CUL4A
| CUL4A | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | КУЛ4А , куллин 4А | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 603137 ; МГИ : 1914487 ; Гомологен : 81724 ; Генные карты : CUL4A ; OMA : CUL4A — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||


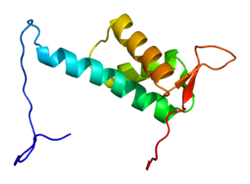
Куллин-4А — это белок , который у человека кодируется CUL4A геном . [ 4 ] [ 5 ] CUL4A принадлежит к и семейству белков убиквитинлигазы куллинов очень гомологичен белку CUL4B . CUL4A регулирует многочисленные ключевые процессы, такие как восстановление ДНК, ремоделирование хроматина , сперматогенез , кроветворение и митотический клеточный цикл . В результате CUL4A участвует в развитии нескольких видов рака и патогенезе некоторых вирусов, включая ВИЧ . Было обнаружено, что компонент комплекса CUL4A, Цереблон, является основной мишенью тератогенного агента талидомида .
Структура
[ редактировать ]Белок CUL4A имеет длину 759 аминокислот и образует протяженную жесткую структуру, состоящую в основном из альфа-спиралей . На N-конце CUL4A связывается с бета-пропеллером адаптерного белка DDB1 , который взаимодействует с многочисленными DDB1-CUL4-ассоциированными факторами (DCAF). В результате N-конец имеет решающее значение для рекрутирования субстратов для комплекса убиквитинлигазы . На С- конце CUL4A взаимодействует с белком RBX1 /ROC1 через его домен RING . RBX1 является основным компонентом комплексов убиквитинлигазы (CRL) Cullin-RING и выполняет функцию рекрутирования ферментов, конъюгирующих убиквитин E2 . Следовательно, С-конец CUL4A вместе с RBX1 и активированными ферментами E2 составляет каталитическое ядро комплексов CRL4. CUL4A также модифицируется путем ковалентного присоединения молекулы NEDD8 к высококонсервативному остатку лизина в С-концевой области. Эта модификация, по-видимому, вызывает конформационные изменения, которые способствуют гибкости домена RING белков куллина и повышению активности убиквитинлигазы. [ 6 ]
В целом комплексы CRL4A имеют модульную структуру, которая позволяет осуществлять сложную регуляцию клеткой и влиять на многочисленные субстраты и процессы в клетке. Хотя отдельные части различаются, все убиквитинлигазы на основе куллина обладают этими характеристиками. [ 7 ]
Функция
[ редактировать ]Повреждение и восстановление ДНК
[ редактировать ]Адаптерный белок DDB1 первоначально был охарактеризован как большая субъединица гетеродимерного комплекса (UV-DDB), который, как было обнаружено, распознает поврежденную ДНК и участвует в форме восстановления, известной как эксцизионная репарация нуклеотидов (NER). Меньшая субъединица этого белкового комплекса, связывающего поврежденную ДНК, известна как DDB2 и способна напрямую связывать повреждения ДНК, связанные с УФ-облучением. DDB2 представляет собой белок DCAF и одновременно является субстратом убиквитинирования комплекса CRL4, а также служит белком лигазы E3 для других субстратов, таких как XPC и гистоны (см. следующий раздел) вблизи места повреждения. [ 8 ] Из-за убиквитинирования белков DDB2 и XPC, распознающих повреждения ДНК, CUL4A был описан как негативный регулятор активности NER. [ 9 ] [ 10 ] В дополнение к «глобальному» типу NER, комплекс CRL4A, по-видимому, также играет роль в «связанном с транскрипцией» NER в сочетании с белком синдрома А Коккейна . [ 11 ] Комплексы CRL4A, по-видимому, активируются определенными типами повреждений ДНК (особенно УФ-облучением), а некоторые субстраты преимущественно убиквитинируются после индукции повреждения ДНК.
Ремоделирование хроматина
[ редактировать ]Роль CUL4A в модификации хроматина во многом связана с деятельностью по восстановлению ДНК и происходит после индукции повреждения ДНК. И CUL4A, и его близкородственный гомолог CUL4B могут убиквитинировать гистоны H2A, H3 и H4. [ 12 ] [ 13 ] Дрожжевой гомолог CUL4A, Rtt101, убиквитинирует гистон H3 и способствует сборке нуклеосом , а комплексы CRL4A выполняют аналогичные функции в клетках человека. [ 14 ] Комплексы CRL4 также влияют на события метилирования гистонов и структуру хроматина посредством регуляции гистоновых метилтрансфераз . [ 15 ] Монометилаза гистона H4 PR-Set7/SET8 убиквитинируется на хроматине комплексами CRL4(Cdt2) во время S-фазы и после повреждения ДНК PCNA -зависимым образом. [ 16 ] [ 17 ] [ 18 ]
Регуляция клеточного цикла и репликации ДНК
[ редактировать ]Комплексы CRL4A регулируют вход в фазу синтеза ДНК, или S-фазу , митотического цикла путем регулирования уровней экспрессии белка фактора лицензирования репликации Cdt1 и ингибитора циклин-зависимой киназы p21 . В обоих случаях CRL4A использует Cdt2 в качестве DCAF для связывания обоих субстратов PCNA-зависимым образом. Во время ненарушенного клеточного цикла происходит убиквитинирование и подавление этих белков с помощью CRL4A. CDT2 происходит в начале репликации ДНК. Повреждение ДНК, такое как УФ-облучение, также индуцирует CRL4A. CDT2 -опосредованное разрушение этих белков. Оба субстрата также регулируются SCF. Скп2 сложный .
