Аллостерическая регуляция

В областях биохимии и фармакологии аллостерический регулятор (или аллостерический модулятор ) - это вещество, которое связывается с сайтом на ферменте или рецепторе , отличном от активного сайта , что приводит к конформационному изменению, которое изменяет активность белка, либо усиливая, либо ингибируя его функция Напротив, вещества, которые связываются непосредственно с активным сайтом фермента или сайтом связывания эндогенного лиганда рецептора, называются ортостерическими регуляторами или модуляторами.
Сайт, с которым связывает эффектор, называется аллостерическим сайтом или регуляторным участком . Аллостерические сайты позволяют эффекторам связываться с белком, часто приводя к конформационному изменению и/или изменению динамики белка . [ 1 ] [ 2 ] Эффекторы, которые усиливают активность белка, называются аллостерическими активаторами , тогда как те, которые снижают активность белка, называются аллостерическими ингибиторами .
Аллостерические правила являются естественным примером управляющих петлей, таких как обратная связь из нисходящих продуктов или прямочная установка от подложков вверх по течению. Алсостерия на большие расстояния особенно важна для передачи сигналов клеток . [ 3 ] Аллостерическая регуляция также особенно важна для способности клетки регулировать активность фермента .
Термин «аллостерия» происходит от древнегреческого аллос ( ἄλλος ), «Другое» и стереосистема ( στερεός ), «твердый (объект)». Это относится к тому факту, что регуляторный сайт аллостерического белка физически отличается от его активного сайта. Аллостерия контрастирует с презентацией субстрата , которая не требует конформационных изменений для активации фермента. Термин «Ортостерия» происходит от древнегреческого ортонов ( ὀρθός ), означающего «прямо», «вертикально», «правильно» или «правильный».
Орто против аллостерических ингибиторов
[ редактировать ]Ортостерический
[ редактировать ]- Сайт связывания: ортостерические ингибиторы связываются непосредственно с активным сайтом фермента, где субстрат обычно связывается.
- Механизм действия: занимая активное место, эти ингибиторы предотвращают связывание субстрата, тем самым непосредственно блокируя каталитическую активность фермента.
- Конкурентное торможение: большинство ортостерических ингибиторов конкурируют с субстратом для активного сайта, что означает, что их эффективность может быть снижена, если концентрация субстрата увеличивается.
Аллостерический
[ редактировать ]- Сайт связывания: аллостерические ингибиторы связываются с сайтом на ферменте, который отличается и отделен от активного сайта, известного как аллостерический сайт.
- Механизм действия: связывание с аллостерическим сайтом вызывает конформационное изменение фермента, которое может либо уменьшить сродство активного сайта для субстрата, либо изменить каталитическую активность фермента. Это косвенное помех может ингибировать функцию фермента, даже если присутствует субстрат.
- Неконкурентное ингибирование: аллостерические ингибиторы часто демонстрируют неконкурентное ингибирование, что означает, что их ингибирующий эффект не зависит от концентрации субстрата.
Модели
[ редактировать ]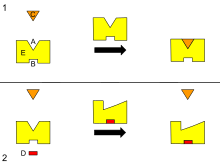
B - аллостерический сайт
C - субстрат
D - ингибитор
E - фермент
Это диаграмма аллостерической регуляции фермента.
