Parareptilis
| Парарептилы | |
|---|---|

| |
| Коллаж из пяти окаменелостей парарептиля. стрелке сверху, мезозавр Tenuisens вероятный аклесторхинид сунсуза ) ( идентификатор , ( пареайайавр ( ), Nyctiphrures accudens a nyctiphruruths ) fenneri ( Карпински По часовой | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Рептилия |
| Клада : | † Parareptilis Олсон , 1947 |
| Приказ | |
Parareptilia «почти рептилии») является вымершим подклассом или кладой базальных ( сауропсидов / рептилий , обычно считающихся сестринским таксоном для Eureptilia (группа, которая, вероятно, содержит все живые рептилии и птицы). Парарептилы сначала возникли ближе к концу каменноугольного периода и достигли своего высшего разнообразия в течение пермского периода . Несколько экологических инноваций были впервые достигнуты парарептилами среди рептилий. К ним относятся первые рептилии, которые возвращаются в морские экосистемы ( мезозавры ), первые двуногими рептилии ( болосауриды, такие как Eudibamus ), первые рептилии с передовыми слуховыми системами ( Nycteroleteterids и другие) и первые крупные травоядные рептилии ( Pareiasaurs ). Единственными парарептилами, чтобы выжить в триасовый период, были проколофоноиды , группа мелких универсалов, всеядных и травоядных. Наибольшее семейство проколофоноидов, проколофонидов , перераспределенных в триасе, но впоследствии снизилось и вымерло к концу периода. [ 2 ] [ 3 ]
По сравнению с большинством эвриптилей, парарептилы сохранили довольно «примитивные» характеристики, такие как надежные, низкопрестанные тела и крупные надпорасорополисные кости в задней части черепа. В то время как все, кроме самых ранних эвриптилей, были диапсидами , с двумя отверстиями в задней части черепа, парарептилы, как правило, были более консервативными в степени временной фенестрации . В своем современном использовании Parareptilia впервые использовалась в качестве кладистистически правильной альтернативы Anapsida , термину, который исторически упоминал рептилии с твердыми черепами, не имеющими дыр за глазами. [ 4 ] Тем не менее, не у всех парарептилов есть «anapsid» черепа, а у некоторых есть большие отверстия в задней части черепа. У них также было несколько уникальных адаптаций, таких как большая яма на верхней челюсти , широкий префронтальный контакт , и отсутствие супраглоидного отверстия лопатки . [ 4 ] [ 5 ]
Как и многие другие так называемые «Anapsids», парарептилы были исторически изученными. Интерес к их отношениям был оживлен в 1990 -х годах, когда в нескольких исследованиях утверждалось, что тестонины ( черепахи и их родственники) были членами Parareptilia. [ 4 ] Хотя это предполагает, что парарептилия в конце концов не вымерла, происхождение черепах все еще широко обсуждается. Многие другие морфологические или генетические анализы находят большую поддержку черепах среди диапсидных эвриптилей, таких как сауроптерии или архозавраморфы , а не парарептили. [ 6 ] [ 7 ] [ 8 ] [ 3 ]
Описание
[ редактировать ]Череп
[ редактировать ]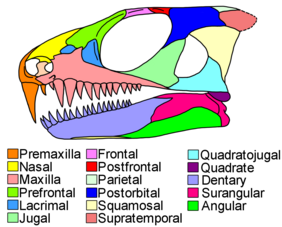
Парарептилианские черепа были разнообразными, от мезозавров с удлиненными мордами, заполненными сотнями тонких зубов, до сносительных носов, инкутированных на ручку черепа парейсавров . Парарептильные зубы были довольно изменчивыми по форме и функции между различными видами. Однако они были относительно однородными на том же черепе. В то время как большинство синапсидов и многих ранних эвриптилей имели канообразную область увеличенных клыков, похожих на зубы, в передней половине черепа, очень немногие парарептилы обладали канообразными зубами. [ 5 ]