CRL4-опосредованное разрушение p21 снимает ингибирование циклина E - Cdk2 и способствует входу в S-фазу. Потеря экспрессии Cdt2 увеличивает экспрессию p21 в клетках и стабилизирует p21 после УФ-облучения. [ 19 ] Делеция CUL4A приводит к задержке входа в S-фазу в эмбриональных фибробластах мыши, что устраняется делецией p21. [ 10 ] В клетках пигментного эпителия сетчатки человека потеря экспрессии Cdt2 также приводит к p21-зависимому задержке входа в S-фазу и повторной экспрессии p21 в S-фазе, что приводит к циклам неполной репликации, долговременному накоплению p21 и, в некоторых случаях, к повторной экспрессии p21 в S-фазе. индукция апоптоза. [ 20 ]
После стимулирования инициации репликации эукариотической ДНК в источнике Cdt1 инактивируется геминином и направляется на деградацию с помощью SCF. Скп2 и CRL4 CDT2 комплексы. Экспрессия Cdt1 стабилизируется за счет РНКи-опосредованного нокдауна DDB1 или обоих CUL4A и CUL4B, что предполагает избыточную или перекрывающуюся функцию двух белков CUL4 для регуляции Cdt1. [ 21 ] [ 22 ] Только снижение экспрессии Geminin, по-видимому, индуцирует повторную репликацию в клетках, сверхэкспрессирующих Cdt1.
CRL4 также используют Cdt2 и PCNA для разрушения субъединицы p12 ДНК-полимеразы δ во время S-фазы и после УФ-облучения. [ 23 ]
Кроветворение
[ редактировать ]Комплексы CRL4A, по-видимому, индуцируют деградацию многочисленных членов транскрипционного семейства НОХ , которые являются важными регуляторами кроветворения. [ 24 ] Первым членом семейства HOX, идентифицированным как мишень CRL4A-опосредованной деградации, является HOXA9 , который необходим для поддержания гемопоэтических стволовых клеток и участвует в подмножестве миелолейкозов . [ 25 ] [ 26 ] HOXA9 Дегрон находится внутри гомеодомена , который имеет решающее значение для связывания ДНК. Исследования выравнивания последовательностей показали, что внутри одной спирали гомеодомена имеется высококонсервативный мотив «LEXE». Когда несколько аминокислот в этом мотиве были мутированы, HOXB4 стал устойчивым к деградации, опосредованной CRL4A. [ 24 ] Субстратный рецептор, или DCAF, необходимый для деградации белка HOX, остается неизвестным.
Сперматогенез и мейоз
[ редактировать ]Ген Cul4a необходим для нормального сперматогенеза и мейоза в мужских половых клетках мышей. [ 27 ] [ 28 ] Cul4a −/− мужчины производят аномальную сперму и бесплодны. Хотя CUL4A и CUL4B экспрессируются в мужских гаметах, CUL4A в высокой степени экспрессируется в пахитенах и диплотенах . Именно на этих стадиях мужские половые клетки с дефицитом CUL4A демонстрируют высокий уровень апоптоза , неправильную репарацию ДНК и накопление субстрата CRL4 Cdt1 .
Дисрегуляция
[ редактировать ]Рак
[ редактировать ]Хромосомная область ch13q34, содержащая ген CUL4A , амплифицируется в 3–6% некоторых карцином , включая рак молочной железы, матки, легких, желудка и колоректальный рак. [ 29 ] CUL4A также мутирует или амплифицируется примерно в 4% меланом (хотя мутации рассредоточены, а отдельные мутации возникают спорадически).
В моделях на мышах нокаут Cul4a приводил к выраженной устойчивости к канцерогенезу кожи, индуцированному УФ-излучением. [ 10 ] Cre -индуцированная сверхэкспрессия Cul4a в тканях легких мышей способствовала гиперплазии . [ 30 ]
В связи с наблюдаемой амплификацией CUL4A в нескольких карциномах и тем фактом, что комплексы CRL4 нацелены на множественные гены репарации ДНК и гены-супрессоры опухолей , CUL4A можно рассматривать как онкоген в определенных контекстах .
Вирусный патогенез
[ редактировать ]Благодаря своей устойчивой экспрессии (особенно во время репликации ДНК) и модульной природе комплексы CRL4A могут быть использованы или «захвачены» для стимулирования пролиферации вируса в клетках млекопитающих.