Многие аллостерические эффекты могут быть объяснены согласованной моделью MWC, выдвинутой Monod , Wyman и Changeux , [ 4 ] или последовательной моделью (также известной как модель KNF), описанная Кошлендом , Немети и фильмом. [ 5 ] Обе постулируют, что белковые субъединицы существуют в одной из двух конформаций , напряженных (t) или расслабленных (R), и что расслабленные субъединицы связывают субстрат более легко, чем в напряженном состоянии. Эти две модели больше всего различаются по своим предположениям о взаимодействии субъединиц и расторжения обоих состояний. Для белков, в которых субъединицы существуют в более чем двух конформациях , модель ландшафта аллостерии, описанная Cuendet, Weinstein и Levine, [ 6 ] можно использовать. Аллестетическая регуляция может способствовать эволюции крупномасштабных, низкоэнергетических конформационных изменений, что позволяет долгоегическому аллостерическому взаимодействию между отдаленными сайтами связывания. [ 7 ]
Согласованная модель
[ редактировать ]Согласованная модель аллостерии, также называемая моделью симметрии или модели MWC , постулаты, которые субъединиц ферментов соединяются таким образом, что конформационное изменение в одной субъединице обязательно передается всем другим субъединицам. Таким образом, все субъединицы должны существовать в той же конформации. Модель также утверждает, что в отсутствие какого -либо лиганда (субстрата или иного) равновесие способствует одному из конформационных состояний, T или R. Равновесие может быть перенесено в состояние R или T через связывание одного лиганда ( аллостерический эффектор или лиганд) на сайт, который отличается от активного сайта
Последовательная модель
[ редактировать ]Последовательная модель аллостерической регуляции гласит, что субъединицы не связаны таким образом, что конформационное изменение в одном вызывает аналогичное изменение в других. Таким образом, все субъединицы фермента не требуют той же конформации. Более того, последовательная модель диктует, что молекулы субстрата связывают с помощью индуцированного протокола соответствия . В то время как такая индуцированная посадка преобразует субъединицу из затянутого состояния в расслабленное состояние, она не распространяет конформационные изменения в соседних субъединицах. Вместо этого субстрат связывание в одной субъединице лишь слегка изменяет структуру других субъединиц, так что их сайты связывания более восприимчивы к субстрату. Суммировать:
- субъединицы не должны существовать в той же конформации
- Молекулы субстрата связываются с помощью протокола индуцированного соответствия
- Конформационные изменения не распространяются на все субъединицы
Морфейн модель
[ редактировать ]Модель аллостерической регуляции Morpheein является диссоциативной согласованной моделью. [ 8 ]
Морфиин-это гомоолигомерная структура, которая может существовать в качестве ансамбля физиологически значимых и функционально разных альтернативных четвертичных сборов. Переходы между альтернативными сборками морфиина включают диссоциацию олигомеров, конформационные изменения в диссоциированном состоянии и повторную сборку к другому олигомеру. Требуемый шаг разборки олигомеров отличает модель Morpheein для аллостерической регуляции от классических моделей MWC и KNF.
Porphobilinogen Synthase (PBG) является прототипом Morpheein.
Ансамблевые модели
[ редактировать ]аллостерической системы Ансамблевые модели аллостерической регуляции перечисляют статистический ансамбль в зависимости от его функции потенциальной энергии , а затем связывают специфические статистические измерения аллостерии с определенными терминами энергии в энергетической функции (например, межмолекулярное солодное мост между двумя доменами). [ 9 ] Ансамблевые модели, такие как ансамбль аллостерическая модель [ 10 ] и аллостерическая модель [ 11 ] Предположим, что каждый домен системы может принять два состояния, аналогичные модели MWC. Модель ландшафта аллостерии, представленная Cuendet, Weinstein и Levine [ 6 ] Позволяет домену иметь любое количество состояний, и вклад конкретного молекулярного взаимодействия в данную аллостерическую связь может быть оценена с использованием строгого набора правил. Моделирование молекулярной динамики может быть использовано для оценки статистического ансамбля системы, чтобы его можно было проанализировать с помощью модели ландшафта аллостерии.
Аллостерическая модуляция
[ редактировать ]Аллостерическая модуляция используется для изменения активности молекул и ферментов в биохимии и фармакологии. Для сравнения производится типичный препарат, чтобы связываться с активным сайтом фермента, который, таким образом, запрещает связывание субстрата с тем ферментом, вызывающим снижение активности фермента. Аллостерическая модуляция возникает, когда эффектор связывается с аллостерическим сайтом (также известным как регуляторный сайт) фермента и изменяет активность фермента. Аллостерические модуляторы предназначены для того, чтобы соответствовать аллостерическому участку, чтобы вызвать конформационное изменение фермента, в частности изменение формы активного сайта, которое затем вызывает изменение его активности. В отличие от типичных лекарств, модуляторы не являются конкурентными ингибиторами . Они могут быть положительными (активирующими), вызывая повышение активности фермента или отрицательное (ингибирование), вызывая снижение активности фермента. Использование аллостерической модуляции позволяет контролировать эффекты специфических ферментов; В результате аллостерические модуляторы очень эффективны в фармакологии. [ 12 ] В биологической системе аллостерическая модуляция может быть трудно отличить от модуляции путем презентации субстрата .