Многие амниоты имеют ряд небольших ям, бегущих по костям на краю рта, но у парарептилов есть всего несколько ям, с одной особенно большой ямой рядом с передней частью верхней челюсти . [ 4 ] [ 9 ] [ 7 ] Остальная часть черепа часто была сильно текстурирована ямками, хребтами и грузчиками в большинстве парарептильных групп, иногда кульминационными в сложных боссах или шипах. Верхняя челюсть обычно низкая, в то время как префронтальные и слезные кости перед глазами довольно большие. Во всех парарептилях, кроме мезозавров, префронталь имеет внутреннюю ветвь, похожую на пластинку, которая образует широкий контакт с неважной костью неба . [ 4 ] [ 7 ] [ 5 ] Выдающаяся дыра, отверстие Orbitonasale, присутствует на пересечении префронтального, лаконичного и слезного. Парарептилианские вкусы также имеют беззубые и уменьшенные эктоптеригоидные кости, состояние, доведенное до крайности в мезозаврах, которые полностью потеряли эктоптеригоид. [ 4 ] [ 5 ]
У большинства парарептилов были большие орбиты (глазные розетки), значительно дольше (спереди до спины), чем область черепа за глазами. [ 5 ] Югальная . кость, которая образует нижний и задний край орбиты, имеет очень тонкий суборбитальный процесс (передняя ветвь), обычно нет подторого временного процесса (нижняя задняя ветвь) и толстый дорсальный процесс (верхняя задняя ветвь) Сквамосовые . и четырехлетные кости, которые лежат за югалом, довольно большие и сзади, чтобы приспособить внутренние уши [ 4 ] [ 5 ] Парарептилы традиционно считали череп-типом « Анапсид », с югальским, сквомозальным и квадратоджугальным, твердо зашившимся без каких-либо пробелов или разрезов между ними. Этот принцип по -прежнему относится к некоторым подгруппам, таким как Pareiasaurs. Тем не менее, известно, что растущее число парарептильных таксонов имело инфратвральный фенестр , большую дыру или эмаргинацию, лежащую среди костей за глазами. В некоторых таксонах маржа таких отверстий может включать в себя дополнительные кости, такие как верхняя челюсть или посторбиталь . [ 10 ] [ 11 ] Когда они видят сверху, задний край черепа прямой или имеет широкую среднюю складку. [ 5 ] Изнутри к внешней стороне задний край черепа образуется тремя парами костей: постпариеталами , таблицами и супратеропочками . Парарептилы имеют особенно большие супратеровицы, которые часто распространяются дальше назад, чем таблицы. [ 12 ]
Помимо длинных стройных челюстей мезозавров, большинство челюстей парарептиля были короткими и толстыми. Шаг -сустав образуется суставной ( в нижней челюсти) и квадратной (в верхней челюсти). Во многих парарептилях челюстный сустав смещается вперед на черепе мимо задней части мозга . [ 4 ] [ 7 ] Мышцы челюсти прикрепляются к короноидному процессу , треугольной шпоре в задней половине челюсти. Как зубчатая зубная кость, так и задний отверстие Intermandibularis (отверстие на внутренней поверхности челюсти) достигают до короноидного процесса. [ 4 ] [ 5 ] Сургульная кость, которая образует верхнюю заднюю часть челюсти, узкая и тарелка. [ 13 ]
Посткраниальный скелет
[ редактировать ]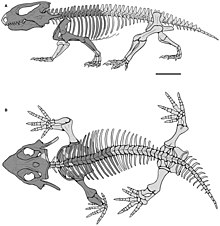
Были некоторые различия в форме тела парарептилов, причем ранние члены группы имели общий вид ящерицы , с тонкими конечностями и длинными хвостами. Наиболее успешные и разнообразные группы парарептилов, пареасавров и проколофонидов , имели массовые тела с уменьшенными хвостами и крепкими конечностями с короткими цифрами. Эта общая форма тела имеет отношение к другим «котилозаврам», такими как капториды , диадктоморфы и Seymouriamorphs . [ 3 ] Еще одна общая «котилозаврская» особенность в парарептилях - «опухший» внешний вид их позвонков , которые имеют широкие и выпуклые верхние поверхности. [ 12 ]
