMID 7937851 .
- ^ Tepass, u; Hartenstein, V (1995). «Нейрогенные и пронуравральные гены контролируют спецификацию судьбы клеток в эндодерме Drosophila». Разработка . 121 (2): 393–405. doi : 10.1242/dev.121.2.393 . PMID 7768181 .
- ^ Jump up to: а беременный в Брюне, JF; Ghysen, A (1999). «Деконструирование определения клеток: проневральные гены и идентичность нейронов» . Биологии . 21 (4): 313–8. doi : 10.1002/(SICI) 1521-1878 (199904) 21: 4 <313 :: AID-bies7> 3.0.co; 2-c . PMID 10377893 . S2CID 9577100 .
- ^ Jump up to: а беременный в Моррисон, SJ (2001). «Дифференцировка нейронов: пронивральные гены ингибируют глиогенез» . Текущая биология . 11 (9): R349–51. Bibcode : 2001cbio ... 11.r349m . doi : 10.1016/s0960-9822 (01) 00191-9 . PMID 11369245 .
- ^ Jump up to: а беременный Chien, Ct; Сяо, CD; Ян, я; Ян, YN (1996). «Информация о нейрональных типах, кодируемая в домене базовых списков-списко-спиральных генов» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (23): 13239–44. Bibcode : 1996pnas ... 9313239c . doi : 10.1073/pnas.93.23.13239 . PMC 24077 . PMID 8917575 .
- ^ Ледент, V; Paquet, o; Vervoort, M (2002). «Филогенетический анализ базовых белков-спираль-спираль человека» . Биология генома . 3 (6): Research0030. doi : 10.1186/gb-2002-3-6-research0030 . PMC 116727 . PMID 12093377 .
- ^ Гисен, а; Дермбер-хаудиер, C (1988). «От ДНК к форме: комплекс ахтета-склеата» . Гены и развитие . 2 (5): 495–501. doi : 10.1101/gad.2.5.495 . PMID 3290049 .
- ^ Jump up to: а беременный Гонсалес, F; Романи, с; Кубас, P; Модолелл, J; Campuzano, S (1989). «Молекулярный анализ гена Asense, члена комплекса Achaete-Scute Drosophila melanogaster и его новую роль в развитии зрительного доли» . Embo Journal . 8 (12): 3553–62. doi : 10.1002/j.1460-2075.1989.tb08527.x . PMC 402034 . PMID 2510998 .
- ^ Jump up to: а беременный Jarman, AP; Grau, y; Ян, я; Ян, YN (1993). «Атональный - это ген -поневраль, который направляет образование аккордотональных органов в периферической нервной системе дрозофилы» . Клетка . 73 (7): 1307–21. doi : 10.1016/0092-8674 (93) 90358-w . PMID 8324823 .
- ^ Гулдинг, SE; Белый, NM; Jarman, AP (2000). «Cato кодирует базовый фактор транскрипции спиральной спирали, участвующий в правильной дифференцировке органов чувства дрозофилы» . Биология развития . 221 (1): 120–31. doi : 10.1006/dbio.2000.9677 . PMID 10772796 .
- ^ Ян, ин; Ян, Ли (1993). «Белки HLH, нейрогенез мух и миогенез позвоночных» . Клетка . 75 (5): 827–30. doi : 10.1016/0092-8674 (93) 90525-U . PMID 8252617 .
- ^ Кампос-Аорга, JA (1993). «Механизмы раннего нейрогенеза у Drosophila melanogaster». Журнал нейробиологии . 24 (10): 1305–27. doi : 10.1002/neu.480241005 . PMID 8228961 .
- ^ Пауэлл, LM; Jarman, AP (2008). «Контекстная зависимость проннуральных белков BHLH» . Текущее мнение в области генетики и развития . 18 (5): 411–7. doi : 10.1016/j.gde.2008.07.012 . PMC 3287282 . PMID 18722526 .
- ^ Jump up to: а беременный Солнце, у; Nadal-Vicens, M; Мисоно, с; Лин, MZ; Зубиага, а; Хуа, х; Фанат, g; Гринберг, ME (2001). «Нейроген способствует нейрогенезу и ингибирует глиальную дифференцировку с помощью независимых механизмов» . Клетка . 104 (3): 365–76. doi : 10.1016/s0092-8674 (01) 00224-0 . PMID 11239394 .
- ^ Jump up to: а беременный в дюймовый Chitnis, a; Кинтнер, С. (1996). «Чувствительность пронуравральных генов к боковому ингибированию влияет на паттерн первичных нейронов у эмбрионов Xenopus». Разработка . 122 (7): 2295–301. doi : 10.1242/dev.122.7.2295 . PMID 8681809 .