Модель энергии
[ редактировать ]Пример этой модели можно увидеть с помощью Mycobacterium Tuberculosis , бактерии , которая идеально подходит для адаптации к жизни в макрофагах людей. Сайты фермента служат связью между различными субстратами. В частности, между AMP и G6P . Подобные сайты также служат чувствительным механизмом для эффективности фермента. [ 13 ]
Положительная модуляция
[ редактировать ]Положительная аллостерическая модуляция (также известная как аллостерическая активация ) происходит, когда связывание одного лиганда усиливает притяжение между молекулами субстрата и другими сайтами связывания. Примером является связывание молекул кислорода с гемоглобином , где кислород является как субстрат , так и эффектор. Аллостерический, или «другой», сайт является активным сайтом соседней белковой субъединицы . Связывание кислорода с одной субъединицей вызывает конформационное изменение в той субъединице, которая взаимодействует с оставшимися активными участками, чтобы усилить их сродство кислорода. Другой пример аллостерической активации наблюдается в цитозольном IMP-GMP, специфичной 5'-нуклеотидазе II (CN-II), где аффинность к субстрату GMP увеличивается при связывании GTP на границе раздела димер.
Отрицательная модуляция
[ редактировать ]Отрицательная аллостерическая модуляция (также известная как аллостерическое ингибирование ) происходит, когда связывание одного лиганда уменьшает сродство к субстрату в других активных сайтах. Например, когда 2,3-BPG связывается с аллостерическим сайтом на гемоглобине, сродство к кислороду всех субъединиц уменьшается. Это когда регулятор отсутствует в сайте связывания.
Прямые ингибиторы тромбина дают отличный пример отрицательной аллостерической модуляции. аллостерические ингибиторы Были обнаружены тромбина, которые могут быть использованы в качестве антикоагулянтов.
Другим примером является Strychnine , судорожный яд, который действует как аллостерический ингибитор рецептора глицина . Глицин является основным пост- синаптическим ингибирующим нейротрансмиттером в млекопитающих спинном мозге и стебле мозга . Стрихнин действует на отдельный сайт связывания на рецепторе глицина аллостерическим образом; т.е. его связывание снижает аффинность глицинового рецептора к глицину. Таким образом, стрихнин ингибирует действие ингибирующего передатчика, что приводит к судороги.
Еще один случай, в котором можно увидеть отрицательную аллостерическую модуляцию, заключается между АТФ и ферментом фосфофруктокиназы в цикле отрицательной обратной связи , которая регулирует гликолиз . Фосфофруктокиназа (обычно называемая PFK ) является ферментом, который катализирует третий этап гликолиза: фосфорилирование фруктозо -6-фосфата в фруктозу 1,6-бисфосфат . PFK может быть аллостерически ингибирована высокими уровнями АТФ в клетке. Когда уровни АТФ высоки, АТФ будет связываться с аллостерическим сайтом на фосфофруктокиназе , вызывая изменение трехмерной формы фермента. Это изменение заставляет его аффинность к субстрату ( фруктоза-6-фосфат и АТФ ) в активном участке для уменьшения, а фермент считается неактивным. Это заставляет гликолиз организма прекращать, когда уровни АТФ высоки, тем самым сохраняя глюкозу и поддерживая сбалансированные уровни клеточного АТФ. Таким образом, ATP служит негативным аллостерическим модулятором для PFK, несмотря на то, что он также является субстратом фермента.
Типы
[ редактировать ]Гомотропный
[ редактировать ]Гомотропный аллостерический модулятор является субстратом для его целевого белка , а также регуляторной молекулой активности белка. Обычно это активатор белка. [ 14 ] Например, O 2 и CO являются гомотропными аллостерическими модуляторами гемоглобина. Аналогичным образом, в специфической 5 'нуклеотидазе IMP/GMP связывание одной молекулы GMP с одной субъединицей тетрамерного фермента приводит к повышению сродства к GMP последующими субъединицами, выявленными Sigmoidal Substrate по сравнению с уровнями скорости. [ 14 ]
Гетеротропный
[ редактировать ]Гетеротропный аллостерический модулятор - это регуляторная молекула, которая не является субстратом фермента. Это может быть либо активатор, либо ингибитор фермента. Например, h + , CO 2 и 2,3-бисфосфоглицерат являются гетеротропными аллостерическими модуляторами гемоглобина. [ 15 ] Еще раз, в специфической 5-'нуклеотидазе IMP/GMP связывание молекулы GTP на границе с димером в тетрамерном ферменте приводит к повышению аффинности к субстрату GMP в активном сайте, указывающем на гетеротропную активацию аллостерической активации K-типа. [ 14 ]
Как было достаточно выделено выше, некоторые аллостерические белки могут регулироваться как их субстратами, так и другими молекулами. Такие белки способны как гомотропные, так и гетеротропные взаимодействия. [ 14 ]
Основные активаторы
[ редактировать ]Некоторые аллостерические активаторы называются «существенными» или «облигационными» активаторами, в том смысле, что в их отсутствие активность их целевого ферментативного фермента является очень низкой или незначительной, как и в случае с активностью N-ацетилглутамата на карбамоил фосфат Синтетаза I, например. [ 16 ] [ 17 ]
Нерегуляторная аллостера
[ редактировать ]Нерегуляторным аллостерическим сайтом является какой-либо нерегуляторный компонент фермента (или любого белка), который сам не является аминокислотой. Например, многие ферменты требуют связывания натрия для обеспечения правильной функции. Однако натрий не обязательно действует как регулирующая субъединица; Натрий всегда присутствует, и нет известных биологических процессов для добавления/удаления натрия для регулирования активности ферментов. Нерегуляторная аллостерия может включать любые другие ионы, помимо натрия (кальций, магний, цинк), а также другие химические вещества и, возможно, витамины.