Некоторые парамиксовирусы избегают интерфероновой реакции в клетках, нацеливаясь на STAT1 и нарушая передачу сигналов. Вирус обезьян 5 и вирус парагриппа человека типа II экспрессируют белок, названный «V», который действует как рецептор-субстрат и обеспечивает взаимодействие между белками DDB1 и STAT (структура CRL4A СВ5В комплекс изображен на вставке) - тем самым индуцируя убиквитинирование и деградацию STAT1. [ 31 ] [ 32 ]
DCAF1 также называют VPRBP ВИЧ-1 из-за его взаимодействия с белком Vpr . Хотя DCAF1/VPRBP, по-видимому, выполняет решающую функцию в подавлении опухоли, репликации ДНК и эмбриональном развитии, ВИЧ-1 «похищает» комплекс убиквитинлигазы, вызывая остановку клеточного цикла в фазе G2 . [ 33 ] [ 34 ] [ 35 ] CRL4A DCAF1-Впр индуцирует убиквитинирование ядерной изоформы урацил-ДНК-гликозилазы . [ 36 ] [ 37 ] ВИЧ-2 также, по-видимому, использует CRL4A. ДКВА1 посредством Vpx индуцированного белком разрушения лентивирус -ингибирующей дезоксинуклеозидтрифосфогидролазы под названием SAMHD1 . [ 38 ] [ 39 ]
Лечение талидомидом
[ редактировать ]В 2010 году Ито и др. сообщили, что Цереблон, белок DCAF, был основной мишенью тератогенного соединения талидомида. [ 40 ] Талидомид и другие производные, такие как помалидомид и леналидомид, известны как иммуномодулирующие препараты (или IMiD) и исследовались в качестве терапевтических средств при аутоиммунных заболеваниях и некоторых видах рака, особенно миеломах. Недавние отчеты показывают, что IMiD связываются с CRL4. КРБН и способствуют деградации факторов транскрипции IKZF1 и IKZF3, на которые обычно не воздействуют комплексы CRL4. [ 41 ] [ 42 ]
Взаимодействия и субстраты
[ редактировать ]Человеческий CUL4A образует прямое взаимодействие с:
Человеческие комплексы CUL4A-DDB1-RBX1 способствуют убиквитинированию:
† белок является субстратом CRL4A только тогда, когда он направляется вирусными белками.
§ белок является субстратом CRL4A только под действием IMiD.
Примечания
[ редактировать ]
Версия этой статьи 2015 года была обновлена внешним экспертом в рамках модели двойной публикации. Соответствующая академическая рецензируемая статья была опубликована в журнале Gene и может цитироваться как: Джеффри Ханна; Пэнбо Чжоу (15 ноября 2015 г.). «Различные и перекрывающиеся функции каркасных белков куллиновой лигазы CUL4A и CUL4B» . Джин . Серия обзоров Gene Wiki. 573 (1): 33–45. дои : 10.1016/J.GENE.2015.08.064 . ISSN 0378-1119 . ПМК 5110433 . ПМИД 26344709 . Викиданные Q28086765 . |
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031446 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Кипреос ET, Лендер LE, Wing JP, He WW, Hedgecock EM (июнь 1996 г.). «cul-1 необходим для выхода из клеточного цикла у C. elegans и идентифицирует новое семейство генов» . Клетка . 85 (6): 829–39. дои : 10.1016/S0092-8674(00)81267-2 . ПМИД 8681378 . S2CID 15805562 .
- ^ «Ген Энтреза: CUL4A Каллин 4А» .
- ^ Дуда Д.М., Борг Л.А., Скотт Д.К., Хант Х.В., Хаммел М., Шульман Б.А. (сентябрь 2008 г.). «Структурные данные об активации NEDD8 лигаз куллин-RING: конформационный контроль конъюгации» . Клетка . 134 (6): 995–1006. дои : 10.1016/j.cell.2008.07.022 . ПМЦ 2628631 . ПМИД 18805092 .
- ^ Босу Д.Р., Кипреос Э.Т. (2008). «Убиквитинлигазы Cullin-RING: глобальные циклы регуляции и активации» . Отделение клеток . 3 :7. дои : 10.1186/1747-1028-3-7 . ПМК 2266742 . ПМИД 18282298 .
- ^ Перейти обратно: а б Сугасава К., Окуда Й., Сайдзё М., Ниси Р., Мацуда Н., Чу Г., Мори Т., Иваи С., Танака К., Танака К., Ханаока Ф (май 2005 г.). «Индуцированное УФ-излучением убиквитилирование белка XPC, опосредованное комплексом УФ-DDB-убиквитинлигазы» . Ячейка 121 (3): 387–400. дои : 10.1016/j.cell.2005.02.035 . ПМИД 15882621 . S2CID 16773579 .
- ^ Чен X, Чжан Дж, Ли Дж, Линь П.С., Форд Дж.М., Чжэн Н., Чжоу П. (май 2006 г.). «Независимая от киназы функция c-Abl в содействии протеолитическому разрушению поврежденных ДНК-связывающих белков» . Молекулярная клетка . 22 (4): 489–99. doi : 10.1016/j.molcel.2006.04.021 . ПМИД 16713579 .
- ^ Перейти обратно: а б с д Лю Л., Ли С., Чжан Дж., Питерс С.Б., Ханна Дж., Чжан Ю., Инь Ю., Кофф А., Ма Л., Чжоу П. (май 2009 г.). «Отмена CUL4A усиливает реакцию на повреждение ДНК и защиту от канцерогенеза кожи» . Молекулярная клетка . 34 (4): 451–60. doi : 10.1016/j.molcel.2009.04.020 . ПМЦ 2722740 . ПМИД 19481525 .