- ^ Schlatter, R.; Maier, D. (2005). «Энхансер разделенных и ахтевых комплексов Drosophilids, полученных из простых UR-комплексов, сохранившихся в комарах и пчелах» . BMC Эволюционная биология . 5 (1): 67. Bibcode : 2005bmcee ... 5 ... 67 с . doi : 10.1186/1471-2148-5-67 . PMC 1310631 . PMID 16293187 .
- ^ Кубо, F; Takeichi, M; Накагава, S (2005). «Wnt2b ингибирует дифференцировку клеток -предшественников сетчатки в отсутствие активности Notch, подавляя экспрессию пронуравральных генов» . Разработка . 132 (12): 2759–70. doi : 10.1242/dev.01856 . PMID 15901663 .
- ^ Skeath, JB; Панганибан, GF; Кэрролл, С.Б. (1994). «Дефектный ген вентральной нервной системы контролирует экспрессию проннурального гена на двух различных этапах во время образования нейробластов у дрозофилы». Разработка . 120 (6): 1517–24. doi : 10.1242/dev.120.6.1517 . PMID 8050360 .
- ^ Jump up to: а беременный Skeath, JB; Кэрролл, С.Б. (1992). «Регуляция экспрессии гена пронураврала и клеточной судьбы во время сегрегации нейробластов у эмбриона дрозофилы». Разработка . 114 (4): 939–46. doi : 10.1242/dev.114.4.939 . PMID 1618155 .
- ^ Урбах, R; Schnabel, R; Technau, GM (2003). «Паттерн образования нейробластов, митотических доменов и экспрессии генов проннуральны во время раннего развития мозга у дрозофилы» . Разработка . 130 (16): 3589–606. doi : 10.1242/dev.00528 . PMID 12835378 .
- ^ Hartenstein, V; Tepass, u; Грузинский-дефео, Е. (1996). «Проневральные и нейрогенные гены контролируют спецификацию и морфогенез предшественников стоматогастральных нервных клеток у дрозофилы» . Биология развития . 173 (1): 213–27. doi : 10.1006/dbio.1996.0018 . PMID 8575623 .
- ^ Jump up to: а беременный Ratié, L; Посуда, м; Barloy-Hubler, F; Роме, ч; Гиккель, я; Дубург, C; Дэвид, V; Dupé, V (2013). «Новые гены активировались, когда передача сигналов Notch нарушается во время развития гипоталамуса» . Нейронное развитие . 8 : 25. doi : 10.1186/1749-8104-8-25 . PMC 3880542 . PMID 24360028 .
- ^ Kageyama, R.; Ohtsuka, T.; Shimojo, H.; Имайоши И. (2008). «Динамическая передача сигналов Notch в клетках нервных предшественников и пересмотренный взгляд на боковое ингибирование». Nature Neuroscience . 11 (11): 1247–1251. doi : 10.1038/nn.2208 . PMID 18956012 . S2CID 24613095 .
- ^ Кубас, P; Де Селис, JF; Campuzano, S; Модолелл, Дж. (1991). «Предураранные кластеры экспрессии ахтета и генерации сенсорных органов в диском видации Drosophila Imaginal Wing» . Гены и развитие . 5 (6): 996–1008. doi : 10.1101/gad.5.6.996 . PMID 2044965 .
- ^ Jump up to: а беременный Руис-Гомес, м; Ghysen, A (1993). «Экспрессия и роль проневрального гена Achaete, в развитии личиночной нервной системы дрозофилы» . Embo Journal . 12 (3): 1121–30. doi : 10.1002/j.1460-2075.1993.tb05753.x . PMC 413313 . PMID 8458326 .
- ^ Бэмбли-Чаудьера, C.; Гисен А. (1987). «Независимые подчинения органов чувств требуют независимых генов комплекса ахтета-склера у личинок дрозофилы» . Гены и развитие . 1 (3): 297–306. doi : 10.1101/gad.1.3.297 .
- ^ Jump up to: а беременный Росс, SE; Гринберг, я; Стайлз, CD (2003). «Основные факторы спирали-петли-спирали в развитии кортикальной коры» . Нейрон . 39 (1): 13–25. doi : 10.1016/s0896-6273 (03) 00365-9 . PMID 12848929 .
- ^ Schuurmans, C; Армант, о; Nieto, M; Стенман, JM; Бритц, о; Кленин, н; Браун, C; Langevin, LM; Seibt, J; Тан, ч; Каннингем, JM; Dyck, R; Уолш, C; Кэмпбелл, K; Polleux, F; Guillemot, F (2004). «Последовательные фазы спецификации кортикальной коры включают нейрогенин -зависимые и -индизависимые пути» . Embo Journal . 23 (14): 2892–902. doi : 10.1038/sj.emboj.7600278 . PMC 514942 . PMID 15229646 .