Фармакология
[ редактировать ]Аллостерическая модуляция рецептора является результатом связывания аллостерических модуляторов на другом сайте (« регуляторный сайт ») от эндогенного лиганда (« активный сайт ») и усиливает или ингибирует эффекты эндогенного лиганда. При нормальных обстоятельствах он действует, вызывая конформационное изменение молекулы рецептора, что приводит к изменению аффинности связывания лиганда. Таким образом, аллостерический лиганд модулирует активацию рецептора своим первичным ортостерическим лигандом, и можно считать, что он действует как диммерный переключатель в электрической цепи, регулируя интенсивность отклика.
Например, GABA A рецептор имеет два активных участка, которые связывается с нейротрансмиттером гамма-аминобутирической кислотой (ГАМК), но также имеет бензодиазепина и общего анестезиозного агента сайты регуляторного связывания . Эти регуляторные сайты могут производить положительную аллостерическую модуляцию, способствуя активности ГАМК. Diazepam является положительным аллостерическим модулятором в регуляторном месте бензодиазепина, а его противоядие флумазенил является антагонистом рецептора .
Более поздние примеры лекарств, которые аллостерически модулируют их цели, включают в себя кальций, имитирующий Cinacalcet лечение ВИЧ и Maraviroc, .
Аллостерические участки как лекарственные цели
[ редактировать ]Аллостерические белки участвуют и являются центральными во многих заболеваниях, [ 18 ] [ 19 ] и аллостерические сайты могут представлять собой новую цель лекарств . Существует ряд преимуществ при использовании аллостерических модуляторов в качестве предпочтительных терапевтических агентов по сравнению с классическими ортостерическими лигандами. Например, аллостерического связывания , связанные с белком (GPCR) сайты . [ 20 ] Следовательно, большая селективность GPCR может быть получена путем нацеливания на аллостерические сайты. [ 20 ] Это особенно полезно для GPCR, где селективная ортостерическая терапия была затруднена из -за сохранения последовательностей ортостерического сайта в подтипах рецепторов. [ 21 ] Кроме того, эти модуляторы имеют снижение потенциала для токсических эффектов, поскольку модуляторы с ограниченной кооперативностью будут иметь уровень потолка для их эффекта, независимо от введенной дозы. [ 20 ] Другой тип фармакологической селективности, которая является уникальной для аллостерических модуляторов, основан на кооперативности. Аллостерический модулятор может отображать нейтральную кооперативность с ортостерическим лигандом во всех подтипах данного рецептора, за исключением интересующего подтипа, который называется «абсолютная селективность подтипа». [ 21 ] Если аллостерический модулятор не обладает заметной эффективностью, он может обеспечить еще одно мощное терапевтическое преимущество по сравнению с ортостерическими лигандами, а именно способность выборочно настраивать или вниз тканевые реакции только в том случае, когда присутствует эндогенный агонист. [ 21 ] Специфичные для олигомер сайты связывания малых молекул являются лекарственными целями для медицинских с медицинской точки зрения морфии . [ 22 ]
Синтетические аллостерические системы
[ редактировать ]Существует много синтетических соединений, содержащих несколько нековалентных сайтов связывания, которые демонстрируют конформационные изменения при занятии одного сайта. Кооперативность между отдельными вкладами связывания в таких супрамолекулярных системах является положительной, если занятие одного сайта связывания усиливает аффинность Δ G на втором месте и отрицательно, если сродство не повышено. Большинство синтетических аллостерических комплексов полагаются на конформационную реорганизацию на связывании одного эффекторного лиганда, который затем приводит к усилению или ослабленной ассоциации второго лиганда в другом сайте связывания. [ 23 ] [ 24 ] [ 25 ] Конформационная связь между несколькими сайтами связывания находится в искусственных системах, как правило, намного больше, чем в белках с их обычно большей гибкостью. Параметр, который определяет эффективность (как измеряется по отношению равновесных константов krel = ka (e)/ka в присутствии и отсутствии эффекторной эпохи), является конформационной энергией, необходимой для принятия закрытой или напряженной конформации для связывания лиганда А. [ 26 ]
Во многих многовалентных супрамолекулярных системах [ 27 ] Может возникнуть прямое взаимодействие между связанными лигандами, что может привести к большой кооперативности. Наиболее распространенным является такое прямое взаимодействие между ионами в рецепторах для ионных пар. [ 28 ] [ 29 ] Эта кооперативность часто также называется аллостерией, хотя конформационные изменения здесь не обязательно вызывают события связывания.