- ^ Ханна Дж., Чжоу П. (апрель 2009 г.). «Регуляция путей ответа на повреждение ДНК с помощью убиквитинлигазы куллин-RING» . Восстановление ДНК . 8 (4): 536–43. дои : 10.1016/j.dnarep.2009.01.011 . ПМЦ 2858918 . ПМИД 19231300 .
- ^ Герреро-Санторо Дж., Капетанаки М.Г., Се С.Л., Горбачинский И., Левин А.С., Рапич-Отрин В. (июль 2008 г.). «ДНК-связывающая протеинлигаза на основе куллина 4B, поврежденная УФ-излучением, связывается с поврежденным УФ-излучением хроматином и убиквитинирует гистон H2A» . Исследования рака . 68 (13): 5014–22. дои : 10.1158/0008-5472.CAN-07-6162 . ПМИД 18593899 .
- ^ Ван Х., Чжай Л., Сюй Дж., Джу ХИ, Джексон С., Эрджумент-Бромаж Х., Темпст П., Сюн Ю., Чжан Ю. (май 2006 г.). «Убиквитилирование гистонов H3 и H4 с помощью убиквитинлигазы CUL4-DDB-ROC1 облегчает клеточный ответ на повреждение ДНК» . Молекулярная клетка . 22 (3): 383–94. doi : 10.1016/j.molcel.2006.03.035 . ПМИД 16678110 . S2CID 25677159 .
- ^ Хан Дж, Чжан Х, Чжан Х, Ван Цз, Чжоу Х, Чжан Цз (ноябрь 2013 г.). «Убиквитинлигаза Cul4 E3 регулирует передачу гистонов во время сборки нуклеосомы» . Клетка . 155 (4): 817–29. дои : 10.1016/j.cell.2013.10.014 . ПМЦ 3994564 . ПМИД 24209620 .
- ^ Хига Л.А., Ву М., Йе Т., Кобаяши Р., Сунь Х., Чжан Х. (ноябрь 2006 г.). «Убиквитинлигаза CUL4-DDB1 взаимодействует с множеством белков-повторов WD40 и регулирует метилирование гистонов». Природная клеточная биология . 8 (11): 1277–83. дои : 10.1038/ncb1490 . hdl : 10397/34293 . ПМИД 17041588 . S2CID 22180568 .
- ^ Перейти обратно: а б Йоргенсен С., Эскильдсен М., Фуггер К., Хансен Л., Ларсен М.С., Кушольт А.Н., Сильюасен Р.Г., Трелле М.Б., Йенсен О.Н., Хелин К., Соренсен К.С. (январь 2011 г.). «SET8 разлагается посредством убиквитилирования CRL4 (CDT2), связанного с PCNA, в S-фазе и после УФ-облучения» . Журнал клеточной биологии . 192 (1): 43–54. дои : 10.1083/jcb.201009076 . ПМК 3019552 . ПМИД 21220508 .
- ^ Перейти обратно: а б Тардат М., Брюстель Дж., Кирш О., Лефевбр К., Калланан М., Сардет К., Жюльен Э. (ноябрь 2010 г.). «Гистон H4 Lys 20 метилтрансфераза PR-Set7 регулирует начало репликации в клетках млекопитающих». Природная клеточная биология . 12 (11): 1086–93. дои : 10.1038/ncb2113 . ПМИД 20953199 . S2CID 6710289 .
- ^ Перейти обратно: а б Ода Х, Хюбнер М.Р., Бек Д.Б., Вермюлен М., Гурвиц Дж., Спектор Д.Л., Рейнберг Д. (ноябрь 2010 г.). «Регуляция монометилазы гистона H4 PR-Set7 посредством CRL4(Cdt2)-опосредованной PCNA-зависимой деградации во время повреждения ДНК» . Молекулярная клетка . 40 (3): 364–76. дои : 10.1016/j.molcel.2010.10.011 . ПМЦ 2999913 . ПМИД 21035370 .
- ^ Перейти обратно: а б Аббас Т., Сивапрасад У., Тераи К., Ловер В., Пэган М., Дутта А. (сентябрь 2008 г.). «PCNA-зависимая регуляция убиквитилирования и деградации p21 через комплекс убиквитинлигазы CRL4Cdt2» . Гены и развитие . 22 (18): 2496–506. дои : 10.1101/gad.1676108 . ПМК 2546691 . ПМИД 18794347 .
- ^ Барр, Алексис Р.; Купер, Сэмюэл; Хелдт, Фрэнк С.; Бутера, Франческа; Стой, Генриетта; Мансфельд, Йорг; Новак, Бела; Бакал, Крис (2017). «Повреждение ДНК во время S-фазы опосредует решение о прекращении пролиферации в последующем G1 посредством экспрессии p21» . Природные коммуникации . 8 : 14728. Бибкод : 2017NatCo...814728B . дои : 10.1038/ncomms14728 . ПМЦ 5364389 . ПМИД 28317845 .