Парарептилям не хватало супраглоидного отверстия на лопате , отверстие, которое также отсутствует в варанопидах и неодиапсидах. [ 5 ] [ 11 ] У большинства был довольно короткий и толстый плечевой кости , который был расширен возле локтя. В отличие от ранних эвриптилей, внешняя часть нижней плечевой кости обладала как небольшой процесс супинатора, так и эктопикондилар -отверстия и канавку. [ 4 ] Умеющая среда, как правило, имеет плохо развитый процесс олекранона , еще одна черта, в отличие от самых ранних эвриптилей. [ 4 ] [ 5 ]
У большинства парарептилов был подвздошной подвздошной ткань , который был в форме вентилятора и вертикально (а не горизонтально), необычной чертой среди ранних амниотов. [ 4 ] [ 9 ] [ 7 ] Сакральные ребра, которые соединяют позвоночник с подвздошной сиденья, обычно были тонкими или вентиляционными, с большими зазорами между ними. [ 4 ] Задних конечностей, как правило, были не намного дольше, чем передние конечности, и имели толстые кости лодыжки и короткие ног. Есть некоторые исключения, такие как Eudibamus , ранний пермский болосаурид с очень удлиненными задними конечностями. [ 14 ]
История классификации
[ редактировать ]Название Parareptilia было придуман Олсоном в 1947 году для обозначения вымершей группы палеозойских рептилий, в отличие от остальных рептилий или eureptilia («истинные рептилии»). [ 15 ] Термин Олсена обычно игнорировался, и различные таксоны, позже известные как парарептилы, обычно не были помещены в исключительные группы друг с другом. Многие были классифицированы как «котилозавры» ( таксон для мусора с крепкими «примитивными» рептилиями или рептильными тетраподами) или « Anapsids » (рептилии без временных фенестров , таких как современные черепахи).
Использование Parareptilia было возрождено кладистическими исследованиями, чтобы ссылаться на те традиционные «анапиды», которые считались не связанными с черепахами. Gauthier et al. (1988) предоставили первые филогенетические определения для названий многих амниота таксонов и утверждали, что капторхиниды и черепахи были родственными группами, составляющими клад Anapsida (в гораздо более ограниченном контексте, чем обычно применялись). Название должно было быть найдено для клады различных пермских и триасовых рептилий, которые больше не включены в Anapsids. Термин Олсена «Парарептилы» был выбран для обозначения этой клады, хотя его нестабильность в их анализе означала, что Gauthier et al. (1988) не были достаточно уверены, чтобы возвести парарептилию в качестве формального таксона. Их кладограмма заключается в следующем: [ 16 ]
| Амниота |
| ||||||||||||



Laurin & Reisz (1995) обнаружил немного другую топологию, в которой рептилия разделена на парарептилию и eureptilia. Они утверждали, что тестодины (черепахи) были членами Parareptilia; Фактически, они явно определили Parareptilia как «Testudines и все амниоты, более тесно связанные с ними, чем с диапсидами». Captorhinidae был переведен в Eureptilia, в то время как Parareptilia включала черепах, а также многие таксоны, названные как таковые Gauthier et al. (1988). Было одно главное исключение: мезозавры были размещены за пределами обеих групп, в качестве сестринского таксона в рептилия Crown Group . Мезозавры все еще считались сауропсидами, так как они были ближе к рептилиям, чем к синапсидам. Традиционная группа «Anapsida» была отвергнута как парафилетическая сборка. Кладограмма Laurin & Reisz (1995) представлена ниже: [ 4 ]
| Амниота |
| ||||||||||||||||||||||||||||||