Онлайн -ресурсы
[ редактировать ]Аллостерическая база данных
[ редактировать ]Аллостерия является прямым и эффективным средством регуляции биологической макромолекулярной функции, создаваемой связыванием лиганда на аллостерическом участке топографически отличается от ортостерического сайта. Из-за часто высокой селективности рецепторов и более низкой токсичности на основе мишеней также ожидается, что аллостерическая регуляция будет играть растущую роль в обнаружении лекарств и биоинженерии. Аллостерическая база данных (ASD) [ 30 ] Обеспечивает центральный ресурс для отображения, поиска и анализа структуры, функции и связанных с ним аннотаций для аллостерических молекул. В настоящее время ASD содержит аллостерические белки из более чем 100 видов и модуляторов в трех категориях (активаторы, ингибиторы и регуляторы). Каждый белок аннотирован с подробным описанием аллостерии, биологического процесса и связанных с ними заболеваний, а также каждый модулятор с аффинностью связывания, физико -химическими свойствами и терапевтической областью. Интеграция информации аллостерических белков в ASD должна позволять прогнозированию аллостерии для неизвестных белков, следовать экспериментальной проверке. Кроме того, модуляторы, курируемые в ASD, могут использоваться для исследования потенциальных аллостерических целей для соединения запроса, и могут помочь химикам реализовать структурные модификации для нового аллостерического дизайна лекарств.
Аллостерические остатки и их прогноз
[ редактировать ]Не все остатки белка играют одинаково важную роль в аллостерической регуляции. Идентификация остатков, которые необходимы для аллостерии (так называемые «аллостерические остатки») были в центре внимания многих исследований, особенно в течение последнего десятилетия. [ 31 ] [ 32 ] [ 33 ] [ 34 ] [ 35 ] [ 36 ] [ 37 ] [ 38 ] Частично этот растущий интерес является результатом их общего значения в белковой науке, а также потому, что аллостерические остатки могут использоваться в биомедицинских контекстах . Фармакологически важные белки с трудными для нацеливания сайтов могут привести к подходам, при которых альтернативно нацелены на более легкие остатки, которые способны аллостерически регулировать первичный сайт, представляющий интерес. [ 39 ] Эти остатки в целом можно классифицировать как поверхностные и внутренние а аминокислоты. Аллостерические участки на поверхности, как правило, играют регуляторные роли, которые в основном отличаются от мест внутри внутренних дел; Поверхностные остатки могут служить рецепторами или эффекторными сайтами при трансмиссии аллостерического сигнала, тогда как те, кто внутри внутренних, может действовать для передачи таких сигналов. [ 40 ] [ 41 ]
Смотрите также
[ редактировать ]- База данных ASD
- Ангармония
- Конкурентное запрещение
- Кооперативное связывание
- Кинетика фермента
- Протеиновая динамика
- Теория рецепторов
Ссылки
[ редактировать ]- ^ Купер А, Драйден Д.Т. (октябрь 1984 г.). «Алсостера без конформационных изменений. Вероятная модель». Европейский журнал биофизики . 11 (2): 103–109. doi : 10.1007/bf002766625 . PMID 6544679 . S2CID 12591175 .
- ^ Лю Дж., Нуссинови Р. (июнь 2016 г.). «Аллостера: обзор его истории, концепций, методов и приложений» . PLOS Computational Biology . 12 (6): E1004966. BIBCODE : 2016PLSCB..12E4966L . doi : 10.1371/journal.pcbi.1004966 . PMC 4890769 . PMID 27253437 . S2CID 3610740 .