- ^ Перейти обратно: а б Хига Л.А., Михайлов И.С., Бэнкс Д.П., Чжэн Дж., Чжан Х. (ноябрь 2003 г.). «Радиационно-опосредованный протеолиз CDT1 комплексами CUL4-ROC1 и CSN представляет собой новую контрольную точку». Природная клеточная биология . 5 (11): 1008–15. дои : 10.1038/ncb1061 . ПМИД 14578910 . S2CID 26070877 .
- ^ Перейти обратно: а б Ху Дж, Сюн Ю (февраль 2006 г.). «Эволюционно консервативная функция пролиферирующего клеточного ядерного антигена для деградации Cdt1 убиквитинлигазой Cul4-Ddb1 в ответ на повреждение ДНК» . Журнал биологической химии . 281 (7): 3753–6. дои : 10.1074/jbc.C500464200 . ПМИД 16407242 .
- ^ Перейти обратно: а б Чжан С., Чжао Х., Даржинкевич З., Чжоу П., Чжан З., Ли Э.Ю. , Ли М.Ю. (2013). «Новая функция CRL4(Cdt2): регуляция структуры субъединиц ДНК-полимеразы δ в ответ на повреждение ДНК и во время S-фазы» . Ж. Биол. Хим . 288 (41): 29550–61. дои : 10.1074/jbc.M113.490466 . ПМЦ 3795253 . ПМИД 23913683 .
- ^ Перейти обратно: а б с Ли Дж., Ши Дж.Х., Чжан Дж., Лю Л., Чжан Ю., Эом Дж.Й., Моррон Дж., Мур М.А., Чжоу П. (май 2013 г.). «Улучшенное расширение ex vivo взрослых гемопоэтических стволовых клеток за счет преодоления CUL4-опосредованной деградации HOXB4» . Кровь . 121 (20): 4082–9. doi : 10.1182/blood-2012-09-455204 . ПМЦ 3656448 . ПМИД 23520338 .
- ^ Перейти обратно: а б Чжан Ю, Моррон Дж, Чжан Дж, Чен Х, Лу Х, Ма Л, Мур М, Чжоу П (ноябрь 2003 г.). «CUL-4A стимулирует убиквитилирование и деградацию гомеодоменного белка HOXA9» . Журнал ЭМБО . 22 (22): 6057–67. дои : 10.1093/emboj/cdg577 . ПМЦ 275435 . ПМИД 14609952 .
- ^ Лоуренс, HJ; Кристенсен, Дж.; Фонг, С.; Ху, ЮЛ; Вайсман, И.; Соважо, Г.; Хамфрис, РК; Ларгман, К. (2005). «Потеря экспрессии гомеобоксного гена Hoxa-9 ухудшает пролиферацию и репопуляционную способность гемопоэтических стволовых клеток» . Кровь . 106 (12): 3988–3994. дои : 10.1182/кровь-2005-05-2003 . ПМЦ 1895111 . ПМИД 16091451 .
- ^ Инь Ю, Линь С., Ким С.Т., Ройг И., Чен Х., Лю Л., Вейт Г.М., Джин РУ, Кини С., Джасин М., Моли К., Чжоу П., Ма Л. (2011). «Убиквитинлигаза Е3 Каллин 4А регулирует мейотическую прогрессию в сперматогенезе мышей» . Дев. Биол . 356 (1): 51–62. дои : 10.1016/j.ydbio.2011.05.661 . ПМК 3130830 . ПМИД 21624359 .
- ^ Копанджа Д., Рой Н., Стоянова Т., Хесс Р.А., Багчи С., Райчаудхури П. (2011). «Cul4A необходим для сперматогенеза и мужской фертильности» . Дев. Биол . 352 (2): 278–87. дои : 10.1016/j.ydbio.2011.01.028 . ПМК 3065526 . ПМИД 21291880 .
- ^ «cBioPortal для геномики рака» . Архивировано из оригинала 21 мая 2015 г.
- ^ Ли Т., Хунг М.С., Ван Й., Мао Дж.Х., Тан Дж.Л., Джахан К., Роос Х., Сюй З., Яблонс Д.М., Ю Л. (март 2011 г.). «Трансгенные мыши для Cre-индуцируемой сверхэкспрессии гена Cul4A» . Бытие . 49 (3): 134–41. дои : 10.1002/dvg.20708 . ПМЦ 3285554 . ПМИД 21381181 .
- ^ Перейти обратно: а б Улане CM, Кенцис А, Круз CD, Парижьен JP, Шнайдер KL, Хорват CM (август 2005 г.). «Состав и сборка комплексов убиквитинлигазы, нацеленных на STAT: карбоксильный конец белка V парамиксовируса представляет собой домен олигомеризации» . Журнал вирусологии . 79 (16): 10180–9. doi : 10.1128/JVI.79.16.10180-10189.2005 . ПМК 1182666 . ПМИД 16051811 .
- ^ Перейти обратно: а б Драгоценный Б., Чайлдс К., Фитцпатрик-Сваллоу В., Гудборн С., Рэндалл Р.Э. (ноябрь 2005 г.). «Белок 5 V обезьяньего вируса действует как адаптер, связывая DDB1 со STAT2, чтобы облегчить убиквитинирование STAT1» . Журнал вирусологии . 79 (21): 13434–41. doi : 10.1128/JVI.79.21.13434-13441.2005 . ПМЦ 1262611 . ПМИД 16227264 .