Напротив, в нескольких исследованиях в середине до конца 1990-х годов Оливье Риппель и Майкл Дебрага утверждали, что черепахи были на самом деле лепидозавраморфными диапсидами, связанными с сауроптеригианами . [ 17 ] [ 18 ] [ 9 ] [ 6 ] [ 7 ] Аффинность диапсидов черепах было подтверждено молекулярными филогениями . [ 19 ] [ 20 ] [ 21 ] [ 22 ] Первый филогенетический анализ всего генома был завершен Wang et al. (2013). Используя черновые геномы Chelonia Mydas и Pelodiscus sinensis, команда использовала самый большой набор данных черепахи на сегодняшний день в своем анализе и пришла к выводу, что черепахи, вероятно, являются родственной группой крокодилов и птиц (архозаврия). [ 8 ] Это размещение в диапсатах предполагает, что линия черепах потеряла характеристики черепа диапсида, поскольку черепахи обладают черепом AnapsId. Это сделало бы Parareptilia полностью вымершей группой с особенностями черепа, которые напоминают чередующие черепахи посредством сходящейся эволюции . С черепахами, расположенными за пределами парарептилей, Цудзи и Мюллер (2009) пересмотрели парарептилию как «самую инклюзивную кладу, содержащую Милретту Рубиджи и Проколофон Тригоницепс , но не Captorhinus aguti ». [ 3 ]
Кладограмма ниже следует анализу MS Lee, в 2013 году. [ 23 ]
| Амниота |
| ||||||



Кладограмма ниже следует анализу Li et al . (2018). [ 24 ]
|
† Parareptilis | |||||||||||||||||||||||||||||||||||||
| Eruptilia |
| ||||||||||||||||||||||||||||||||||||



Исследование, проведенное в 2020 году Дэвида П. Форда и Роджера Б.Дж. Бенсона, показало, что парарептилия была вложена в Диапсиде как сестринскую группу для неодиапсиды , а клада, содержащая неодиапсида и парарептилию, дублированные неорептилией, что предполагает, что парарептилов были древними диапсидом. Это исключило мезозавры , которые снова были признаны базальными среди сауропсидов. [ 11 ] Некоторые исследования показали, что парарептилиа является парафилетической, с некоторыми парарептилами, более тесно связанными с диапсидами, чем с другими парарептилами, с Simões et al. (2022) Использование Neoreptilia для клады, содержащей procoloponoorpha+neodiapsida. [ 25 ]
Эволюционная история
[ редактировать ]Самыми старыми известными парарептилами являются болозавр эрпетоникса и аклейсторинид углеродако из позднего каменноугольного ( московийского -гжелианского ) Северной Америки, который представляет собой единственные известные каменноугольные парарептилы, что указывает на то, что первоначальная диверсификация группы произошла в позднем каменном. [ 1 ] Многочисленные парарептильные линии появились во время раннего пермца, и группа достигла космополитического распределения. Разнообразие парарептилей снизилось к концу пермского и проколофоноидов , которые впервые появились во время позднего пермца, была единственной группой парарептилов, чтобы пережить пермс -триассическое событие вымирания . Разнообразие проколофонидов резко снизилось, начиная с среднего триаса, и к концу триаса группа вымерла. [ 26 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Манн А., Макдэниел Э.Дж., Маколвийл Э.Р., Источник HC (ноябрь 2019). « Carbondraco Lundi Gen et sp. Nov., самый старый Parareptle, из Линтона, Огайо, и новое понимание раннего излучения репта » Королевского общества Открытая наука 6 (11): 191191. Bibcode : 2019rsos .... 691191m Doi : 10.1098/ rsos.1 PMC 6894558 31827854PMID
- ^ Бота-Бринк Дж., Смит Р.М. (2012-09-01). «Палеобиология триасовых проколофонидов, выведенная из костной микроструктуры». Comptes rendus palevol . 11 (6): 419–433. doi : 10.1016/j.crpv.2012.03.002 . ISSN 1631-0683 .