- ^ Bu Z, Callaway DJ (2011). «Протеины движутся! Протеиновая динамика и долгосрочная аллостерия в клеточной передаче». Структура белка и заболевания . Достижения в области химии белка и структурной биологии. Тол. 83. С. 163–221. doi : 10.1016/b978-0-12-381262-9.00005-7 . ISBN 9780123812629 Полем PMID 21570668 .
- ^ Monod J, Wyman J, Changeux JP (май 1965). «О природе аллостерических переходов: правдоподобная модель». Журнал молекулярной биологии . 12 : 88–118. doi : 10.1016/s0022-2836 (65) 80285-6 . PMID 14343300 .
- ^ Koshland DE, Némethy G, Filmer D (январь 1966 г.). «Сравнение экспериментальных данных связывания и теоретических моделей в белках, содержащих субъединицы». Биохимия . 5 (1): 365–85. doi : 10.1021/bi00865a047 . PMID 5938952 .
- ^ Jump up to: а беременный Cuendet MA, Weinstein H, Levine MV (декабрь 2016 г.). «Ландшафт аллостерии: количественная оценка термодинамических муфт в биомолекулярных системах» . Журнал химической теории и вычислений . 12 (12): 5758–5767. doi : 10.1021/acs.jctc.6b00841 . PMC 5156960 . PMID 27766843 .
- ^ Eckmann JP, Rougemont J, Tlusty T (30 июля 2019 г.). «Коллоквиум: белки: физика аморфного развивающегося материи» . Обзоры современной физики . 91 (3): 031001. Arxiv : 1907.13371 . BIBCODE : 2019RVMP ... 91C1001E . doi : 10.1103/revmodphys.91.031001 . ISSN 0034-6861 . S2CID 199001124 .
- ^ Jaffe EK (сентябрь 2005 г.). «Morpheeins-новая структурная парадигма для аллостерической регуляции». Тенденции в биохимических науках . 30 (9): 490–7. doi : 10.1016/j.tibs.2005.07.003 . PMID 16023348 .
- ^ Motlagh HN, Forabl Jo, Li J, Hilser VJ (апрель 2014). «Ансамблевая природа аллостерии» . Природа . 508 (7496): 331–9. Bibcode : 2014natur.508..331M . doi : 10.1038/nature13001 . PMC 4224315 . PMID 24740064 .
- ^ Hilser VJ, Forabl Jo, Motlagh HN (2012). «Структурная и энергетическая основа аллестеи» . Ежегодный обзор биофизики . 41 : 585–609. doi : 10.1146/annurev-biophys-050511-102319 . PMC 3935618 . PMID 22577828 .
- ^ Levine MV, Weinstein H (май 2015). «Цель аллостерии: использование модели ISING для понимания обработки и передачи информации в аллостерических биомолекулярных системах» . Энтропия . 17 (5): 2895–2918. BIBCODE : 2015Entrp..17.2895L . doi : 10.3390/e17052895 . PMC 4652859 . PMID 26594108 .
- ^ Abdel-Magid AF (февраль 2015 г.). «Аллостерические модуляторы: появляющаяся концепция в обнаружении наркотиков» . ACS лекарственная химия . 6 (2): 104–7. doi : 10.1021/ml5005365 . PMC 4329591 . PMID 25699154 .
- ^ Zhong W, Cui L, Goh BC, Cai Q, Ho P, Chionh YH, et al. (Декабрь 2017). «Аллостерическая пируваткиназа на основе« логических ворот »синергически определяет уровни энергии и сахара при туберкулезе Mycobacterium» . Природная связь . 8 (1): 1986. Bibcode : 2017natco ... 8.1986z . doi : 10.1038/s41467-017-02086-y . PMC 5719368 . PMID 29215013 .
- ^ Jump up to: а беременный в дюймовый Сринивасан Б., Фоухар Ф., Шукла А., Сампанги С., Кулкарни С., Абашидзе М. и др. (Март 2014 г.). «Аллостерская регуляция и активация субстрата в цитозольной нуклеотидазе II из легионелла пневмофила» . Журнал FEBS . 281 (6): 1613–1628. doi : 10.1111/febs.12727 . PMC 3982195 . PMID 24456211 .
- ^ Эдельштейн С.Дж. (1975). «Кооперативные взаимодействия гемоглобина». Ежегодный обзор биохимии . 44 : 209–32. doi : 10.1146/annurev.bi.44.070175.001233 . PMID 237460 .