- ^ Юн, К; Fischman, S; Джонсон, J; Hrabe de Angelis, M; Weinmaster, G; Рубенштейн, JL (2002). «Модуляция передачи сигналов Notch с помощью MASH1 и DLX1/2 регулирует последовательную спецификацию и дифференцировку типов клеток -предшественников в подкорковой телэнсфалоне». Разработка . 129 (21): 5029–40. doi : 10.1242/dev.129.21.5029 . PMID 12397111 .
- ^ Jump up to: а беременный Бритц, о; Маттар, P; Нгуен, L; Langevin, LM; Циммер, C; Alam, S; Guillemot, F; Schuurmans, C (2006). «Роль для проннуральных генов в созревании клеток -предшественников кортикальных препаратов» . Кора головного мозга . 16 (Suppl 1): I138–51. doi : 10.1093/cercor/bhj168 . PMID 16766700 .
- ^ Чен, CK; Chien, CT (1999). «Негативная регуляция атональной в проннойрной кластере образование фоторецепторов Drosophila R8» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (9): 5055–60. Bibcode : 1999pnas ... 96.5055c . doi : 10.1073/pnas.96.9.5055 . PMC 21815 . PMID 10220417 .
- ^ Jump up to: а беременный Domínguez, M (1999). «Двойная роль ежа в регуляции атонального гена проневрала во время развития Омматидии» . Разработка . 126 (11): 2345–53. doi : 10.1242/dev.126.11.2345 . HDL : 10261/307536 . PMID 10225994 .
- ^ Pogoda, HM; von der Hardt, s; Герцог, w; Крамер, C; Schwarz, h; Hammerschmidt, M (2006). «Предуравральный ген ASCL1A необходим для эндокринной дифференцировки и выживания клеток в аденогипофизе рыбок данио» . Разработка . 133 (6): 1079–89. doi : 10.1242/dev.02296 . PMID 16481349 .
- ^ Западный человек, бакалавр; Путсма, а; Maruyama, K; Schrians, HF; От слабых, ij; Oudjans, CB (2002). «Гены пронантеральных Neurod1 и NeuroD2 - это явно Thumblast Thumblast Trophoblast Invasion» . Механиды или разработка . 113 (1): 85–90. doi : 10,1016/S0925-473 (01) 0065-7 . PMID 11900979 .
- ^ Йоханссон, Ка; Dursun, U.; Джордан, Н.; Гу, Г.; Beermann, F.; Gradwohl, Gr; Грапин-Боттон, А. (2007). «Временный контроль активности нейрогенана в предшественниках поджелудочной железы выявляет окна компетенции для генерации различных типов эндокринных клеток» . Ячейка развития . 12 (3): 457–465. doi : 10.1016/j.devcel.2007.02.010 . PMID 17336910 .
- ^ Ли, JC; Смит, SB; Ватада, ч; Лин, J; Scheel, D; Ван, J; Mirmira, RG; Немец, MS (2001). «Регуляция про-эндокринного гена поджелудочной железы» . Диабет . 50 (5): 928–36. doi : 10.2337/диабет.50.5.928 . PMID 11334435 .
- ^ Cline, TW (1976). «Пол-специфический, чувствительный к температуре материнский эффект не дочери мутации Drosophila melanogaster» . Генетика . 84 (4): 723–42. doi : 10.1093/Genetics/84.4.723 . PMC 1213604 . PMID 827461 .
- ^ Wrischnik, La; Тиммер, младший; Межа, Ла; Cline, TW (2003). «Рекрутирование проневрального гена Scute к пути определения пола Drosophila» . Генетика . 165 (4): 2007–27. doi : 10.1093/Genetics/165.4.2007 . PMC 1462923 . PMID 14704182 .
- ^ Кармен, а; Бейт, м; Jiménez, F (1995). «Смертельный из Scute, пронуравральный ген, участвует в спецификации мышечных предшественников во время эмбриогенеза Drosophila» . Гены и развитие . 9 (19): 2373–83. doi : 10.1101/gad.9.19.2373 . PMID 7557389 .
- ^ Бейт, CM; Grunewald, EB (1981). «Эмбриогенез нервной системы насекомых II: второй класс клеток -предшественников нейронов и происхождение межсегментарных соединений». Журнал эмбриологии и экспериментальной морфологии . 61 : 317–30. PMID 7264548 .
- ^ Ge, w; Он, f; Ким, KJ; Бланчи, б; Coskun, V; Нгуен, L; Wu, x; Чжао, J; Хенг, Джи; Martinowich, K; Дао, J; Wu, h; Кастро, D; Sobeih, MM; Corfas, g; Глисон, JG; Гринберг, я; Guillemot, F; Sun, Ye (2006). «Соединение клеточной миграции с нейрогенезом с помощью пронейральных факторов BHLH» . Труды Национальной академии наук . 103 (5): 1319–24. Bibcode : 2006pnas..103.1319G . doi : 10.1073/pnas.0510419103 . PMC 1345712 . PMID 16432194 .