- ^ Jump up to: а беременный в дюймовый Tsuji LA, Müller J (2009). «Сборка истории парарептилии: филогения, диверсификация и новое определение клады» . Запись ископаемого . 12 (1): 71–81. doi : 10.1002/mmng.200800011 . ISSN 1860-1014 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не Лаурин М., Рейс Р.Р. (1995-02-01). «Переоценка ранней филогения амниоте» (PDF) . Зоологический журнал Линневого общества . 113 (2): 165–223. doi : 10.1111/j.1096-3642.1995.tb00932.x . ISSN 0024-4082 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Müller J, Tsuji LA (сентябрь 2007 г.). «Слушание по борьбе с импедансом в палеозойских рептилиях: свидетельство продвинутого сенсорного восприятия на ранней стадии эволюции амниота» . Plos один . 2 (9): E889. Bibcode : 2007ploso ... 2..889m . doi : 10.1371/journal.pone.0000889 . PMC 1964539 . PMID 17849018 .
- ^ Jump up to: а беременный Rieppel O, Debraga M (декабрь 1996 г.). «Черепахи как рептилии диапсидов» (PDF) . Природа . 384 (6608): 453–455. Bibcode : 1996natur.384..453r . doi : 10.1038/384453A0 . ISSN 1476-4687 . S2CID 4264378 .
- ^ Jump up to: а беременный в дюймовый и фон Дебрага М., Риппель О (1997-07-01). «Филогения рептилий и взаимосвязь черепах» . Зоологический журнал Линневого общества . 120 (3): 281–354. doi : 10.1111/j.1096-3642.1997.tb01280.x . ISSN 0024-4082 .
- ^ Jump up to: а беременный Wang Z, Pascual-Anaya J, Zadissa A, Li W, Niimura Y, Huang Z, et al. (Июнь 2013 г.). «Проектные геномы черепахи с мягкой оболочкой и зеленой морской черепахи дают представление о развитии и эволюции плана тела, специфичной для черепах» . Природа генетика . 45 (6): 701–706. doi : 10.1038/ng.2615 . PMC 4000948 . PMID 23624526 .
- ^ Jump up to: а беременный в Дебрага М., Рейс Р.Р. (1996-09-19). «Ранняя пермская рептилий Acleistorhinus pteroticus и его филогенетическое положение» (PDF) . Журнал палеонтологии позвоночных . 16 (3): 384–395. doi : 10.1080/02724634.1996.10011328 . ISSN 0272-4634 .
- ^ MacDougall MJ, Reisz RR (2014). «Первая запись общенавременного парарепта из раннего пермияна Северной Америки, с обсуждением парарептлианского висового височного фенестрации» Зологический журнал линейного общества 172 (3): 616–6 Doi : 10.1111/ Zoj.1
- ^ Jump up to: а беременный в Ford DP, Benson RB (январь 2020 г.). «Филогения ранних амниотов и сродство парарептилий и варанопиды» . Природа экология и эволюция . 4 (1): 57–65. doi : 10.1038/s41559-019-1047-3 . PMID 31900445 . S2CID 209673326 .
- ^ Jump up to: а беременный Tsuji LA, Müller J, Reisz RR (2012-01-01). «Анатомия эмеролета Левиса и филогения парарептилов Nycteroleter» (PDF) . Журнал палеонтологии позвоночных . 32 (1): 45–67. doi : 10.1080/02724634.2012.626004 . ISSN 0272-4634 . S2CID 55268829 .
- ^ Haridy Y, Macdougall MJ, Reisz RR (13 декабря 2017 г.). «Нижняя челюсть раннего пермского парарептиля Delorhynchus, сначала свидетельство множественных зубчатых короноидов в рептилии». Зоологический журнал Линневого общества . 184 (3): 791–803. doi : 10.1093/Zoolinnean/zlx085 . ISSN 0024-4082 .