- ^ МакКолл СМ, Милиани де Марваль П.Л., Честейн П.Д., Джексон С.С., Хэ Ю.Дж., Котаке Ю., Кук Дж.Г., Сюн Ю. (сентябрь 2008 г.). «Vpr-связывающий белок VprBP вируса иммунодефицита человека типа 1, белок WD40, связанный с убиквитинлигазой DDB1-CUL4 E3, необходим для репликации ДНК и эмбрионального развития» . Молекулярная и клеточная биология . 28 (18): 5621–33. дои : 10.1128/MCB.00232-08 . ПМК 2546929 . ПМИД 18606781 .
- ^ Ле Рузик Э., Белайдуни Н., Эстрабо Э., Морель М., Рейн Х.К., Транси С., Марготен-Гоге Ф. (январь 2007 г.). «Vpr ВИЧ1 останавливает клеточный цикл, привлекая DCAF1/VprBP, рецептор убиквитинлигазы Cul4-DDB1» . Клеточный цикл . 6 (2): 182–8. дои : 10.4161/cc.6.2.3732 . ПМИД 17314515 .
- ^ Вэнь, Сяоюнь; Дуус, Карен М.; Фридрих, Томас Д.; Де Норонья, Карлос MC (2007). «Белок Vpr ВИЧ1 способствует остановке цикла G2-клеток путем взаимодействия с комплексом убиквитин-лигазы, содержащим DDB1 и Cullin4A, с использованием VprBP/DCAF1 в качестве адаптера» . Журнал биологической химии . 282 (37): 27046–27057. дои : 10.1074/jbc.M703955200 . ПМИД 17620334 .
- ^ Перейти обратно: а б Ан Дж., Ву Т., Новинс З., Герреро-Санторо Дж., Рапич-Отрин В., Гроненборн А.М. (ноябрь 2010 г.). «Vpr ВИЧ-1 загружает урацил-ДНК-гликозилазу-2 на DCAF1, субъединицу распознавания субстрата убиквитинлигазы E3 куллина 4A-кольца для протеасомо-зависимой деградации» . Журнал биологической химии . 285 (48): 37333–41. дои : 10.1074/jbc.M110.133181 . ПМЦ 2988339 . ПМИД 20870715 .
- ^ Перейти обратно: а б Вэнь X, Кейси Клоков Л., Некорчук М., Шарифи Х.Дж., де Норонья К.М. (2012). «Белок Vpr ВИЧ1 усиливает конститутивный DCAF1-зависимый оборот UNG2» . ПЛОС ОДИН . 7 (1): e30939. Бибкод : 2012PLoSO...730939W . дои : 10.1371/journal.pone.0030939 . ПМЦ 3265533 . ПМИД 22292079 .
- ^ Перейти обратно: а б Хофманн Х, Лог Э.К., Блох Н., Даддача В., Польский С.Б., Шульц М.Л., Ким Б., Ландау Н.Р. (декабрь 2012 г.). «Лентивирусный добавочный белок Vpx нацелен на деградацию SAMHD1 в ядре» . Журнал вирусологии . 86 (23): 12552–60. дои : 10.1128/JVI.01657-12 . ПМК 3497686 . ПМИД 22973040 .
- ^ Перейти обратно: а б Ан Дж., Хао С., Ян Дж., ДеЛюсия М., Меренс Дж., Ван С., Гроненборн А.М., Скоуронски Дж. (апрель 2012 г.). «Дополнительный фактор вирулентности вируса иммунодефицита ВИЧ/обезьян (SIV) Vpx загружает фактор рестрикции клетки-хозяина SAMHD1 на комплекс убиквитинлигазы E3 CRL4DCAF1» . Журнал биологической химии . 287 (15): 12550–8. дои : 10.1074/jbc.M112.340711 . ПМК 3321004 . ПМИД 22362772 .
- ^ Ито, Т.; Андо, Х.; Сузуки, Т.; Огура, Т.; Хотта, К.; Имамура, Ю.; Ямагучи, Ю.; Ханда, Х. (2010). «Идентификация основной мишени тератогенности талидомида». Наука 327 (5971): 1345–1350. Бибкод : 2010Науч... 327.1345I дои : 10.1126/science.1177319 . ПМИД 20223979 . S2CID 17575104 .
- ^ Перейти обратно: а б с Лу Дж., Миддлтон Р.Э., Сан Х., Нанионг М., Отт СиДжей, Мициадес К.С., Вонг К.К., Брэднер Дж.Э., Кэлин В.Г. младший (2014). «Лекарственное средство от миеломы леналидомид способствует цереблон-зависимому разрушению белков Ikaros» . Наука . 343 (6168): 305–9. Бибкод : 2014Sci...343..305L . дои : 10.1126/science.1244917 . ПМК 4070318 . ПМИД 24292623 .