- ^ Ши Д., Аллуэлл Н.М., Тучман М (июнь 2015 г.). «Семейство N-ацетилглютамат-синтазы: структуры, функции и механизмы» . Международный журнал молекулярных наук . 16 (6): 13004–22. doi : 10.3390/ijms160613004 . PMC 4490483 . PMID 26068232 .
- ^ De Cima S, Polo LM, Díez-Fernández C, Martínez Ai, Cervera J, Fita I, et al. (Ноябрь 2015). «Структура человеческой карбамоилфосфат -синтетазы: расшифровка переключателя в включении/выключении человеческого урегенеза» . Научные отчеты . 5 (1): 16950. Bibcode : 2015natsr ... 516950d . doi : 10.1038/srep16950 . PMC 4655335 . PMID 26592762 .
- ^ Nussinov R, Tsai C (2013). «Аллостера при болезни и в обнаружении наркотиков» . Клетка . 153 (2): 293–305. doi : 10.1016/j.cell.2013.03.034 . PMID 23582321 .
- ^ Abrusan G, Ascher DB, Inouye M (2022). «Известные аллостерические белки играют центральную роль в генетическом заболевании» . PLOS Computational Biology . 18 (2): E1009806. Arxiv : 2107.04318 . BIBCODE : 2022PLSCB..18E9806A . doi : 10.1371/journal.pcbi.1009806 . PMID 10138267 .
- ^ Jump up to: а беременный в Christopoulos A, May Lt, Avlani Va, Sexton PM (ноябрь 2004 г.). «Рецептор-рецептор, связанный с G-белком: обещание и проблема (ы)». Биохимическое общество транзакций . 32 (Pt 5): 873–7. doi : 10.1042/bst0320873 . PMID 15494038 .
- ^ Jump up to: а беременный в May Lt, Leach K, Sexton PM, Christopoulos A (2007). «Аллостерская модуляция рецепторов, связанных с белком,». Ежегодный обзор фармакологии и токсикологии . 47 : 1–51. doi : 10.1146/annurev.pharmtox.47.120505.105159 . PMID 17009927 .
- ^ Jaffe EK (2010). «Morpheeins-новый путь для аллостерических наркотиков ~! 2010-02-12 ~! 2010-05-21 ~! 2010-06-08 ~!» Полем Открытая конференция журнала . 1 : 1–6. doi : 10.2174/2210289201001010001 (неактивная 11 марта 2024 г.). PMC 3107518 . PMID 21643557 .
{{cite journal}}: CS1 Maint: doi неактивен с марта 2024 года ( ссылка ) - ^ Takeuchi M, Ikeda M, Sugasaki A, Shinkai S (ноябрь 2001 г.). «Молекулярный дизайн систем искусственного молекулярного и ионов распознавания с аллостерическими гостями». Счета химических исследований . 34 (11): 865–73. doi : 10.1021/ar0000410 . PMID 11714258 .
- ^ Кремер С, Люцен А (май 2013). «Искусственные аллостерические рецепторы». Химия: европейский журнал . 19 (20): 6162–96. doi : 10.1002/chem.201203814 . PMID 23463705 .
- ^ Kovbasyuk L, Krämer R (июнь 2004 г.). «Аллостерические супрамолекулярные рецепторы и катализаторы». Химические обзоры . 104 (6): 3161–87. doi : 10.1021/cr030673a . PMID 15186190 .
- ^ Schneider HJ (сентябрь 2016 г.). «Параметры эффективности в искусственных аллостерических системах». Органическая и биомолекулярная химия . 14 (34): 7994–8001. doi : 10.1039/c6ob01303a . PMID 27431438 .
- ^ Баджич Дж. Д., Нельсон А., Кантрилл С.Дж., Тернбулл В.Б., Стоддарт Дж.Ф. (сентябрь 2005 г.). «Многоваленность и кооперативность в супрамолекулярной химии». Счета химических исследований . 38 (9): 723–32. doi : 10.1021/ar040223k . PMID 16171315 .
- ^ Ким С.К., Сесслер Дж.Л. (октябрь 2010 г.). «Ионные парные рецепторы» . Обзоры химического общества . 39 (10): 3784–809. doi : 10.1039/c002694h . PMC 3016456 . PMID 20737073 .
- ^ McConnell AJ, Beer Pd (май 2012 г.). «Гетеродитопные рецепторы для распознавания ионной пары». Angewandte Chemie . 51 (21): 5052–61. doi : 10.1002/anie.201107244 . PMID 22419667 .