- ^ Берман Д.С., Рейс Р.Р., Скотт Д., Хенрири А.С., Сумида С.С., Мартенс Т (ноябрь 2000 г.). «Ранняя пермская двуночная рептилия». Наука . 290 (5493): 969–972. Bibcode : 2000sci ... 290..969b . doi : 10.1126/science.290.5493.969 . PMID 11062126 .
- ^ Олсон Е.С. (1947). Рой С.К. (ред.). «Семейство Diadectidae и ее приспособление к классификации рептилий» . Геология . ФИЛИАНА. 11 (1). Чикаго: Чикагский музей естественной истории: 1–53 . doi : 10.5962/bhl.title.3579 . Получено 15 января 2015 года .
- ^ Gauthier JA, Kluge AG, Rowe T (1988). «Ранняя эволюция амниоты». В Бентоне М.Дж. (ред.). Филогения и классификация тетрапод, том 1: амфибии, рептилии, птицы. Систематическая ассоциация Специальный том . Тол. 35а. Кларендон Пресс. С. 103–155.
- ^ Риппель О (1994). «Остеология Симозавра Гайярдоти и отношения стволовой группы Sauropterygia» . ФИЛИАНА ГЕОЛОГИЯ . 28 (1462): I-85.
- ^ Риппель О (1995). «Исследования по формированию скелетов у рептилий: последствия для отношений черепах». Зоология . 98 : 298–308.
- ^ Zardoya R, Meyer A (ноябрь 1998 г.). «Полный митохондриальный геном предполагает сродство диапсидов черепах» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (24): 14226–14231. Bibcode : 1998pnas ... 9514226Z . doi : 10.1073/pnas.95.24.14226 . PMC 24355 . PMID 9826682 .
- ^ Ивабэ Н., Хара Ю., Кумазава Ю., Шибамото К., Сайто Ю., Мията Т., Като К (апрель 2005 г.). «Сестринская групповая связь черепах с птичьей кладом, выявленной ядерными ДНК-кодированными белками» . Молекулярная биология и эволюция . 22 (4): 810–813. doi : 10.1093/molbev/msi075 . PMID 15625185 .
- ^ Roos J, Aggarwal RK, Janke A (ноябрь 2007 г.). «Расширенные митогеномные филогенетические анализы дают новое понимание эволюции крокодилиана и их выживаемости меховой третичной границы». Молекулярная филогенетика и эволюция . 45 (2): 663–673. doi : 10.1016/j.ympev.2007.06.018 . PMID 17719245 .
- ^ Катсу Ю., Браун Э.Л., Гилетт Л.Дж., Игути Т. (2009). «От рептильной филогеномики до геномов рептилий: анализ протоонкогенов C-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. doi : 10.1159/000297715 . PMID 20234127 . S2CID 12116018 .
- ^ Ли М.С. (декабрь 2013 г.). «Происхождение черепах: понимание филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. doi : 10.1111/jeb.12268 . PMID 24256520 . S2CID 2106400 .
- ^ Ли, Чун; Фрейзер, Николас С.; Риппель, Оливье; Ву, Ся-Чун (август 2018 г.). «Триасовая черепаха с беззурным клювом» . Природа . 560 (7719): 476–479. doi : 10.1038/s41586-018-0419-1 . ISSN 0028-0836 . PMID 30135526 . S2CID 52067286 .
- ^ Simões, tr; Kammerer, CF; Caldwell, MW; Пирс, SE (2022). «Последовательные климатические кризисы в глубоком прошлом привело к ранней эволюции и радиации рептилий» . Наука достижения . 8 (33): EABQ1898. doi : 10.1126/sciadv.abq1898 . PMC 9390993 . PMID 35984885 .
- ^ Macdougall MJ, Brocklehurst N, Fröbisch J (март 2019 г.). «Видовое богатство и неравенство парарептилов в терминальном массовом вымирании» . Разбирательство. Биологические науки . 286 (1899): 20182572. DOI : 10.1098/rspb.2018.2572 . PMC 6452079 . PMID 30890099 .