- ^ Перейти обратно: а б с Крёнке Дж., Удеши Н.Д., Нарла А., Грауман П., Херст С.Н., МакКонки М., Свинкина Т., Хекл Д., Комер Е., Ли Х, Сиарло С., Хартман Э., Мунши Н., Шеноне М., Шрайбер С.Л., Карр С.А., Эберт Б.Л. (2014). «Леналидомид вызывает избирательную деградацию IKZF1 и IKZF3 в клетках множественной миеломы» . Наука . 343 (6168): 301–5. Бибкод : 2014Sci...343..301K . дои : 10.1126/science.1244851 . ПМК 4077049 . ПМИД 24292625 .
- ^ Шиянов П., Наг А., Райчаудхури П. (декабрь 1999 г.). «Кулин 4А связывается с поврежденным УФ-излучением ДНК-связывающим белком DDB» . Журнал биологической химии . 274 (50): 35309–12. дои : 10.1074/jbc.274.50.35309 . ПМИД 10585395 .
- ^ Диас Д.К., Долиос Дж., Ван Р., Пан ZQ (декабрь 2002 г.). «CUL7: Куллин, содержащий домен DOC, избирательно связывает Skp1.Fbx29 с образованием SCF-подобного комплекса» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (26): 16601–6. дои : 10.1073/pnas.252646399 . ПМК 139190 . ПМИД 12481031 .
- ^ Охта Т., Мишель Дж.Дж., Шоттелиус А.Дж., Сюн Ю. (апрель 1999 г.). «ROC1, гомолог APC11, представляет собой семейство партнеров куллина со связанной активностью убиквитинлигазы» . Молекулярная клетка . 3 (4): 535–41. дои : 10.1016/s1097-2765(00)80482-7 . ПМИД 10230407 . S2CID 19371828 .
- ^ Мин К.В., Хван Дж.В., Ли Дж.С., Пак Ю, Тамура Т.А., Юн Дж.Б. (май 2003 г.). «TIP120A связывается с куллинами и модулирует активность убиквитинлигазы» . Журнал биологической химии . 278 (18): 15905–10. дои : 10.1074/jbc.M213070200 . ПМИД 12609982 .
- ^ Серино Дж., Дэн XW (2003). «Сигналосома COP9: регулирование развития растений посредством контроля протеолиза» (PDF) . Ежегодный обзор биологии растений . 54 : 165–82. doi : 10.1146/annurev.arplant.54.031902.134847 . ПМИД 14502989 . Архивировано из оригинала (PDF) 19 января 2015 г. Проверено 18 января 2015 г.
- ^ Чен X, Чжан Ю, Дуглас Л, Чжоу П (декабрь 2001 г.). «ДНК-связывающие белки, поврежденные УФ-излучением, являются мишенями убиквитинирования и деградации, опосредованного CUL-4A» . Журнал биологической химии . 276 (51): 48175–82. дои : 10.1074/jbc.M106808200 . ПМИД 11673459 .
- ^ Нишитани Х., Сиоми Ю., Иида Х., Мичишита М., Таками Т., Цуримото Т. (октябрь 2008 г.). «Ингибитор CDK p21 разлагается по пути Cul4-DDB1Cdt2, связанному с ядерным антигеном пролиферирующих клеток, во время S-фазы и после УФ-облучения» . Журнал биологической химии . 283 (43): 29045–52. дои : 10.1074/jbc.M806045200 . ПМК 2662008 . ПМИД 18703516 .
Внешние ссылки
[ редактировать ]- человека Расположение генома CUL4A и CUL4A страница сведений о гене в браузере генома UCSC .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q13619 (Cullin-4A) в PDBe-KB .
Дальнейшее чтение
[ редактировать ]- Осака Ф, Кавасаки Х, Аида Н, Саеки М, Чиба Т, Кавасима С, Танака К, Като С (август 1998 г.). «Новая система лигирования NEDD8 для куллина-4А» . Гены и развитие . 12 (15): 2263–8. дои : 10.1101/gad.12.15.2263 . ПМК 317039 . ПМИД 9694792 .
- Чен Л.К., Манджешвар С., Лу Ю., Мур Д., Люнг Б.М., Куо В.Л., Дайркей Ш., Верник М., Коллинз С., Смит Х.С. (август 1998 г.). «Человеческий гомолог гена Caenorhabditis elegans cul-4 амплифицируется и сверхэкспрессируется при первичном раке молочной железы». Исследования рака . 58 (16): 3677–83. ПМИД 9721878 .
- Охта Т., Мишель Дж.Дж., Шоттелиус А.Дж., Сюн Ю. (апрель 1999 г.). «ROC1, гомолог APC11, представляет собой семейство партнеров куллина со связанной активностью убиквитинлигазы» . Молекулярная клетка . 3 (4): 535–41. дои : 10.1016/S1097-2765(00)80482-7 . ПМИД 10230407 . S2CID 19371828 .
- Хори Т., Осака Ф., Чиба Т., Миямото С., Окабаяши К., Симбара Н., Като С., Танака К. (ноябрь 1999 г.). «Ковалентная модификация всех членов белков семейства куллинов человека с помощью NEDD8» . Онкоген . 18 (48): 6829–34. дои : 10.1038/sj.onc.1203093 . ПМИД 10597293 .