- ^ Huang Z, Zhu L, Cao Y, Wu G, Liu X, Chen Y, et al. (Январь 2011). «ASD: комплексная база данных аллостерических белков и модуляторов» . Исследование нуклеиновых кислот . 39 (проблема базы данных): D663–9. doi : 10.1093/nar/gkq1022 . PMC 3013650 . PMID 21051350 .
- ^ Panjkovich A, Daura X (октябрь 2012 г.). «Использование гибкости белка для прогнозирования местоположения аллостерических участков» . BMC Bioinformatics . 13 : 273. DOI : 10.1186/1471-2105-13-273 . PMC 3562710 . PMID 23095452 .
- ^ Süel GM, Lockless SW, Wall MA, Ranganathan R (январь 2003 г.). «Эволюционно консервативные сети остатков опосредуют аллостерическую связь в белках». Природа структурная биология . 10 (1): 59–69. doi : 10.1038/nsb881 . PMID 12483203 . S2CID 67749580 .
- ^ Mitternacht S, Berezovsky In (сентябрь 2011 г.). «Связывающее левередж как молекулярную основу для аллостерической регуляции» . PLOS Computational Biology . 7 (9): E1002148. BIBCODE : 2011PLSCB ... 7E2148M . doi : 10.1371/journal.pcbi.1002148 . PMC 3174156 . PMID 21935347 .
- ^ Гапер П.М., Фуглестад Б., Комивс Э.А., Марквик П.Р., Маккаммон Дж.А. (декабрь 2012 г.). «Аллостерические сети в тромбине различают прокоагулянт против антикоагулянтной активности» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (52): 21216–22. doi : 10.1073/pnas.1218414109 . PMC 3535651 . PMID 23197839 .
- ^ Гош А., Вишвешвара С (ноябрь 2008 г.). «Изменения в моделях клики и сообщества в структурах белков во время аллостерической связи: исследование динамически уравновешенных структур комплексов метионилл -тРНК -синтетазы». Биохимия . 47 (44): 11398–407. doi : 10.1021/bi8007559 . PMID 18842003 .
- ^ Sethi A, Eargle J, Black AA, Luthey-Schulten Z (апрель 2009 г.). «Динамические сети в тРНК: белковые комплексы» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (16): 6620–5. Bibcode : 2009pnas..106.6620S . doi : 10.1073/pnas.0810961106 . PMC 2672494 . PMID 19351898 .
- ^ Vanwart AT, Eargle J, Luthey-Schulten Z, Amaro Re (август 2012 г.). «Изучение вклада компонентов остатков в динамические сетевые модели аллостерии» . Журнал химической теории и вычислений . 8 (8): 2949–2961. doi : 10.1021/ct300377a . PMC 3489502 . PMID 23139645 .
- ^ Rivivalta I, Sultan MM, Lee NS, Manley GA, Loria JP, Batista Vs (май 2012 г.). «Аллостерические пути в имидазоле глицеринофосфат -синтазы» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (22): E1428–36. doi : 10.1073/pnas.1120536109 . PMC 3365145 . PMID 22586084 .
- ^ Негере К.Ф., Морзан У.Н., Хендриксон Х.П., Пал Р., Лиси Г.П., Лория Дж.П. и др. (Декабрь 2018). «Связочная центральность характеристики белковых аллостерических путей» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (52): E12201 - E12208. Arxiv : 1706.02327 . Bibcode : 2018pnas..11512201N . doi : 10.1073/pnas.1810452115 . PMC 6310864 . PMID 30530700 .
- ^ Clarke D, Sethi A, Li S, Kumar S, Chang RW, Chen J, et al. (Май 2016 г.). «Идентификация аллостерических горячих точек с динамикой: применение к сохранению между и внутривидовым» . Структура 24 (5): 826–837. doi : 10.1016/j.str.2016.03.008 . PMC 4883016 . PMID 2706750 .
- ^ Dutta S, Eckmann JP, Libchaber A, Tlusty T (май 2018). «Зеленая функция коррелированных генов в минимальной механической модели эволюции белка» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (20): E4559 - E4568. Arxiv : 1801.03681 . Bibcode : 2018pnas..115e4559d . doi : 10.1073/pnas.1716215115 . PMC 5960285 . PMID 29712824 .
Внешние ссылки
[ редактировать ]- Мгновенное понимание внедрения системы классификации для механизмов аллостерии белков из Королевского химического общества