- Ляпина С., Коуп Г., Шевченко А., Серино Г., Цуге Т., Чжоу С., Вольф Д.А., Вэй Н., Шевченко А., Деше Р.Дж. (май 2001 г.). «Стимулирование расщепления конъюгата NEDD-CUL1 сигналосомой COP9». Наука . 292 (5520): 1382–5. Бибкод : 2001Sci...292.1382L . дои : 10.1126/science.1059780 . ПМИД 11337588 . S2CID 14224920 .
- Чен X, Чжан Ю, Дуглас Л, Чжоу П (декабрь 2001 г.). «ДНК-связывающие белки, поврежденные УФ-излучением, являются мишенями убиквитинирования и деградации, опосредованного CUL-4A» . Журнал биологической химии . 276 (51): 48175–82. дои : 10.1074/jbc.M106808200 . ПМИД 11673459 .
- Ясуи К., Арии С., Чжао С., Имото И., Уэда М., Нагаи Х., Эми М., Инадзава Дж. (июнь 2002 г.). «TFDP1, CUL4A и CDC16 идентифицированы как мишени для амплификации 13q34 при гепатоцеллюлярной карциноме» . Гепатология . 35 (6): 1476–84. дои : 10.1053/jhep.2002.33683 . ПМИД 12029633 .
- Лю Дж., Фурукава М., Мацумото Т., Сюн Ю. (декабрь 2002 г.). «Модификация CUL1 NEDD8 диссоциирует p120 (CAND1), ингибитор связывания CUL1-SKP1 и лигаз SCF» . Молекулярная клетка . 10 (6): 1511–8. дои : 10.1016/S1097-2765(02)00783-9 . ПМИД 12504025 . S2CID 28959307 .
- Мин К.В., Хван Дж.В., Ли Дж.С., Пак Ю, Тамура Т.А., Юн Дж.Б. (май 2003 г.). «TIP120A связывается с куллинами и модулирует активность убиквитинлигазы» . Журнал биологической химии . 278 (18): 15905–10. дои : 10.1074/jbc.M213070200 . ПМИД 12609982 .
- Гройсман Р., Полановска Дж., Кураока И., Савада Дж., Сайджо М., Драпкин Р., Киселев А.Ф., Танака К., Накатани Ю. (май 2003 г.). «Активность убиквитинлигазы в комплексах DDB2 и CSA по-разному регулируется сигналосомой COP9 в ответ на повреждение ДНК» . Клетка . 113 (3): 357–67. дои : 10.1016/S0092-8674(03)00316-7 . ПМИД 12732143 . S2CID 11639677 .
- Хига Л.А., Михайлов И.С., Бэнкс Д.П., Чжэн Дж., Чжан Х. (ноябрь 2003 г.). «Радиационно-опосредованный протеолиз CDT1 комплексами CUL4-ROC1 и CSN представляет собой новую контрольную точку». Природная клеточная биология . 5 (11): 1008–15. дои : 10.1038/ncb1061 . ПМИД 14578910 . S2CID 26070877 .
- Вертц И.Е., О'Рурк К.М., Чжан З., Дорнан Д., Арнотт Д., Деше Р.Дж., Диксит В.М. (февраль 2004 г.). «Человеческий деэтиолированный-1 регулирует c-Jun путем сборки убиквитинлигазы CUL4A» (PDF) . Наука . 303 (5662): 1371–4. Бибкод : 2004Sci...303.1371W . дои : 10.1126/science.1093549 . ПМИД 14739464 . S2CID 40501515 .
- Обусе С., Ян Х., Нозаки Н., Гото С., Оказаки Т., Йода К. (февраль 2004 г.). «Протеомный анализ центромерного комплекса интерфазных клеток HeLa: поврежденный УФ-излучением ДНК-связывающий белок 1 (DDB-1) является компонентом CEN-комплекса, тогда как ИМТ-1 временно колокализуется с центромерной областью в интерфазе» . Гены в клетки . 9 (2): 105–20. дои : 10.1111/j.1365-2443.2004.00705.x . ПМИД 15009096 . S2CID 21813024 .
- Ху Дж., МакКолл СМ, Охта Т., Сюн Ю. (октябрь 2004 г.). «Направленное убиквитинирование CDT1 лигазой DDB1-CUL4A-ROC1 в ответ на повреждение ДНК». Природная клеточная биология . 6 (10): 1003–9. дои : 10.1038/ncb1172 . ПМИД 15448697 . S2CID 34194616 .
- Наг А., Багчи С., Райчаудхури П. (ноябрь 2004 г.). «Cul4A физически связывается с MDM2 и участвует в протеолизе р53» . Исследования рака . 64 (22): 8152–5. doi : 10.1158/0008-5472.CAN-04-2598 . ПМИД 15548678 .
- Мацуда Н., Азума К., Сайдзё М., Иемура С., Хиоки Ю., Нацуме Т., Тиба Т., Танака К., Танака К. (май 2005 г.). «DDB2, продукт гена пигментной ксеродерма группы E, непосредственно убиквитилируется комплексом убиквитинлигазы на основе Cullin 4A». Восстановление ДНК . 4 (5): 537–45. дои : 10.1016/j.dnarep.2004.12.012 . ПМИД 15811626 .
галерея PDB |
|---|

